橡膠樹雜交子代遺傳特性及遺傳多樣性分析
2022-06-06 13:25:04李文秀賀軍軍張華林翁俊亮李進良
西南農業學報 2022年5期
關鍵詞:產量
李文秀,賀軍軍,張華林,翁俊亮,李進良,羅 萍
(1. 中國熱帶農業科學院湛江實驗站/廣東省旱作節水農業工程技術中心,廣東 湛江 524091; 2. 中國熱帶農業科學院南亞熱帶作物研究所,廣東 湛江 524091)
【研究意義】巴西橡膠樹(Heveabrasiliensis)為大戟科(Euphobiaceae)橡膠樹屬(Hevea)多年生喬木,染色體2n=2x=36,原產于亞馬遜森林[1],在我國主要分布于海南、云南和廣東等省份。天然橡膠是主要的工業原料之一,廣泛應用于醫療、交通和國防等方面,是無污染可再生的重要自然資源[2]。根據國際橡膠研究組織提出的意見,隨著經濟發展,全球對橡膠的需求將會持續增加[3]。橡膠樹雜交育種是最常規的育種方法,也是選育優良橡膠樹品種的一個基本策略[4]。在橡膠育種實踐中,優良品種的選擇一般須通過有性雜交。橡膠樹雜交育種是指選擇各種特異親本進行雜交,在子代中選擇優良單株,利用芽接擴繁優良無性系進行推廣,是橡膠樹優良品系創育的一個基本策略[5]。實踐證明,通過雜交育種,育種家們選育出了一批優良品種,如熱研7-33-97[6]、熱墾525[7]、PR107[8]、RRIM600[9]、GT1和PB86[10]等品種。提前發現雜交子代的優劣能反映出親本雜交的有效性,縮短育種進程,及時更正育種策略。目前,經有性雜交選育出的抗寒性強產量中等湛試327-13、湛試327-4等[11]優良品系,都來自同一對親本,這反映出了親本選配合理,能將部分優良性狀傳遞給后代,縮短了品種選育的成本與時間。將傳統表型方法與標記選擇相結合,可以更準確地實現雜交后代的鑒定。形態鑒定雖然直接方便,但數量有限,易受環境影響。研究表明,具有相似形態的相似個體可能包含較大的遺傳變異,這些變異可以通過分子標記來識別[12]。SSR分子標記具有多態性高、重復性好、操作簡單等優點[13],已被廣泛應用于橡膠樹種質資源的研究,如構建橡膠樹遺傳圖譜[14]、種質多態性分析[13]、品種鑒定[15]、遺傳多樣性分析[16]。【前人研究進展】在其他作物上,表型特征和分子標記用于雜交后代雜種優勢評價和遺傳多樣性分析已較為普遍。如Monica等[17]利用SRAP標記研究了花生殖細胞及其F1代的遺傳變異及親緣關系。Gao等[18]利用12個表型和11個SRAP標記對45個牡丹品種的遺傳多樣性進行了分析。此外,橡膠樹表型多樣性分析也有相關的研究報道[19]。【本研究切入點】由于橡膠親本基因雜合度高,F1雜種往往存在較大差異。且到目前為止,橡膠樹F1雜種的遺傳特征和優劣特性研究依然比較有限。【擬解決的關鍵問題】本研究分別以PR107和93-114為父本和母本,通過剖析親本以及雜交后代群體的重要性狀和遺傳關系,從而鑒定出雜交子代的變異程度及優劣情況,旨在為橡膠樹雜交育種和親本選配提供參考意義。
1 材料與方法
1.1 材料來源
供樣材料來自中國熱帶農業科學院湛江實驗站收集保存的93-114(母本)和PR107(父本)35份雜交后代群體資源,葉片形態如圖1所示,于2016年建立親本和后代群體的初級系比試驗區,試驗區分為3個小區,每小區按照株行距為3 m×3 m隨機種植親本和后代群體,每份資源相鄰種植3株,共3次重復。

圖1 雜交子代葉片形態
SSR標記:104對SSR引物,24對來自Souza[20],4對來自Saha[21],5對來自謝黎黎[22],71對來自Gouva[23],由上海生工生物技術有限公司合成,每對引物都進行重復擴增。
1.2 試驗方法
1.2.1 表型性狀的調查 型性狀主要調查橡膠樹莖干、產量和葉片等相關性狀,包括26個描述性狀和6個測量性狀。按照黃華孫編寫的《橡膠樹種質資源描述規范和數據標準》[24]對26個描述性狀進行調查,具體調查性狀和分級標準見表1。大葉柄長度,小葉柄長度,小葉枕膨大長度,主測脈角度4個性狀利用直尺和角度尺進行測量,每份資源測量3株,每株取5個重復。樹圍從2017年開始進行測量,每年12月用卷尺測量距離地面1 m處周長,用厘米進行表示。割膠參考Souza[25]的方法,在2020年9月和12月,使用S/2 d/3方法,在離地面50 cm的地方進行開割,每次割5刀,收集乳膠進行烘干稱重,以克進行計量,每株產量計算方法以1刀進行計算,取3個小區的平均值得出。

表1 橡膠樹F1性狀描述分級

續表1 Continued table 1
1.2.2 基因組DNA提取 參考Murray等[26]的方法對橡膠樹基因組葉片DNA進行提取。用1%瓊脂糖凝膠電泳檢測純度與濃度,原液保存于-20 ℃備用。
1.2.3 引物篩選 用4份(PR107,93-114,A4和A8)表型差異較大的材料對合成的104對引物進行篩選,選出擴增效果好、條帶清晰穩定的多態性引物15對,引物序列如表2所示。

表2 橡膠樹SSR引物名稱及序列
1.2.4 SSR-PCR體系 PCR擴增反應在TAKARA TP-600 PCR儀上進行。采用10 μL反應體系,反應體系包含DNA模板1 μL,ddH2O 4 μL,4 μL mix,上游引物和下游引物各0.5 μL。反應條件參考Liu等[27]的做法:94 ℃預變性5 min;94 ℃變性30 s,復性30 s(溫度以引物最適退火溫度而定),72 ℃延伸30 s,35個循環;72 ℃延伸7 min。
1.2.5 PAGE凝膠電泳及檢測 采用6%聚丙烯酰胺凝膠對PCR產物進行電泳,電壓180 V,電流80 A,時間60~90 min。銀染方法參考Creste等[28]的方法。
1.2.6 數據分析 在Excel 2016中計算各性狀的變異系數和多樣性系數[29];在Excel中計算每份材料年平均生長量,并使用ORIGIN v7.0繪制圖表。采用SPSS 21.0軟件計算最大值、最小值、平均值、標準差。計算中親值和雜種優勢如下公式所示:
MPs= (P1+P2)/2
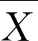
對擴增穩定清晰的條帶采用“0-1”記錄其位置,在相同的遷移位置上的條帶標記為“1”,無帶標記為“0”,建立0、1的SSR標記矩陣圖。
先用DataFormater軟件[30]進行數據轉換,利用 NTSYS-pc V2.10 軟件按UPGMA方法進行聚類分析;利用PopGen3.2計算群體的觀測等位基因數(Na)、有效等位基因數(Ne)、觀察雜合度(Ho)、Shannon’s信息指數(I);使用Powermaker Version3.25軟件計算PIC值和基因型數;利用MEGA(V7.0)和ITOL在線工具graphic (https://itol.embl.de/)進行遺傳距離分析和繪制樹狀圖。
2 結果與分析
2.1 表型性狀的多樣性分析
橡膠樹93-114×PR107組合雜交子代F1群體主要表型性狀的變異系數和遺傳多樣性指數見表3,32個表型性狀的平均變異系數為32.59%,變異系數最大的表型性狀是葉枕溝(60.90%),最小的是小葉柄溝(0)。變異系數超過50%的有葉枕溝和大葉柄形狀,說明這2個性狀變異較豐富。而小葉柄溝(0)、小葉枕膨大(22.88%)、葉片質地(19.40%)、葉片顏色(18.14%)、三小葉間距(13.30%)、大葉柄長度(14.32%)和主側脈角度(3.66%)變異系數均低于25%,表明以上性狀遺傳特性較穩定;其中小葉柄溝和主側脈角度變異系數接近于0,說明這2個性狀在雜交子代中基本沒有差異。
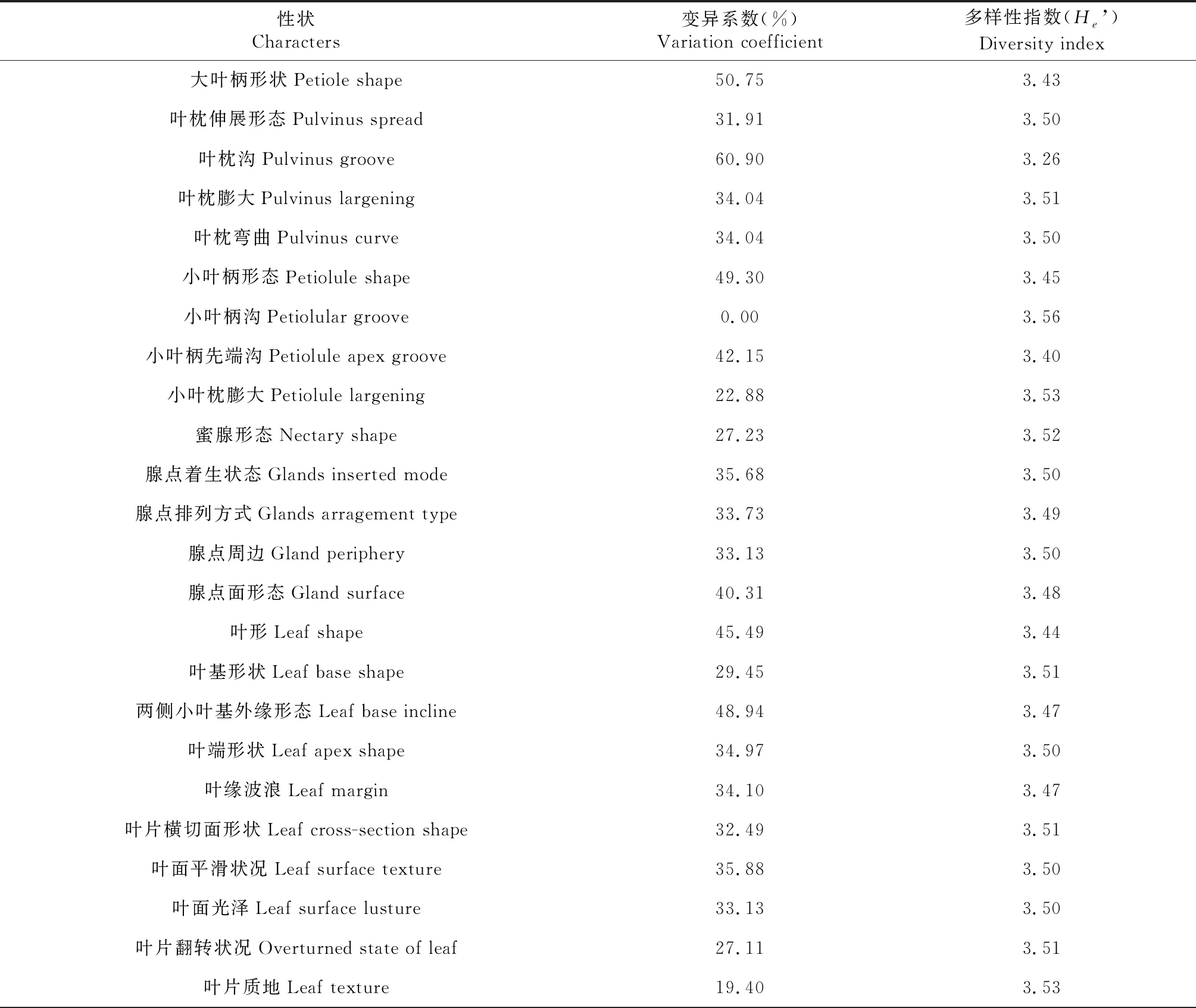
表3 雜交后代F1群體的變異系數和遺傳多樣性指數
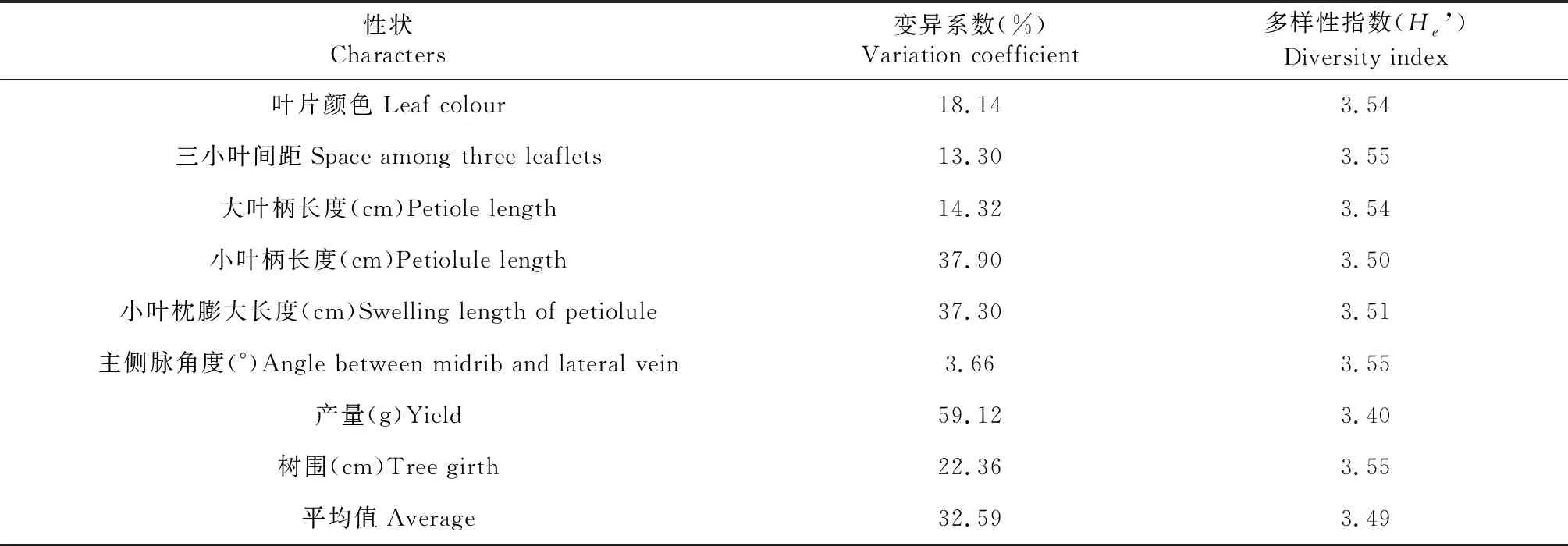
續表3 Continued table 3
32個表型性狀多樣性指數分布在3.22~3.53,平均值為3.46,表明雜交子代表型性狀分布較為均勻。多樣性指數受種數和均勻度的影響,在種數相同的情況下,Shannon指數越大,均勻性越好。其中,小葉柄溝(3.56)多樣性指數最大,其分布較為均勻;而葉枕溝(3.26)的多樣性指數最小,分布最不均勻。總體上,32個性狀的多樣性指數較大,且差異較小,表明表型性狀在供試材料中分布較為均勻。
2.2 產量和樹圍雜種優勢分析
雜交后代和親本的樹圍和產量統計結果(表4)顯示,雜交后代的平均樹圍較親本有所增加,而平均產量較親本有所減少。雜交后代的平均樹圍和年樹圍增長量均大于中親值,雜種優勢分別為101.2%和117.2%,雜種優勢較強,這與小麥的雜種育種較為相似,雜交種雜種優勢通常會高于中親值[31]。而平均產量較中親值卻稍有下降,雜種優勢為95.38%,處于中親本優勢。但值得注意的是,有40%雜交后代的產量高于雙親,說明在F1雜種中仍然可以選擇高產單株。

表4 雜種優勢
2.3 雜交后代樹圍和產量比較分析
在35個雜交后代中,產量最高的是材料A38,最低的是A9。從圖2可以看出,12月份的產量數據中,材料A38、A22、A12和A25單株產量均在20 g以上,其中A38產量最高,單株產量超過25 g,產量最低的是A9。9月份的產量數據中,A29、A25、A24、A26、A38單株產量均超過5 g,其中A29單株產量最高,產量最低的也是A9。綜合來看,A38的平均產量最高,A9的平均產量最低。
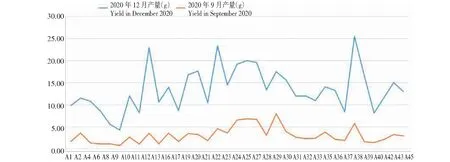
圖2 雜交子代早期試割產量
從圖3中可以看出,A22每年都是樹圍最大的材料,最大樹圍材料與最小樹圍材料的差異在逐年增大。2017—2020年4年期間,材料A22的樹圍最大(39.3 cm),而A8、A23、A21和A23分別是每年樹圍最小的材料,極差分別為4.6、8.9、9.6和10.8 cm,可以看出,極差每年都在增大,最小樹圍材料每年都在輪換,說明個別材料前期生長慢,但后期生長會逐漸加快。此外,2017—2020年,雜交后代樹圍年生長量為5.6~10.4 cm,平均年生長量為6.7~8.8 cm(表5),年均周長增量逐年降低,說明苗期生長速度快于后期。
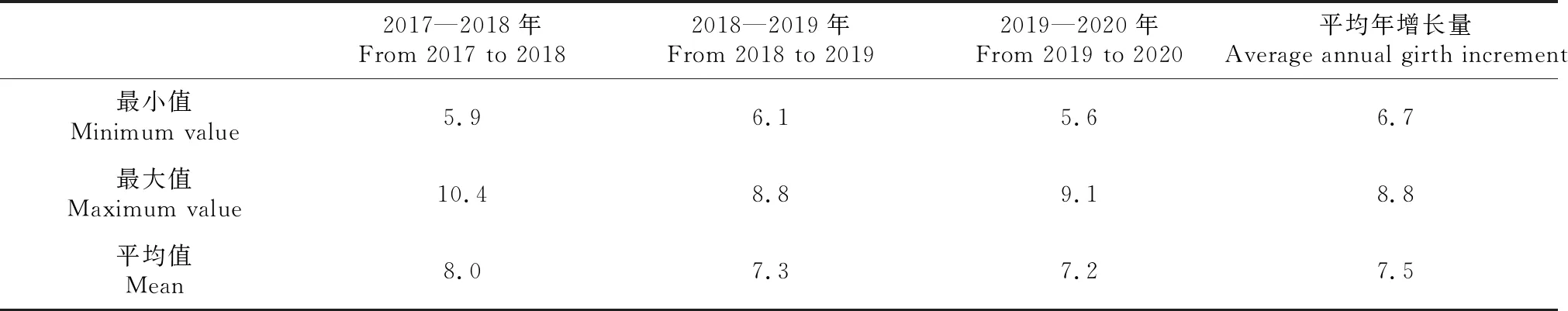
表5 雜交子代樹圍的年生長量
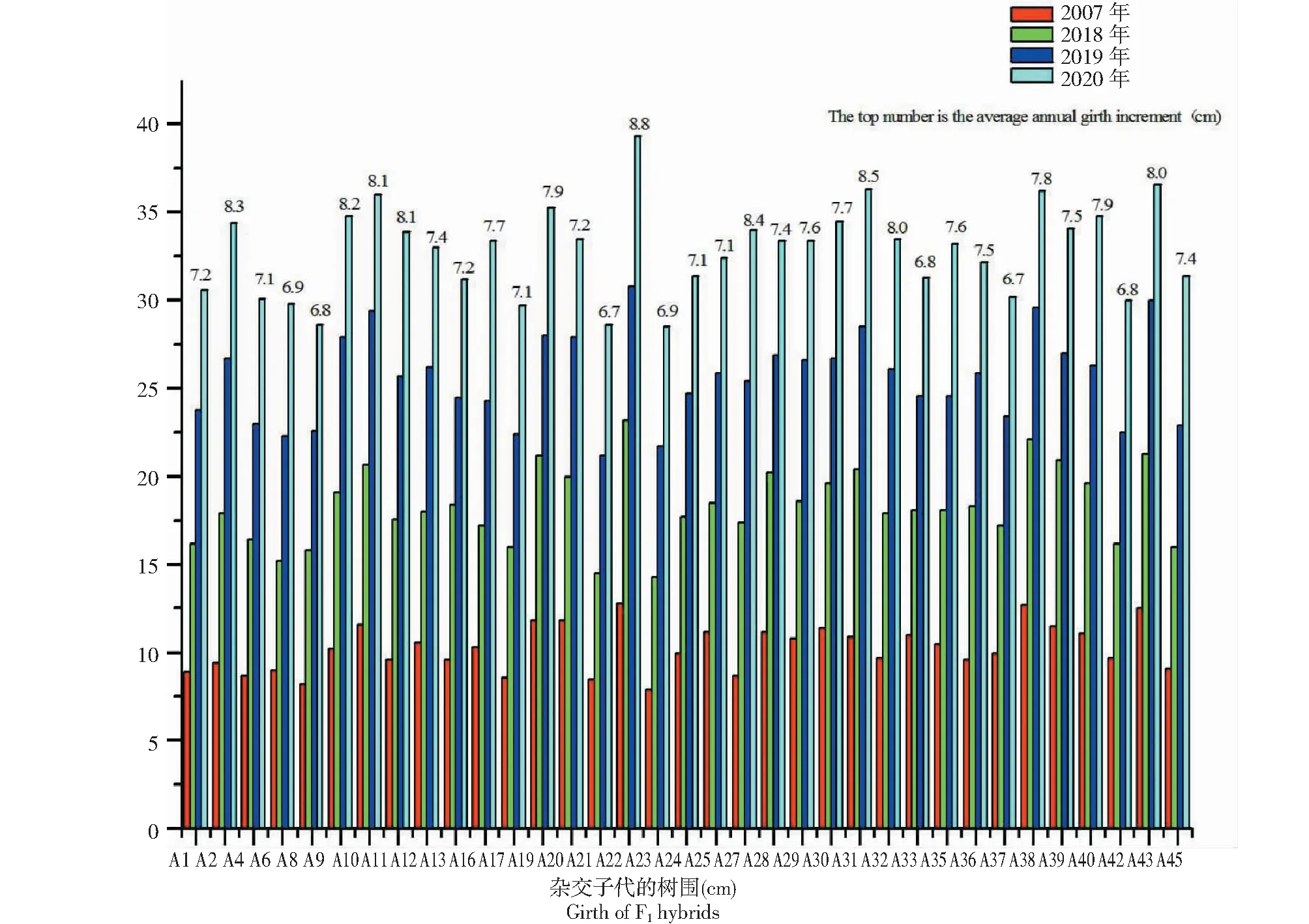
圖3 2017—2020年雜交子代樹圍生長
2.4 SSR標記多態性分析
利用表型差異較大的4個材料對104對引物進行擴增篩選了15對多態性引物,利用15對引物對親本和雜交后代進行多態性檢查,結果(表6)表明,每對引物位點的等位基因數為2~4個,平均為3個,有效等位基因數在1.4706~3.6953,所用引物位點Shannon’s指數、觀測雜合度(Ho)、預期雜合度(He)和多態信息含量(PIC)分別在0.5004~1.3459、0.1714~1.0000、0.3246~0.7400和0.2710~0.6827,其中ACT59(0.2710)位點的多態性最低,ACT61(0.6827)位點的多態性最豐富。
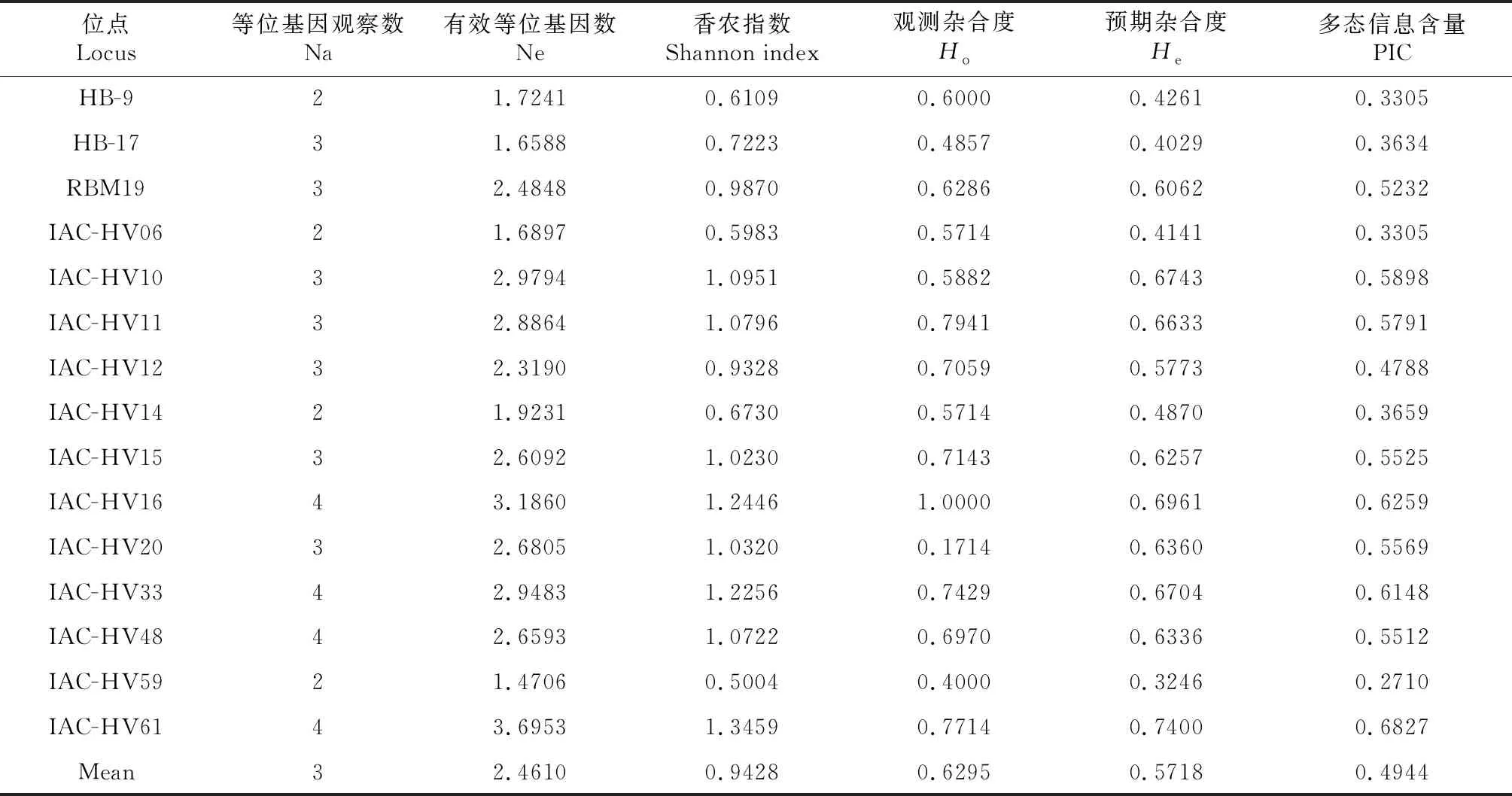
表6 SSR標記多態性分析
2.5 SSR聚類分析
聚類分析結果(圖4)表明,35個雜交后代可分為3類,遺傳相似系數在0.54~0.89(結果未列出),雜交產生的遺傳變異較多,個體間遺傳變異較明顯。其中12個雜交后代 (A35、A1、A17、A16、A27、A20、A6、A31、A19、A2、A42和A40)和父本66 (PR107)分為第一組(I);另外20個雜交后代(A4、A13、A9、A43、A36、A30、A29、A24、A38、A23、A28、A39、A33、A10、A25、A45、A21、A12、A37、A22)組成第二組(II);母本45(93-114)和材料A8、A11和A32形成第三組(III)。此外,由于標記數量有限,在進化樹中分辨率不夠高,無法區分某些雜交后代,如材料A12和A22。
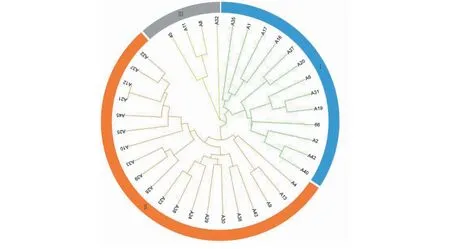
圖4 雜交子代SSR聚類分析
3 討 論
對于基因型高度雜合作物,獲得新類型材料和選育有價值新品種的一個有效辦法就是開展雜交育種[32]。開展雜交育種的前提是選擇性狀優良互補、遺傳差異較大和一般配合力強的親本,然后通過研究雜交后代的遺傳和變異規律,了解性狀組合情況及遺傳傾向[33],最終選出理想的品系或品種。
本研究對35份PR107和93-114組合F1雜交后代的32個主要表型性狀的變異類型和形態多樣性進行分析,雜交后代平均變異系數和遺傳多樣性指數分別為32.59%和3.46,具有豐富的形態多樣性。這說明了雜交親本基因型高度雜合,對于選出具有雜種優勢的材料具有重要的意義。葉枕溝(60.90%)和大葉柄形狀(50.75%)2個性狀變異較大,結果與張曉飛等[34]對119份橡膠樹栽培種的21個農藝性狀進行比較分析的結果較為類似;與之不同的是遺傳多樣性指數平均值,張曉飛得出的結果是0.83,而本研究是3.46,其中的差異有待后續研究。另外,在本研究中,變異系數很低的情況下遺傳多樣性系數一樣也很高。變異系數和多樣性指數都是反映材料多樣性的重要指標,變異系數反映的是變異類型的離散程度,遺傳多樣性系數反映的是性狀分組數目和組內性狀分布的均勻程度[35],兩者不存在相關關系。
利用該組合和雜交后代群體有可能篩選出高產子代和速生子代。樹圍和年增量的雜種優勢指數均大于100%,但產量雜種優勢小于100%,表明在樹圍和產量上均存在雜種優勢,但產量在該雜交后代群體中稍弱。比較2020年9月和12月的產量結果,可以發現產量曲線趨勢基本一致,說明產量結果可靠,可重復性好。唯一的偏差是材料A12,其結果的準確性有待日后再次驗證。
產量和樹圍綜合分析結果表明,材料A38和A22在雜交后代綜合表現較好。通過比較樹圍和產量數據,雜交后代中,A22、A43和A38的樹圍較好,樹圍生長顯著快于親本;A38、A22和A25是雜交后代中產量最高的3個材料。因此,綜合來看,材料A38和A22是較好的種質。
通過對雜交后代的遺傳多樣性分析,了解親本選配和雜交后代遺傳背景情況,對育種工作具有重要的指導意義。PR107是初生代無性系[5],具有的優良特征描述,以PR107為親本,先后選育出的品種有熱研7-33-97、云研77、大豐95等[36];品種93-114是華南熱帶作物科學研究院粵西試驗站于1967年從無性系天任31-45和合口3-11雜交子代中選育的次生代無性系,抗寒力強,抗風能力好[37]。本研究應用分子標記對35份PR107和93-114雜交后代進行了遺傳多樣性分析,雜交后代中大多數位點與父本PR107的基因型位點相似,說明PR107和93-114組合雜交F1代更偏向于父本遺傳。遺傳多樣性分析結果可將雜交后代分為3類,材料類型較為豐富,反映出雜交親本的選配合理。本研究從表型性狀和分子標記兩個方面分析了親本和雜交子代群體的遺傳特性和遺傳分化,為親本組合的甄選和雜交后代的差異分析等工作提供參考依據。
4 結 論
93-114×PR107雜種F1表型性狀的變異系數和多樣性指數分別為32.59%和3.49%。雜種優勢分析表明,雜種一代的樹圍雜交優勢比產量強。綜合產量和周長分析結果表明,A38和A22在雜交種中表現較好。利用UPGMA對雜交種進行聚類分析,結果表明,雜交種可分為3個類群。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
