植物激素調控頂端分生組織發育的研究進展
2022-06-06 13:55:06胡慶毅陳本佳楊立凡馬永海李沁妍李慶懋黃立鈺
湖南農業科學 2022年1期
胡慶毅,陳本佳,楊立凡,馬永海,李沁妍,李慶懋,黃立鈺
(云南大學農學院資源植物研究院,云南 昆明 650000)
高等植物的器官形成離不開分生組織(meristem)。分生組織是在植物體的一定部位,具有持續或周期性分裂能力的細胞群,又被稱為植物干細胞。由其分裂產生的細胞排列緊密,無細胞間隙;其中一小部分仍保持高度分裂的能力,大部分則陸續長大并分化為具有一定形態特征和生理功能的細胞,構成植物體的其他各種組織,使器官得以生長或新生[1]。分生組織在植物體內根據分布位置主要分為頂端分生組織和側生分生組織。其中,頂端分生組織發育為根、莖,使植株得以伸長生長,側生分生組織則形成腋芽(axillary bud meristem,AM)和側根(lateral root,LR),進而產生新的分枝并重新建立頂端分生組織和側生分生組織。在高等植物的發育中,胚胎從縱軸兩端到胚胎兩端分別形成特殊區域,即莖端分生組織(stem apical meristem,SAM)和根端分生組織(root apical meristem,RAM)[1]。分生組織的存在保證了植物整個生長發育過程的可塑性,通過干細胞的分裂和分化產生新器官所需要的子細胞[2]。
1 植物頂端分生組織的結構特征及核心調控基因
分生組織的發育主要依賴于植物干細胞的分化,干細胞分化后的一部分子細胞經分裂分化形成新器官,一部分始終保持自身未分化狀態,并在分生組織中心區域內維持干細胞的自我更新[3-4]。
SAM 形成于胚胎發育的早期階段,結構上分為中央分生組織區(central zone,CZ)、肋狀分生組織區(rib zone,RZ)和外緣分生組織區(peripheral zone,PZ)。CZ 區由全能干細胞組成,處于未分化狀態,細胞分裂周期長,分裂頻率低,由外向內分為L1、L2、L3這3 個不同的胚性細胞層,分別形成表皮、皮下組織以及芽的相關內部器官;RZ 區為維管分生組織提供細胞支持;PZ 區是進一步細胞分裂、分化發育成為側生器官的核心區域[5]。CZ 區需要提供干細胞補充到PZ 區和RZ 區,CZ 區和RZ 區之間還存在一個組織中心(Organization center,OC)區域,它介導不同區域間信息的傳遞和干細胞的維持(圖1A)。SAM 中央區的組織中心細胞和其上方的干細胞構成的干細胞微環境是維持分生組織的結構基礎。wuschel(WUS)基因受生長素的誘導,是體細胞胚胎發生過程中干細胞更新的關鍵,并維持未分化干細胞自我更新的平衡[6]。WUS 蛋白作為建立SAM 的一個重要調控因子,在WUS 表達區域內維持SAM 組織中心干細胞的數量,同時和clavata3(CLV3)形成負反饋調節機制以維持SAM 的大小[7];STM(SHOOT MERI STEMLESS)在維持莖端分生組織中干細胞的產生也是必需的,它在整個莖端分生組織中的表達阻止了干細胞向器官特異性細胞的轉化[8-9]。
RAM 鄰近根冠,由根端靜止中心(quiescent center,QC)與根端干細胞區域組成。QC 位于RAM 的根端并在該區域衍生出根端干細胞,根端干細胞產生子細胞,子細胞隨后分化成為過渡區細胞的成員(圖1B)。因此,干細胞分裂和分化速度之間的平衡對維持根端分生組織至關重要。在靜止中心的形成和根部干細胞的建立過程中,主要受依賴生長素信號的plethora(PLT)途徑和不依賴生長素信號的shortroot(SHR)/scarecrow(SCR)途徑的調控[10-12]。其中PLT基因編碼了含有AP2(apetala2)結構域的轉錄因子,可以促進細胞分裂;SHR激活SCR的轉錄,SCR通過細胞自主性機制維持QC 的穩定[13]。QC 作為根發育中心的同時也是RAM 確立和維持的必要條件,在胚根或側根形成的早期原基細胞中開始分裂,最終形成根分生組織的早期結構。側根分生組織是從根頂端分生組織外更遠的內皮層開始的,在分生組織的OC 內干細胞分裂分化形成側根原基(lateral root primordia,LRP),繼而發育成為LR。研究發現wuschel homeobox5(WOX5,在QC 細胞中表達)和PLTs(在QC 周圍的干細胞中表達)是干細胞形成和維持的主要調節因子,在生長素信號下游起作用,以維持根端干細胞的活性[14]。
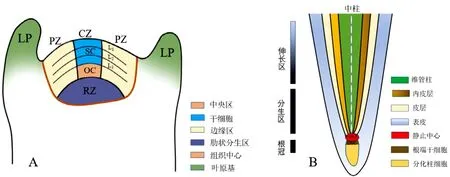
圖1 植物莖端(A)和根端(B)分生組織
2 生長素、細胞分裂素調控分生組織的作用及相關信號途徑
生長素(auxin)和細胞分裂素(cytokinin,CTK)是植物體內2 類重要有機化合物,調節著細胞分化、生長方向、發育過程以及植物生命活動等過程。植物形態結構的形成是植物適應環境并長期進化的結果,植物在生長發育過程中,感受外界環境信號,傳遞并影響生長素和細胞分裂素等激素的合成、運輸、信號傳遞和代謝,進而影響植物分生組織的維持和分化。生長素在SAM 中抑制細胞分裂而促進細胞分化,而在RAM 中,生長素引導分生組織細胞分裂;細胞分裂素在SAM 和RAM 中均可以通過抑制生長素響應信號阻礙細胞分裂,并促進細胞從分裂狀態向分化狀態轉變[15-16]。
2.1 生長素對植物分生組織的調控
2.1.1 生長素的合成及運輸 生長素(auxin)在植物體內主要以吲哚-3-乙酸(indole-3-acetic acid,IAA)形式存在[17]。IAA 的生物合成主要有色氨酸依賴型和
非色氨酸依賴型2 種途徑。其中,色氨酸依賴型途徑是植物體內IAA 的主要合成途徑,TAA1(tryptophan aminotransferase of arabidopsis 1,TAA 家族)催化色氨酸生成吲哚丙酮酸(indole-3-pyruvate,IPA),進而由YUCCA(YUC 家族)催化IPA 生成IAA,并參與器官的形成[18]。生長素除了在植物體內原位合成發揮作用外,還能運輸到其他部位,使植物組織器官的生長素濃度產生差異,進而發揮作用[19]。研究表明,生長素運輸包括長距離運輸和短距離運輸,主要有吲哚-3-乙酰胺(IAM)途徑、吲哚-3-乙醛肟(IAQx)途徑、色胺(TAM)途徑和吲哚-3-丙酮酸(IPA)途徑4 種運輸途徑。長距離運輸是指生長素從合成部位(如莖端分生組織)運輸到其他組織的運輸,例如頂端優勢的作用機制:生長素在莖頂端分生組織合成,向下運輸,進而抑制側生分生組織的生長發育;短距離運輸為細胞之間的單向極性運輸,主要通過生長素轉運蛋白pinformeds(PINs)進行生長素細胞間運輸,形成生長素的梯度分布從而調控分生組織發育[20-21]。
2.1.2 生長素信號調控途徑 生長素合成并運輸至分生組織區域達到最適水平,通過Aux/IAA-ARF 信號途徑激活下游生長素響應基因,從而調節分生組織的發育[22]。在分生組織中,生長素可以通過抑制分生組織細胞分化基因shoot meristimless(STM)的表達,進而抑制擬南芥A 型調節因子ARR7/ARR15和葉極性基因PHAN的同源基因AS1、AS2(asymmetric leaves1、asymmetric leaves2)在分生組織CZ 區和PZ 區的表達,最終抑制器官原基的產生(圖2)。

圖2 植物激素和相關基因的調控關系
高濃度的生長素會抑制植株側生分生組織的發育,而適當的生長素濃度則可促進側生分生組織的發育,并通過向外運輸PINs 加快側原基的形成。生長素還可以調控根的形成、增殖、伸長和分化,通過PLT、ARF、SCF、LBD等基因來維持RAM 的穩定性,并調控側根分生組織的發生。LBD家族轉錄因子JLO(jagged lateral organs)一方面可以抑制PIN1的表達使生長素在內皮層積累,促進中柱鞘細胞的增生使其發育成側根原基[16,23],另一方面可以通過調控生長素受體TIR1、AFB1和同源異型盒基因KNOX1的表達,控制莖端分生組織的生長發育[23]。生長素響應因子ARF10/ARF16負調控wuschel-related homeobox5(WOX5),并將其表達區域限制在QC 區域,從而抑制PLT基因的表達以影響干細胞的分化,進而影響根端分生組織的形成及發育。生長素還可通過負調控SCFTIR1復合物的形成抑制ARF7/ARF19的表達,進而促進細胞在根端分生組織的不對稱分裂以形成側根原基[16,24]。
2.2 細胞分裂素對植物分生組織的調控
2.2.1 細胞分裂素的合成及運輸 細胞分裂素(cytokinin,CTK)在種子萌發、細胞分化和器官發育等植物發育和生理過程中都發揮了重要作用[25]。細胞分裂素關鍵合成酶主要有異戊烯基轉移酶IPT、細胞色素單氧酶P450(CYP735A1、CYP735A2)和細胞分裂素激活酶LOG[26]。在分生組織區域,細胞分裂素的生物合成還受STM的影響,STM通過誘導IPT7的表達來促進細胞分裂素的生物合成[27]。在SAM 中細胞分裂素通過KNOX1(knotted1-like homeobox1)蛋白介導運輸,并且負調控生長素的轉運從而影響葉原基的形成,同時還通過KNOX1 維持著分生組織的活性,調控植物器官的形態建成[27]。
2.2.2 細胞分裂素信號調控途徑 細胞分裂素主要通過依賴CLV3 和依賴AHKs 的信號調控途徑,進而影響STM、IPT7的表達量,實現對植物分生組織的調控。細胞分裂素以AHKs 介導的途徑誘導WUS和CLV3,進而促進SAM 的形成[28]。同時,細胞分裂素激活酶LOG 可將細胞分裂素前體的一種非活性反式玉米素-糖甘轉化為活性細胞分裂素,在STM 啟動子的控制下,促進細胞分裂素合成關鍵基因IPT7的表達,從而促使側芽的形成(圖2)。
擬南芥中有14 種細胞分裂素信號響應蛋白,其中有8 種A 型調節蛋白ARR 是細胞分裂素轉導的負調節因子,有6 種B 型調節蛋白ARR(ARR1/2/10/12/14/18)是細胞分裂素響應的正向調節因子[29]。細胞分裂素B 型應答因子可直接激活莖尖干細胞,并穩定WUS基因的表達水平,從而維持干細胞的活性[30]。細胞分裂素通過AHK2/AHK4 依賴途徑和CLV 依賴途徑來誘導WUS基因的表達,從而實現對莖端分生組織的調控;而WUS基因在異位表達時卻能夠激活CLV3的表達,導致莖分生組織增大;說明二者之間通過負反饋調節機制共同調控分生組織的發生[31]。WUS 蛋白從組織中心OC 區域轉移到CZ 區,在CZ區中它直接與CLV3啟動子結合影響CLV3的表達[32]。
3 生長素和細胞分裂素互作調控分生組織的發生與發育
生長素和細胞分裂素間的拮抗作用是頂端分生組織或側生分生組織器官發育起始的關鍵調控因素[24]。在植物分生組織的發生過程中,生長素、細胞分裂素的相互作用主要是通過WUS-CLV3的反饋調節機制、不同比例的CTK/IAA 水平、B-ARR 和A-ARR 介導的激素響應及運輸等一系列途徑來調控分生組織和相關器官的形態建成(圖2)。
在植物SAM 中,生長素通過影響STM的表達來上調細胞分裂素合成關鍵基因IPT7的表達,促進細胞分裂素的合成,并激活AHKs 途徑和細胞分裂素信號應答基因ARR7/ARR15以促進WUS-CLV3通路間接調控干細胞的增殖分化(圖2);WUS 可以直接抑制ARR7/ARR15的表達,是細胞分裂素信號的有效調節因子,還可直接調控細胞分裂素和生長素相關基因,控制生長素和細胞分裂素之間的平衡[33-34]。在植物RAM 中,MP 是激活QC 特異性WOX5基因和生長素響應基因PLT的調節因子,調節生長素信號介導分生組織模式的建立。另外,生長素可以通過單肽轉錄因子(monopteros,MP)介導的途徑來抑制細胞分裂素的生物合成,從而觸發器官原基的啟動;而MP/ARF5作為生長素信號的整合因子,使生長素信號和細胞分裂素信號在調控網絡中有機結合以參與SAM 的形成和維持[8]。
SAM 的形成與生長素的梯度和WUS表達水平密切相關,細胞分裂素的局部合成是細胞分裂素區域分布和WUS表達所必需的,是體細胞胚胎發生過程中SAM 形成的必要條件。細胞分裂素可以調控生長素的合成、運輸并在側原基起始時期擾亂生長素輸出載體PIN 的表達模式,從而間接調控生長素的合成。細胞分裂素B 類調節因子B-ARR 則通過抑制YUCCA基因的表達來降低生長素的積累;另外,B-ARR 還可以結合到生長素合成基因TAA1啟動子上,激活TAA1的表達從而調控生長素的生物合成[35]。
側生分生組織發育也受生長素和細胞分裂素共同作用調控。擬南芥葉腋處ATH1基因通過誘導分生組織STM的表達,維持了葉腋分生細胞的特征;另外,ATH1 蛋白能夠與STM 蛋白發生互作,形成一個STM基因的自激活回路[36]。ARR7/ARR15 在生長素的介導下負調控細胞分裂素的產生,PIN 蛋白介導的生長素運輸也會抑制細胞分裂素的產生,從而影響側生器官的形成。例如:TB1基因在水稻、玉米中對側枝的發育起負調控作用,主要通過調控生長素的運輸來調節生長素的水平,并參與腋分生組織的調節過程[37];在擬南芥葉腋處,隨著生長素和細胞分裂素水平的提高,SPS(supershoot)和BUS(bushy)基因的表達水平提高,并通過反饋調節相關激素水平來調控植株腋分生組織的發育[38-39]。
在RAM 中由AHK3/ARR1 介導的細胞分裂素響應信號途徑通過生長素抑制因子short hypocotyl 2(SHY2)負調控IPTs 的表達,從而影響細胞分裂素的合成,來促進根的伸長[40]。在側根分生組織發生過程中,維管柱內中柱鞘對生長素的運輸和信號感知的差異將引發不對稱的細胞分裂,從而形成側根原基,繼而形成側根。在這一過程中細胞分裂素通過抑制生長素轉運蛋白PINs 的表達以及生長素的梯度分布來負調控LRP 的發生[41]。CTK 通過依賴細胞分裂素受體AHKs 途徑介導細胞分裂素響應信號,控制分生組織的大小,并負調控PIN基因,從而促進側根形成[8]。擬南芥細胞分裂素B 型調節因子ARR1 介導RAM 調控以及側生分生組織的形成,ARR1 在中柱鞘激活SHY2的表達,以抑制生長素轉運蛋白PINs 的運輸,從而減緩側根原基的形成,并抑制側根分生組織的發生[42]。
4 其他植物激素對分生組織發生的影響
脫落酸(abscisic acid,ABA)除了在植物抗逆境和脅迫耐受性中發揮重要作用外,還可以通過脫落酸轉運蛋白NRT1、DTX50、ABCG25 等將ABA 運輸至植物生長發育的各個部位,從而實現對植物生長的調控[43]。在植物正常生長時,頂端形成的生長素促進相關誘導基因的表達和蛋白的抑制,而在頂端優勢去除之后,頂端不能合成足夠的IAA,細胞分裂素合成基因IPT開始表達并大量合成細胞分裂素,降低ABA水平,從而解除腋芽的休眠,促進芽的生長。
茉莉酸(jasmonate acid,JA)通過茉莉素轉運蛋白JAT1/ABCG16 的運輸使其在核內外形成濃度差,從而抑制主根的伸長[25]。茉莉素通過激活生長素生物合成基因anthranilate synthase a1(ASA1)的表達來調控生長素的主動運輸,從而控制側根的形成。而茉莉素響應因子MYC2 抑制生長素響應基因PLT1/2的表達,并調控IAA-PLT1/2 介導的RAM 的形成。另外,茉莉素可以誘導受傷部位快速積累生長素,從而改變損傷處的植物干細胞功能,并形成側原基[44],具體表現為MYC2 結合在PLT1/2的啟動子上抑制其表達,從而影響主根的伸長,促進側根的發育。同時茉莉酸調控轉錄因子ERF109可以促進生長素合成基因YUUC2的表達,從而促進生長素的生物合成以促進側原基的形成[45]。
赤霉素(gibberellin,GA)在分生組織發育中也發揮了重要作用。STM 可以激活莖端分生組織和側生分生組織邊界的GA2ox2(GA20-oxidases2)的表達,導致赤霉素的活性喪失,從而影響側生器官原基的形成[46]。在SAM 中KNOX1 抑制赤霉素的合成而促進細胞分裂素的合成以維持分生組織的大小,是植物分生組織發育的關鍵調控因子[47];相反的是,細胞分裂素通過刺激GA2ox2的表達并負調節KNOX1,而促進側生分生組織的發育[48]。
獨腳金內酯(strigolactones,SLs)作為一類由類胡蘿卜素衍生而來的新型植物激素,在植物根部產生,并沿莖干向上運輸,從而抑制植物的分枝發育。水稻D3/14/53(dwarf3/14/53)作為獨角金內酯的受體基因和F-box 家族基因的MAX(more axillary growth)一起參與SL 的生物合成和信號轉導。D3/314/53、MAX1/2/4則通過調控獨角金內酯的合成和轉導來抑制分枝的發育,D10編碼的類胡蘿卜素裂解雙加氧酶8(OsCCD8)受生長素的誘導,從而控制水稻側芽向外伸長,而OsMAX1則通過編碼OsCCD7來抑制水稻腋芽分生組織的生長[49]。
5 植物激素調控分生組織的展望
分生組織的維持和發育是器官形態建成的基礎,同時也是挽救植物機械損傷或自然損傷,以保證植物正常生長和作物產量的一種植物生存機制。在植物分生組織的發育過程中,生長素和細胞分裂素通過相互協同或者拮抗作用來共同調控分生組織的維持與發育,并且通過調控生長素/細胞分裂素的不同水平來實現對SAM、RAM 的促進或者抑制。在整個分生組織的發育過程中ABA、GA、JA、SLs 等通過協同或拮抗生長素和細胞分裂素的合成、運輸,響應信號途徑,從而實現對分生組織發育的調控。目前眾多研究已經揭示了植物激素在細胞增殖和新器官產生中的重要作用,初步明確了維持干細胞和植物器官發育的平衡機制,并且對于生長素和細胞分裂素的合成、運輸、信號轉導途徑及其調控頂端分生組織發育的機制已經比較清晰。
在植物分生組織發育的調控途徑中除了文中所述幾種激素的調控途徑外,還存在其他的信號調控途徑的協同作用。在植物的整個發育進程中,所有器官的產生都來自于分生組織內細胞的分裂、分化活動,并貫穿于植物生長發育的整個生長周期。因此,進一步明確頂端分生組織的調控方式對植物生長發育意義重大。植物體內生長素的梯度分布影響了側根、側芽的起始以及側根原基的發育,但生長素梯度的差異是如何轉化為不同的信號輸出并影響側生器官的形成還有待進一步研究;脫落酸是否受生長素表達水平的限制或誘導降解為非活性物質從而影響植物分生組織的形成,這一機制還不清楚;獨角金內酯除了調控植物側枝的形成外,還通過抑制植物下胚軸的生長來限制莖的伸長,它通過抑制生長素的合成來影響莖的負向地性的作用機制目前正在解析中。另外,植物激素調控分生組織形態建成的途徑僅在擬南芥等模式植物中研究得較為清楚,但對復雜植物組織分化現象的解釋還不夠,而更加深入研究植物激素調控分生組織發生及發育的分子機制,并將之擴展至各作物中去,將為進一步提升農作物產量和品質提供遺傳和理論支撐。
目前,利用野生近緣種培育多年生作物是解決農業生產和生態環境安全的重要手段,具有廣闊的應用前景。一年生作物(栽培種)收獲后,腋芽再生率或成活率低,表現為隨作物馴化過程慢慢被弱化,腋芽再生相關機制及野生種向栽培種的馴化機制還缺乏相關研究。分生組織的形成在作物馴化過程中受到人工選擇的影響,從而衍生了許多對分生組織調控途徑的研究,期待解析更多關鍵調控途徑來進一步提高生產效率和作物產量。因此,綜述植物激素對頂端分生組織發生和發育的調控,對未來研究植物頂端優勢的去除、植物腋芽的再生以及越冬休眠機制具有指導性的意義。
猜你喜歡
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2016年3期)2016-11-12 13:23:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
