土傳病原細菌的生存與致病權衡*
2022-06-09 01:16:00王佳寧江高飛王孝芳徐陽春沈其榮
土壤學報 2022年2期
關鍵詞:能力
韋 中,王佳寧,江高飛,王孝芳,徐陽春,沈其榮
(江蘇省固體有機廢棄物資源化高技術研究重點實驗室,作物免疫學重點實驗室,國家有機類肥料工程技術研究中心,江蘇省有機固體廢棄物資源化協同創新中心,南京農業大學資源與環境科學學院,南京 210095)
由土壤病原細菌引起的植物土傳病害日益引起人們的重視。常見的土傳病原細菌有引起番茄、煙草等茄科作物青枯病的青枯菌()、誘發豆科作物產生冠癭瘤的根癌農桿菌()和導致甘薯莖腐病的達旦提狄克氏菌()。土壤是植物土傳病原細菌生活史循環中的重要生境,當不存在寄主植物時,病原細菌可通過改變細胞狀態在土壤或水環境中存活數年之久;一旦感知到寄主植物信號,便會從土體向根際及寄主根系遷移,進而定殖根表和吸附植物皮層,隨后分泌致病因子穿透木質部,進入寄主體內繁殖,最終隨著寄主植物的死亡再次返回土壤環境。化學農藥、抗病品種和生物防治等手段盡管在病害防控中有較好的潛力,但對于土傳病害,其應用效果和穩定性通常受土壤條件的影響。由病害發生三要素可知,病原細菌的成功侵染離不開易感寄主和適宜病原菌發揮致病毒性特征的環境因子。在入侵寄主的過程中,土壤中的病原細菌會經受溫度、pH、含氧量、營養物質種類和數量等環境因素的驟變、土著微生物空間與資源競爭、原生動物捕食、噬菌體寄生以及寄主植物分泌抗菌物質等生物脅迫。為達到增殖和傳播的目的,病原細菌必須在不斷變化的環境中時刻協調生存和致病之間的權衡關系。了解土壤病原細菌適應環境變化的機制對于掌握其傳播、擴散和侵染規律有重要意義。
目前,人們對病原細菌與寄主植物的互作機制以及對病原細菌生存和致病能力的影響因素已經有大量的研究和總結,但是對于從土壤存活至根表入侵這個過程中土壤環境因素對病原細菌生存-致病權衡尚缺少系統的梳理和思考。本文首先總結了土傳病原細菌的生存-致病權衡規律和典型現象,歸納了土壤非生物和生物因子對病原細菌生存-致病權衡的影響;其次闡述了土傳病原細菌在植物根際入侵過程中的生存-致病權衡機制,并提出一些土傳病原細菌生存-致病權衡相關的科學問題。呼吁基于生存-致病權衡理論,建立降低環境中病原細菌豐度和毒性的土傳病害生態防控策略,為防控其他土傳病原體(如病原真菌)、實現綠色農業可持續發展提供理論參考。
1 土壤病原細菌的生存-致病權衡
1.1 病原細菌的生存-致病權衡規律
1859 年達爾文在《物種起源》中提到歌德的補償法則:“In order to spend on one side,nature is forced to economise on the other side”——自然在一處開支,就被迫在另一邊節省。不同特征間的負相關關系稱為權衡(Trade-off)。權衡效應在動植物領域得到廣泛的認可,研究人員認為動植物將有限的資源用于繁殖或者生存是造成權衡的原因,因此,權衡效應的核心是資源配置。權衡效應在微生物界同樣適用:一種細胞特性分配的資源多,那么其他特性配置的資源相對減少。微生物特別是病原微生物(病原細菌)具備諸多特性,如生物膜形成、代謝、運動性、毒性、DNA 修復能力以及對噬菌體、抗生素或其他脅迫的抗性等,而不同特性之間很可能存在負相關關系。微生物本身的細胞資源、膜蛋白結構和遺傳信息儲存及處理能力是有限性的,很難同時將不同特性最大化,因此在不同環境條件下表現出了一種特性增強而另一種特性減弱的權衡現象。病原細菌和非致病細菌的區別在于病原細菌能否分泌損害寄主健康的毒性因子,比如可堵塞植物輸水系統的胞外多糖、降解植物細胞壁的降解酶、破壞植物免疫的Ⅲ型效應蛋白等。除了與植物互作時發揮直接作用的毒性因子外,一些毒性特征的表達在整個植株發病過程中也十分重要,例如趨化性、運動性、鐵載體的分泌和生物膜的形成等決定著病原細菌能否成功入侵寄主根際。權衡是微生物適應環境的基本策略,生存-致病權衡對土傳病原細菌適應不斷變化的土壤-植物環境非常重要。根據權衡理論,當生存條件適宜,病原細菌不需要啟動高成本的抗脅迫機制,從而能將更多的資源分配給生長、繁殖、毒性因子等高成本的特性,進而提高病原細菌的致病性;當脅迫增強時,病原細菌必須提升其抵御脅迫能力以保證自身的生存,這通常伴有細胞結構改變導致的資源攝入能力降低、毒性基因丟失、毒性基因表達準確度的降低以及有害突變的增加等,即生存成本提高,這樣分配給生長、繁殖、毒性因子的資源相對減少,即表現為致病能力下降(圖1)。
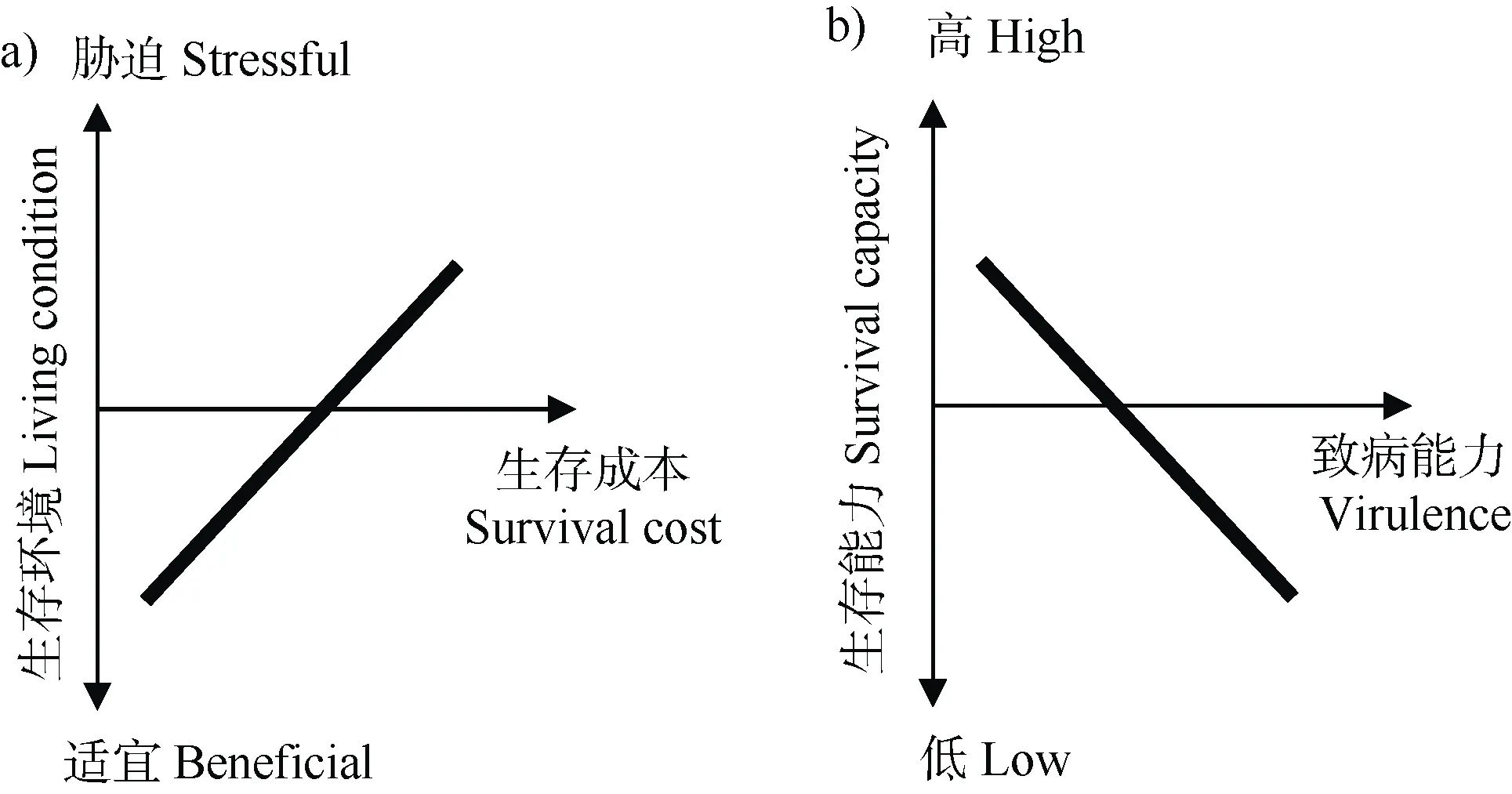
圖1 病原菌在不同環境中的生存成本(a)及病原菌的生存與致病的權衡規律(b)Fig.1 Survival cost of pathogens in different living environments(a)and the trade-off between survival capacity and virulence of pathogens(b)
1.2 土壤病原菌生存-致病權衡的典型現象
1.2.1 存活但不可培養狀態 當遭遇極端環境脅迫時,細菌會進入存活但不可培養狀態(viable but non-culturable,VBNC)。在這種狀態下,細菌的細胞完整,但呼吸、代謝活性降低,因此VBNC 被認為是細菌應對脅迫的一種自我保護機制。進入VBNC 狀態后,植物病原細菌會喪失致病能力,如青枯菌根癌土壤桿菌、十字花科黑腐病菌(pv.)、柑橘潰瘍病菌(pv.)和火疫病病原菌()等。VBNC 狀態的誘導因素有很多,如不適宜的溫度、pH、光照、氧氣不足、高鹽環境、干燥和重金屬脅迫等。一旦生存條件適宜或存在誘導物,病原細菌可從VBNC 狀態復蘇并恢復致病能力。據報道,4℃的低溫條件可誘導青枯菌進入VBNC 狀態,但當VBNC 狀態的青枯菌靠近寄主植物根系后能復蘇,且能繼續侵染植物。火疫病病原菌在硫酸銅的脅迫下進入VBNC 狀態;將VBNC 狀態的火疫病病原菌接種至寄主植物后,發現植物發病癥狀顯著低于未受脅迫的病原菌處理;不過復蘇后的病原菌的致病性卻與正常狀態下的病原菌致病力相當。由于檢測難度大,VBNC 狀態的病原菌是農業生產的一大隱患。
1.2.2 表型轉換 微生物會通過改變表型來增加種群多樣性,從而增強對環境的適應性。該現象常見于人類病原菌或病原真菌入侵寄主過程中,通常伴隨著毒性的改變。在土傳病原細菌中,野生型青枯菌在土壤、靜止液體和瓊脂平板的長期培養條件下菌落形態會發生變化,即從黏液態轉變為非黏液態(圖2)。這個過程稱為青枯菌的表型轉換(phenotypic conversion,PC),轉換后的青枯菌不能引起宿主發病。已經探明青枯菌的A 毒性網絡系統調控表型轉換,通過群體感應調控青枯菌生存與致病的權衡。在資源充足的情況下,青枯菌種群數量累積,當種群密度達到一定的閾值時啟動毒性系統表達,上調一系列毒性因子,如EPS(胞外多糖)、植物細胞壁降解酶、Ⅲ型分泌系統等。但當資源有限時,種群數量降低,受抑制,毒性減弱,但是資源利用能力、運動能力和種群增長速率均增強。有研究表明,在適宜條件下,PC 突變體還可轉變為野生型菌株,從而恢復其致病能力。但是這種現象在自然環境下尚未見報道。
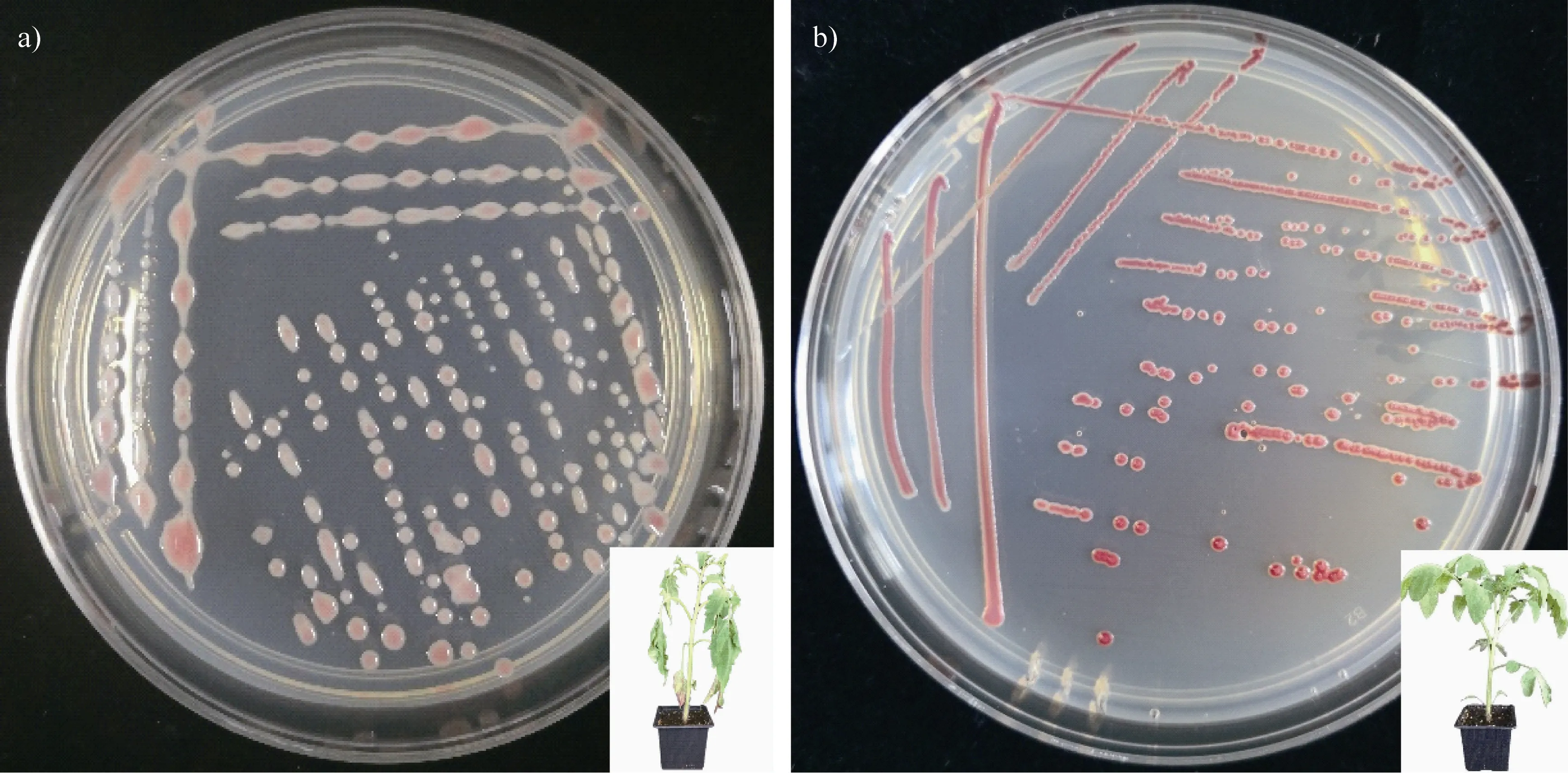
圖2 青枯菌的表型轉換:a 為黏液型青枯菌,一般致病性強;b 為非黏液型青枯菌,一般喪失致病性Fig. 2 Phenotype conversion of Ralstonia solanacearum. Mucoid and virulent colonies(a);Non-mucoid and avirulent ones(b)
2 驅動病原菌生存-致病權衡的主要土壤因素
2.1 非生物因素脅迫
土壤非生物因子的變化對土壤病原細菌生存與致病能力的影響較大。眾所周知,土壤溫度、養分濃度、pH、水分和氧氣條件等會隨著季節、天氣或是有無植物而發生瞬時劇烈或漫長微弱的改變。在遭遇非生物脅迫時,微生物將細胞資源優先分配給高成本的脅迫應激保護機制,削減細胞增殖所需資源。對病原細菌而言,對自我保護機制的高投入可能會導致減少甚至停止毒性因子的表達。例如,因環境溫度過低而進入VBNC 狀態時,病原細菌對pH、重金屬、高溫、抗生素等物理和化學脅迫的抗性水平顯著提高。此時,病原細菌雖然具備致病的潛力,但不能引起寄主發病。養分資源匱乏也是迫使病原菌權衡生存和毒性表達的常見環境脅迫因素。攜帶和表達毒性因子成本高昂,當營養資源匱乏時,病原細菌可能啟動寡營養物質運輸系統,以增加在群落中的優勢,比如無毒性的PC 型在貧營養條件下較有毒野生型的青枯菌生長更快,對鐵離子的利用效率更高。病原細菌也能通過丟失致病基因來增加在貧營養條件下的競爭力。若養分資源極度匱乏不足以支持穩定生長時,大多數微生物會停止分裂而進入生長停滯階段。環境脅迫過強可能會破壞遺傳信息的穩定,從而導致毒性基因難以表達。例如氧化、酸性和高溫脅迫條件會影響DNA 結構的穩定性,降低達旦提狄克氏菌的主要毒性基因——果膠裂解酶基因的表達。
2.2 生物因素脅迫
病原細菌在土壤中應對寄生、競爭和捕食等生物脅迫提升生存能力的同時,可能會被迫調整自身的致病能力。例如,細胞表面鞭毛是常見的噬菌體受體,同時也是病原菌的重要致病因子。面對噬菌體裂解侵染脅迫,微生物會通過修飾、丟棄、改變其細胞表面的噬菌體受體,提高對噬菌體的抗性,但鞭毛致弱導致運動能力降低,進而降低其致病能力。此外,一些病原菌的噬菌體受體與吸收營養物質有關,所以通過破壞受體而獲得噬菌體抗性的突變體會降低其營養吸收,從而減弱其競爭能力。同營養級內的其他微生物不僅可通過爭奪營養和空間間接影響病原菌的致病能力,還能產生抗生素類物質直接影響病原菌的生存。病原細菌的一些抗性一般是通過染色體突變獲得,并涉及關鍵基因的修飾,以及質粒復制和質粒基因的表達等影響其代謝水平,最終導致抗性菌株生長速率下降、競爭能力減弱;另一些抗性是通過改變細胞膜滲透性獲得,如大腸桿菌孔蛋白通道能同時滲透營養和抗生素,降低抗生素滲透速率、提高抗生素抗性的同時,其資源攝取速率也隨之降低,導致生長減緩。此外,土壤中的有益菌還能通過分泌的揮發性有機物(volatile organic compounds,VOCs)抑制青枯菌運動能力、根際定殖、生物膜形成和氧化脅迫應答能力,這可能是由于VOCs 脅迫導致青枯菌的生存成本提高而毒性因子減弱。為應對原生動物等捕食脅迫,病原菌會增強抗捕食特征,這可能會導致致病相關的特征減弱,如運動能力和資源利用能力下降,從而導致致病能力被削弱。
3 土傳病原菌根際入侵過程中的生存-致病權衡
土傳病原菌侵染植物的第一步是感知寄主并從土體移動至根際,同時要與根際土著微生物競爭資源和空間,才能在根際成功定殖。當根際種群達到一定數量后,病原菌啟動毒性基因的表達,從而入侵植物根系。所以在入侵植物體內之前,病原菌就需要啟動一些幫助其成功侵染寄主植物的毒性因子以及應對一系列的生存脅迫(圖3)。
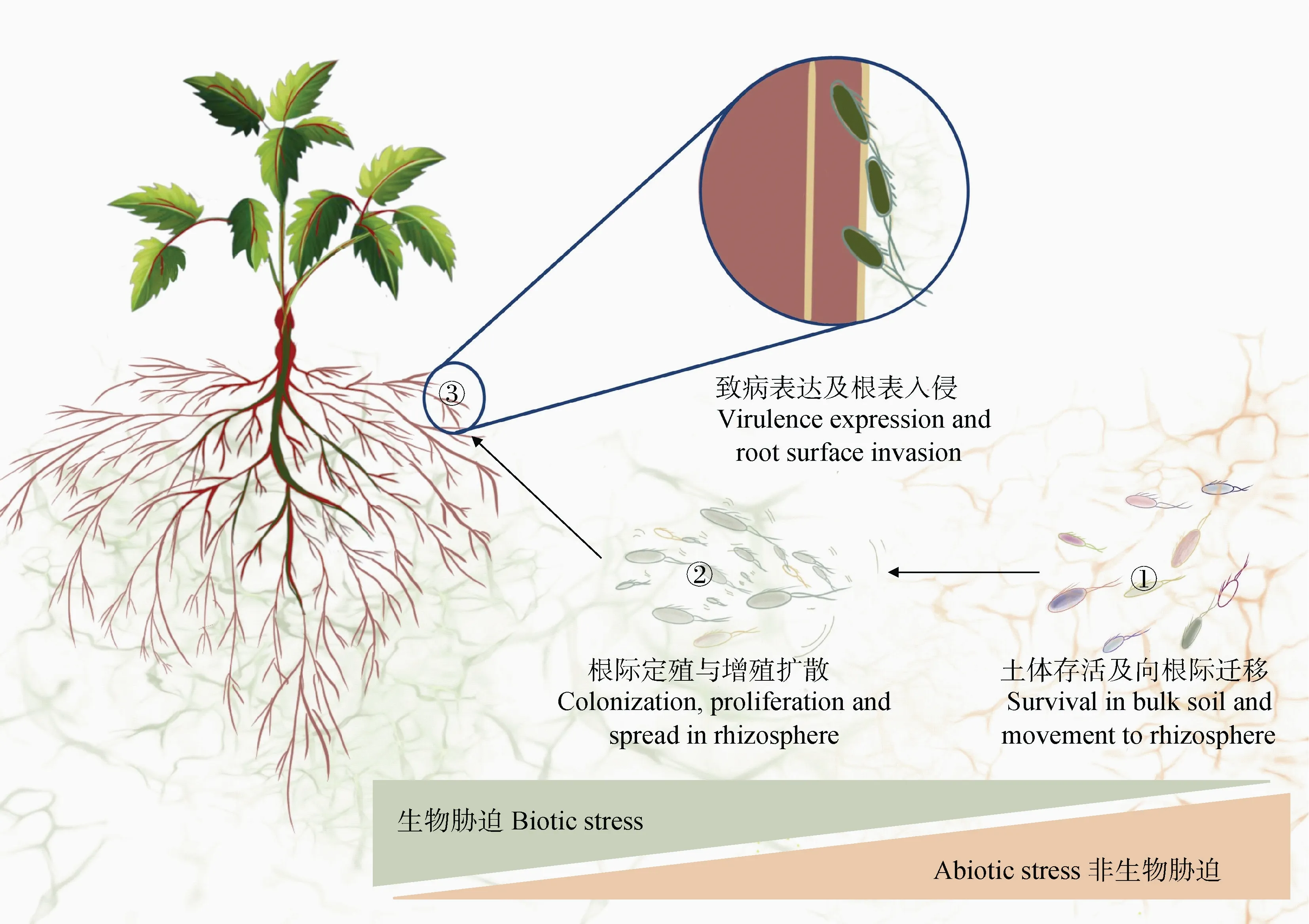
圖3 土傳病原菌在土壤中生存、定殖和侵染的過程Fig. 3 Survival,colonization and infection processes of soil-borne pathogen in soil
3.1 從土體向根際遷移過程的生存-致病權衡
在土壤中,低溫、干燥和養分貧瘠的土壤環境會淘汰大量抗脅迫能力低下的病原菌個體,VBNC、饑餓細胞、轉換成抗脅迫表型和形成生物膜的細胞可以存活下來。一旦感知到根系分泌物等植物信號時,病原菌利用趨化性向寄主植物根際的方向遷移。趨化作用通常具有寄主特異性,例如青枯菌能被幾種特定的氨基酸和有機酸寄主植物信號吸引,而水稻等寄主植物的根系分泌物不能引起青枯菌的定向遷移。趨化作用和運動能力密不可分,微生物靠鞭毛或菌毛運動,青枯菌在鞭毛驅動下向根際移動并通過根系進入植物維管束系統。青枯菌的突變體缺少鞭毛細絲,突變體不能激活鞭毛基因的表達,這兩種突變體完全不能移動也不能朝植物根系運動,所以在土壤中它們的致病能力較野生型青枯菌顯著降低。病原細菌對脅迫抗性的提高可能會以犧牲驅化運動能力為代價。細菌普遍會通過形成生物膜來提高重金屬或抗生素等脅迫的抗性,對青枯菌趨氧性的研究表明,喪失趨氧能力的突變體青枯菌能形成更厚的生物膜,而喪失趨氧性的青枯菌幾乎無運動能力。因此,致弱土壤中的病原菌可通過添加可降低病原菌運動能力的生物脅迫或非生物脅迫,比如已有研究發現,根際促生菌——解淀粉芽孢桿菌產生的易揮發性有機物VOCs 脅迫可使青枯菌的基因表達下調,降低青枯菌向根際遷移和定殖的能力。
3.2 根際定殖與增殖過程的生存-致病權衡
到達寄主植物根際后,病原菌需要與土著微生物進行營養競爭和拮抗競爭以實現在根際的成功定殖。在根際環境中,能夠高效利用有限資源的微生物通常會占據競爭優勢。比如在根際限鐵的條件下,能否產生鐵載體以獲取鐵離子對狄克氏菌和青枯菌等病原菌的根際入侵至關重要。細菌間的拮抗競爭作用也會通過增加病原菌的生存脅迫而降低其毒性。因此,利用病原菌的拮抗菌可達到致弱病原菌毒性的效果。劉波等發現短小芽孢桿菌()ANTI-8098A 可作為青枯病的生防菌,因為ANTI-8098A 處理后的青枯菌會喪失致病能力,即使回接至番茄植株上也不能致病。這可能是由于強致病力菌株對拮抗菌的拮抗物質抵御能力弱,而弱致病力的菌株對拮抗菌拮抗物質的抵抗能力強,因此,無致病力菌株在拮抗菌的脅迫下種群優勢明顯,從而表現出青枯菌整體致病能力的減弱。根際是微生物聚集的熱區,病原菌不僅與同營養級微生物競爭,而且頻繁地遭遇噬菌體的寄生和原生動物的捕食,從而降低其在根際的適應性。因此,向根際加入病原菌專性裂解性噬菌體或捕食病原菌的原生動物可降低病害的發生。Wang 等發現噬菌體組合處理根際病原青枯菌數量顯著降低,并且噬菌體組合多樣性越高,青枯菌成功突變抵御噬菌體的概率越低。而成功進化出噬菌體抗性的青枯菌,生長顯著減緩,再次侵染作物根系的能力也顯著降低。Xiong 等發現發病植株中吞噬型原生動物的相對豐度隨植物生長而顯著降低,而這一降低與青枯病的爆發相吻合。青枯病爆發的原因除了由于根際原生動物捕食脅迫減弱導致根際青枯菌數量增加外,也可能存在青枯菌在抵御原生動物捕食機制上投入減少、致病能力增強的情況。
3.3 根表入侵過程的生存-致病權衡
根際成功定殖后,病原細菌需要吸附在植物的根表來完成根系入侵。鞭毛不僅能增強細菌的運動能力,還能增加病原菌對寄主植物根表細胞的黏附作用,比如鞭毛有助于病原菌的黏附能力。粘附至根表后,病原細菌會釋放一系列毒性因子以助其進入寄主體內,但也伴隨著相應的生存成本的提高。根癌農桿菌的Ⅳ型分泌系統分泌的一種毒性效應蛋白可誘導植物產生有利于自身生長的物質(冠癭堿),但是該蛋白的表達會降低自身的適應性,所以當冠癭堿不足時,致病型根癌農桿菌的競爭能力顯著低于不攜帶該致病基因的菌株。對于病原菌的入侵,植物也會產生物質抑制病原菌生長,比如植物產生的羥基香豆素能抑制青枯菌生物膜的形成和Ⅲ型分泌系統效應蛋白的轉錄。植物還會產生抗菌多肽和次級代謝產物,比如羥基肉桂酸(HCA)、類黃酮、類異戊二烯和植物堿。青枯菌必須分泌能夠降解HCA 的相關蛋白才能免受其毒害,當青枯菌的HCA 降解蛋白相關基因缺陷時,其根系定殖效率大大降低。此外,青枯菌還需要多藥物外排泵將抗菌化合物排出體外,使其免受植物分泌的抗生物質的毒害。
4 結論與展望
土傳病原菌侵染作物是一個時空動態過程,涉及了病原菌與土壤生物、非生物因素的互作。土壤環境本身以及病原菌與環境互作的復雜性,是制約土傳病害防控效率的關鍵因素。掌握病原菌在土壤中的生存與致病權衡的規律和機制,對建立土傳病害的精準防控策略具有重要意義。盡管本文梳理了土壤病原細菌的生存與致病權衡規律、影響因素和作用機制,但相關研究證據尚亟待豐富。近期,土壤學家呼吁土壤學工作者關注土壤病原菌的研究,病原菌在土壤中的生存與致病權衡可作為土壤學工作者涉足土壤病原菌研究的一個重要切入點。針對土壤病原菌的生存與致病權衡,以下幾方面研究值得關注。
(1)深入探究生物和非生物環境因素對病原菌存活和致病能力的影響,以及對病原菌生存-致病權衡的驅動作用;加強土壤病原菌的生態型和基因型多樣性的研究及環境對病原菌多樣性的影響;建立原位條件下土壤病原菌生存、根際遷移、入侵致病過程中病原菌表型-基因型的關系模型。
(2)以往的研究側重于單一脅迫下生存-致病的權衡規律,忽略了環境本身的復雜性,導致相關理論成果不能有效指導實踐。微生物與病原菌間的互作結果通常受到多種非生物因素的影響,因此在利用生物脅迫防控土傳病害時需加強根際微生物網多級互作認識,需要厘清生物和非生物環境因素疊加作用對土壤病原菌生存與致病權衡的影響機制研究,構建以生存和致病權衡理論為指導的綜合生態防控體系。
(3)加強對病原菌適應性進化分子機制的研究。研究脅迫條件下病原菌的堿基置換、插入、缺失等響應,結合全基因組關聯分析,以探究生存-致病權衡的分子機制。除此之外,應加強生態學、微生物學、進化生物學、基因組學、轉錄組學等多學科、多維度聯合揭示病原菌的生存與致病權衡規律和作用機制,以獲得對病原菌致病-生存權衡的準確認識,從而精準高效阻控土傳病害。
衷心感謝南京農業大學微生態與根際健康實驗室張耀予為本文圖3 提供插圖。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
中學生數理化·七年級數學人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(2018年6期)2018-08-16 07:23:10
新高考(英語進階)(2018年1期)2018-04-18 14:00:11
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
