氮素水平對土壤甲烷氧化和硝化微生物相互作用的影響*
2022-06-09 01:16:58孟春梅劉杏梅諸葛玉平賈仲君邸洪杰徐建明
土壤學報 2022年2期
關鍵詞:水平
潘 紅,李 勇,孟春梅,鄭 燕,劉杏梅,諸葛玉平,賈仲君,邸洪杰,徐建明
(1. 浙江省農業資源與環境重點實驗室,浙江大學環境與資源學院,杭州 310058;2. 土肥高效利用國家工程研究中心,山東農業大學資源與環境學院,山東泰安 271018;3. 鄭州輕工業大學食品與生物工程學院,鄭州 450002;4. 中國科學院南京土壤研究所,南京 210008)
甲烷(CH)和氧化亞氮(NO)是兩種重要的溫室氣體,其全球增溫潛勢分別是二氧化碳(CO)的28 倍~34 倍和265 倍~298 倍。旱地土壤(森林土壤、草原土壤等)是大氣甲烷的有效匯,因其具有能夠氧化大氣濃度甲烷的高親和力甲烷氧化單加氧酶(MMO)。好氧土壤中,NO 的主要產生途徑是有氧硝化作用。氨氧化過程是有氧硝化的第一步,也是限速步驟,主要由具有氨單加氧酶(AMO)的氨氧化古菌(AOA)和氨氧化細菌(AOB)完成。研究發現,MMO 在進化上和AMO 密切相關,它們有高度相似的氨基酸序列、相似的蛋白復合體結構、相似結構的底物(CH和NH)、相似的被抑制特性和相似的生態位,而且都競爭N 源。甲烷氧化細菌(MOB)的生長繁殖需要氮源,但其自身不具備固氮能力,因此,MOB 需要獲取外源氮源。鑒于以上研究背景,甲烷氧化和硝化過程有著必然的內在聯系。
草原生態系統是世界上最大的陸地生態系統,草地面積占我國國土面積的41%。草原土壤既是CH的匯,又是NO 的源,因此,有關草原生態系統中土壤的碳氮循環一直是研究熱點問題。當前關于草原土壤系統中硝化過程和甲烷氧化過程的研究發現,AOB 和MOB 分布廣泛,在多種草原土壤生境中參與硝化和甲烷氧化過程。旱地土壤類群(USC)MOB 因其具有高甲烷親和力在草原土壤中的重要性逐漸被發現。但目前關于草原土壤中不同氮水平下甲烷氧化活性和相關活性微生物,以及甲烷氧化和硝化之間復雜的相互作用關系機制的研究較少。因此本研究選取內蒙古錫林郭勒盟草原土壤為研究對象,結合穩定同位素核酸探針(DNA-SIP)、實時熒光定量PCR和Miseq 測序等分子生物學技術,分析不同氮水平下,甲烷氧化和硝化微生物的相互關系,闡明草原生態系統CH氧化和硝化的交互作用機制,以期為深入了解草原土壤碳氮循環、提高草原氮肥利用率、增強草原生產力、實現溫室氣體減排提供科學依據。
1 材料與方法
1.1 土壤樣品概況
土壤樣品采自位于內蒙古錫林郭勒盟草原(43°26′—44°39′N,115°32′—117°12′E)的中國科學院內蒙古草原生態系統定位研究站。該地區冬季寒冷,夏季涼爽,屬于半干旱大陸性季風氣候,每年大約102~136 d 無霜期,全年平均氣溫-0.4 ℃,年均降水量350 mm 左右,降雨多發生在六月至八月,雨熱同期,季節分布不均,與植物生長期一致。主要植被類型包括羊草()、大針茅() 和 粗 隱 子 草()。土壤類型為暗栗鈣土。該樣地1996 年以前自由放牧,自1996 年起禁牧圍封。樣地大小為400 m × 60 m,包括5 個小區,每個小區70 m × 60 m,相鄰的兩個小區之間有10 m 的緩沖帶便于取樣。2016 年8 月用5 cm 直徑土鉆按照“S”型取樣法采集0~20 cm 表層土,全部混勻,研磨過2 mm 篩。土壤基本理化性質如下:pH 7.22,容重1.32 g·cm,有 機 質 25.47 g·kg,全 碳 16.67 g·kg,全 氮1.50 g·kg, 硝 態 氮 63.36 mg·kg, 銨 態 氮4.45 mg·kg,有效磷(Olsen P)7.31 mg·kg,速效鉀 251.10 mg·kg。
1.2 微宇宙培養實驗
將相當于 6.0 g 干重的新鮮土壤樣品轉移至120 mL 血清瓶,調節40%田間持水量,膠塞加鋁蓋封口,25 ℃黑暗中預培養,每周監測CO濃度,當濃度降至5 000 μL·L時開始正式培養。
正式培養實驗共設7 個處理,如表1 所示,每個處理3 個重復。CH處理(99 atom%C,Sigma-Aldrich,St Louis,MO,USA)用以分析甲烷氧化活性和活性甲烷氧化微生物;CO+Urea(99 atom%C,上海穩定性同位素工程技術研究中心,上海,中國)處理用以分析硝化活性和活性硝化微生物;CH+CO+Urea 和CH+CO+Urea處理用以分析甲烷氧化和硝化之間的相互作用。其中,添加尿素的處理均包括N 20 μg·g干土(U20)和100 μg·g干土(U100)兩個氮素水平。試驗流程如圖1 所示,每周用高純空氣(20% O,80% N)換氣1 min,保證有氧環境,同時以溶液的形式逐滴加入尿素溶液,調節60%田間持水量,血清瓶膠塞加鋁蓋密封后加5%的CO。其中甲烷濃度動態監測,隨時添加,保證1%濃度。在25 ℃生化培養箱中黑暗培養21 d,分別在0、7、14、21 d 進行破壞性取樣。所取樣品分兩部分,2 g 左右保存在-80 ℃冰箱,用以提取土壤DNA;剩余樣品用以分析硝化活性。
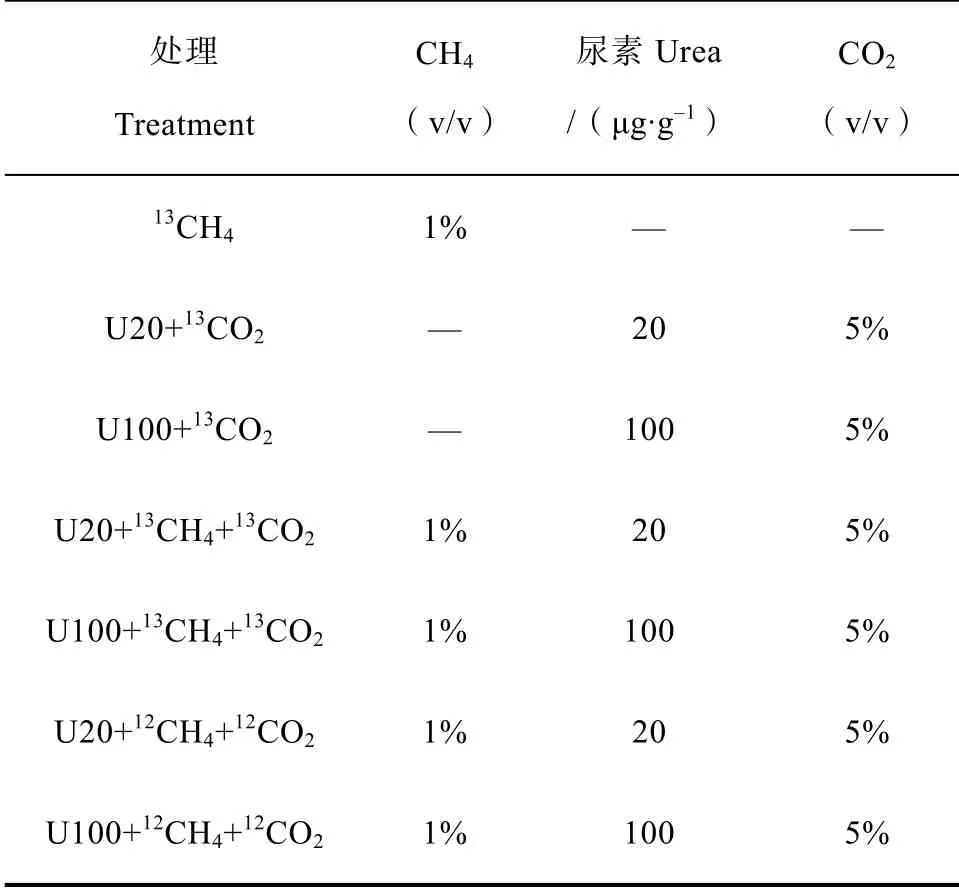
表1 微宇宙培養試驗處理概況Table 1 Schedule of the treatments
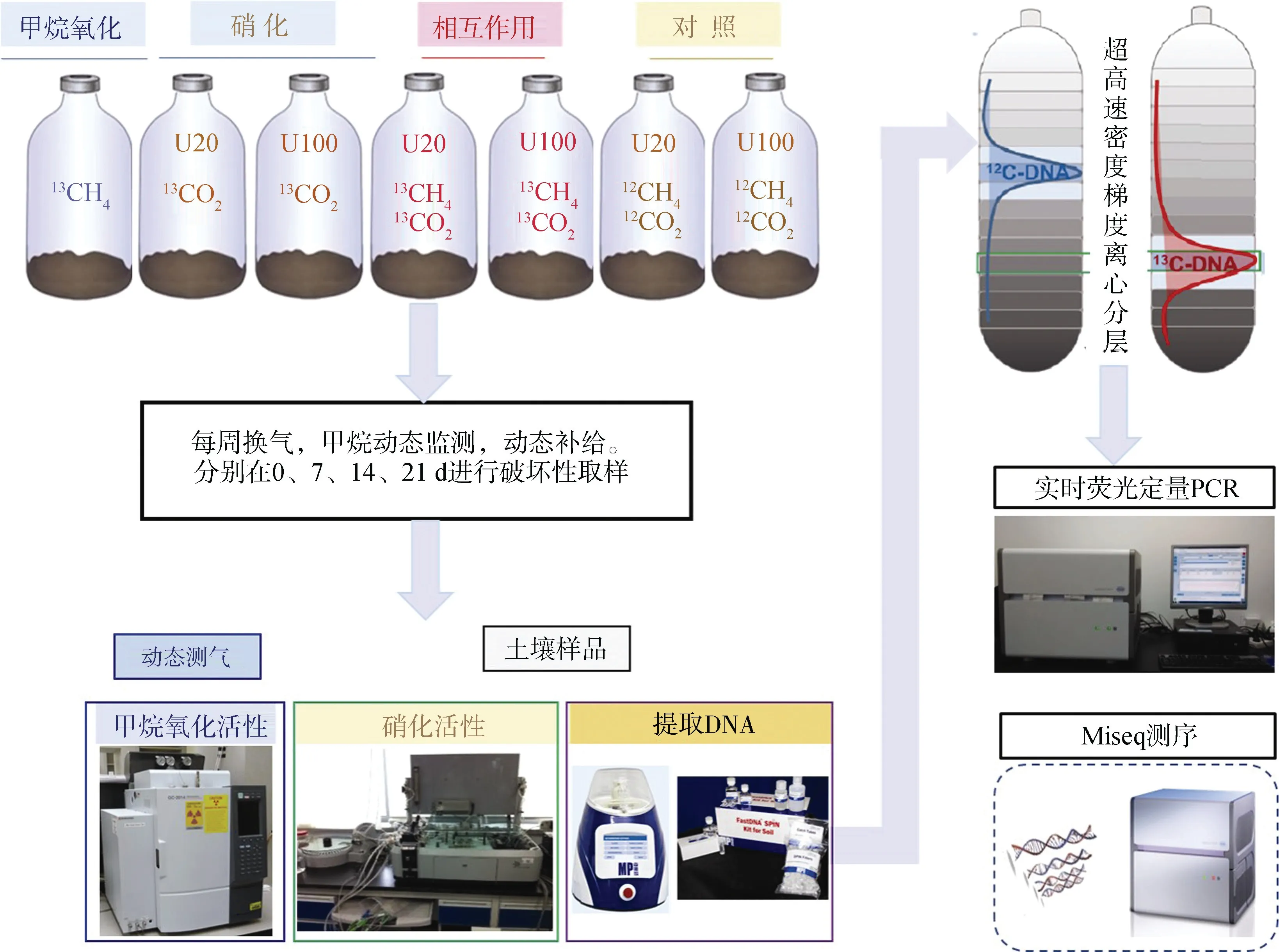
圖1 實驗流程圖Fig. 1 The experimental procedure
1.3 DNA 提取和SIP 分層
將0 d 和21 d 的土壤樣品用FastDNA Spin Kit for Soil(MP)提取土壤基因組總DNA。用0.7%的凝膠電泳檢測DNA 的完整性,NanoDrop? ND-2000 UV-Vis spectrophotometer(NanoDrop Technologies,Wilmington,DE,USA)檢測DNA 的質量和純度。DNA 樣品-20 ℃冰箱保存,用以進行超高速等密度梯度離心。
SIP 分層方法參考文獻[13]。具體操作方法如下:大約3 μg DNA 與4.9 mL 氯化銫(CsCl)溶液混合,調節浮力密度至1.725 g·mL。然后將混合液轉移至5.1 mL 超高速離心試管中,調平密封后用貝克曼Vti65.2 轉子(Beckman Coulter Inc.,Palo Alto,CA,USA)在20 ℃條件下以177 000×的速度離心44 h。離心結束后,利用NE-1000 注射器泵(New Era Pump Systems Inc.,Farmingdale,NY,USA)將新滅菌蒸餾水以穩定流速(0.38 mL· min)從上向下將溶于CsCl 中的DNA 置換到15 個1.5 mL 離心管中,即每個土壤DNA 樣品分層后總共得到15層DNA 組分溶液,每層體積為380 μL,取65 μL用手持式AR200 折光率儀(Reichert Inc.,Buffalo,NY,USA)測定折光率。將分層后的DNA 組分溶液用PEG6000 和乙醇純化,溶于30 μL 新滅菌蒸餾水進行后續分析。
1.4 甲烷氧化菌pmoA 和氨氧化菌amoA 基因的定量PCR
將0 d 和21 d 的總DNA 以及上述分層純化后的DNA 樣品進行和基因的定量PCR分析(LightCycler 480,Roche Applied Science),具體引物信息和PCR 條件見表2。反應體系20 μL,包括10 μL SYBR Premix Ex Taq(TaKaRa,Dalian,China)、1.5 μL DNA 模板(1~10 ng)、0.79 μL milli-Q水,體系中引物終濃度0.5 μmol·L。最后跑溶解曲線,利用 50~99 ℃的升溫連續檢測熒光以確認PCR產物的特異性。本實驗定量PCR 擴增效率在80.5%~108.3%之間,值在0.990~0.998 之間。

表2 本試驗中引物信息及PCR 條件Table 2 Primers and conditions used in this study
1.5 Miseq 測序、系統發育分析和共線性網絡分析
將0 d 和21 d 的總DNA 以及上述分層純化后的重層DNA 樣品進行Miseq 高通量測序。利用Illumina?MiSeq 測序儀(Illumina,San Diego,CA,USA)對上述DNA 的16S rRNA 基因的V4 區進行測序,引物序列及擴增條件見表2。本實驗中,只有序列長度大于200 bp,平均質量大于25,沒有模糊噪聲,至少80%符合16S rRNA 序列的結果才會進行后續分析。在97%序列相似度水平上進行OTU(可操作分類單元)聚類,最后參考Silva 數據庫對OTU 進行物種注釋。
總DNA 樣品共得到1 149 320 條高質量序列,平均每個樣品得到47 888 個高質量序列。其中屬于細菌的序列有1 123 384 條,占97.74%;屬于古菌的序列有25 936 條,占2.26%。重層DNA 樣品在CH、U20+CO、U100+CO、U20+CH+CO和U100+CH+CO處理下分別得到426 842、407 179、429 748、501 376 和480 393 條高質量序列。
選取屬于MOB、AOA、AOB 和NOB 的序列,用Molecular Evolutionary Genetics Analysis(MEGA 7.0)分別進行系統發育分析。以重層MOB、AOB 和NOB 的OTU 豐度表為原始數據,采用Spearman 相關系數模型,顯著性< 0.05,通過CoNet 軟件構建共線性網絡圖,Gephi 0.9.2 對網絡圖進行可視化分析。網絡圖包括節點和邊,各節點代表不同的物種,以顏色區分。邊代表物種之間的相互作用,紅色和綠色分別代表正相關和負相關,線的粗細表示相關性的大小。將總DNA 和重層DNA 的測序數據上傳至NCBI 的Sequence Read Archive(SRA)數據庫,檢索號分別為PRJNA611699 和PRJNA611713。
1.6 數據處理
硝化活性、甲烷氧化活性、/基因豐度和相對豐度的顯著性差異分析通過SPSS 20.0 單因素方差分析(one-way ANOVA)的Duncan 多重檢驗完成;作圖通過Origin 9.0 完成;< 0.05 表示差異顯著。
2 結 果
2.1 甲烷氧化活性和甲基營養微生物
氮素水平對甲烷氧化活性和相關微生物豐度均有顯著影響(圖2),經過21d 的微宇宙培養,相比不施氮(U0),甲烷氧化活性、基因豐度和甲基營養微生物相對豐度在低氮水平(U20)下顯著增加,在高氮水平(U100)下顯著降低。在U0、U20 和 U100 三個處理下,甲烷氧化活性分別是37.79、45.27 和16.11 μmol·g干土,基因豐度分別是3.08×10、4.24×10和9.50×10copies·g干土。甲烷氧化活性、基因豐度以及甲基營養微生物的相對豐度均在不同氮水平下呈現出“低促高抑”的規律(圖2)。
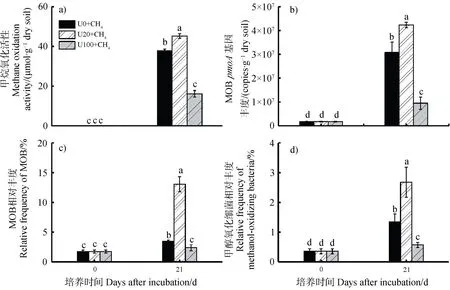
圖2 甲烷氧化活性(a)、甲烷氧化細菌pmoA 基因豐度(b)和甲基營養微生物相對豐度(c~d)分析Fig. 2 Changes in methane oxidation activity(a) and quantitative distribution of methylotrophs(b-d) in soil microcosms over an incubation period of 21 days
2.2 硝化活性和硝化微生物
凈硝化速率、硝化微生物豐度和相對豐度如圖3 所示。U20、U20+CH、U100 和U100+CH的凈硝化速率分別為51.57、34.60、163.50 和165.02 μg·g干土(圖3a);AOB基因豐度分別為1.90×10、1.23×10、6.34×10、8.04×10copies·g干土(圖3b)。低氮水平下,CH的存在顯著抑制硝化活性和AOB基因豐度;而高氮水平下,CH的存在對硝化活性和AOB基因豐度沒有顯著影響。AOB 和NOB 的相對豐度也符合該規律(圖3d,3f)。AOA基因的豐度和相對豐度在培養21 d 之后顯著降低,甲烷存在與否對AOA基因豐度影響不顯著(圖3c,圖3e)。
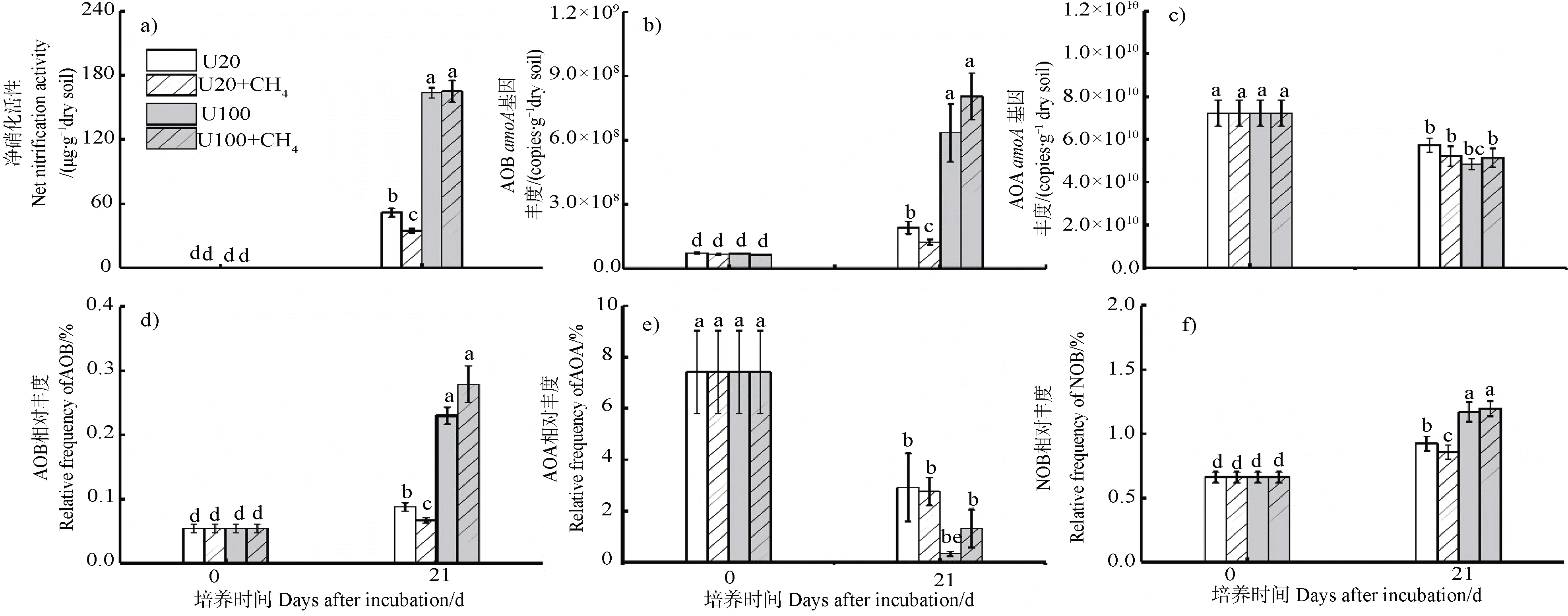
圖3 凈硝化活性(a)和氨氧化微生物amoA 基因豐度(b~c),硝化微生物相對豐度(d~f)分析Fig. 3 Net nitrification activity (a) and quantitative distribution of nitrifiers (b-f) in soil microcosms over an incubation period of 21 days
2.3 DNA-SIP 技術鑒定活性甲烷氧化和硝化微生物
離心后每個樣品分為 15 層,浮力密度在1.676~1.791 g·mL之間。對每一層進行和基因的定量PCR 分析,然后將每一層的分析結果與各層次中最大的基因豐度作比,得到和基因在各浮力密度梯度中的分布圖(圖4a)。甲烷氧化菌在不施氮處理中被顯著標記,低氮處理增加了MOB 被標記的程度,而高氮處理則降低了MOB 的標記程度。AOB 在低氮和高氮水平下均被顯著標記,尤其是在高氮水平下的標記更顯著。低氮水平下,甲烷的存在抑制AOB 的被標記程度。低氮和高氮水平下,AOA 均沒有被標記。根據定量分布結果選取重層DNA(3~12 層)進行高通量測序,選擇分別屬于MOB、AOB 和NOB 的OTU 繪制各浮力密度上的相對豐度分布圖,結果如圖4b、圖4c。MOB 和AOA 的相對豐度在各浮力密度的分布趨勢和上述定量分析的結果完全一致。AOB 和NOB 的相對豐度分布圖顯示低氮水平和高氮水平下,甲烷的存在對AOB 和NOB 的被標記程度均有抑制效果。

圖4 甲基營養微生物和硝化微生物豐度和相對豐度在不同浮力密度上的分布Fig. 4 Quantitative distribution of methanotrophs as well as nitrifiers across the entire buoyant density gradient of the fractionated DNA from soil microcosms after incubation for 21 days
2.4 活性甲烷氧化微生物和硝化微生物的群落結構
如圖5 所示,草原土壤99.7%以上的MOB 屬于甲基桿菌屬();活性MOB 包括甲基桿菌屬()和甲基暖菌屬(),以為主,占90%以上。活性MOB 在U0+CH、U20+CH和U100+CH的處理中所占的比例分別為92.62%、96.35%和91.79%,活性MOB 所占的比例分別為7.38%、3.65%和8.21%。硝化微生物AOB 主要包括亞硝化球菌屬()、亞硝化螺旋菌屬()和亞硝化單胞菌屬(),分別占72.00%、26.00%和2.00%。65%以上的活性AOB 為,其在U20、U20+CH、U100、U100+CH處理中對AOB 的占比分別為65.33%、80.23%、78.30%和82.10%;在AOB 中的占比分別為28.41%、10.13%、20.77%和16.13%;在AOB 中的占比分別為6.25%、9.64%、0.93%和1.77%。該草原土壤中的亞硝酸鹽氧化細菌(NOB)均隸屬于,包括、和三個種。活性NOB 以為主,其在U20、U20+CH、U100、U100+CH處理的占比分別是:71.3%、61.9%、 78.4%和78.8%;的占比分別為:23.6%、27.7%、16.9%和 14.5%;的占比分別為5.1%、10.4%、4.7%和6.7%。
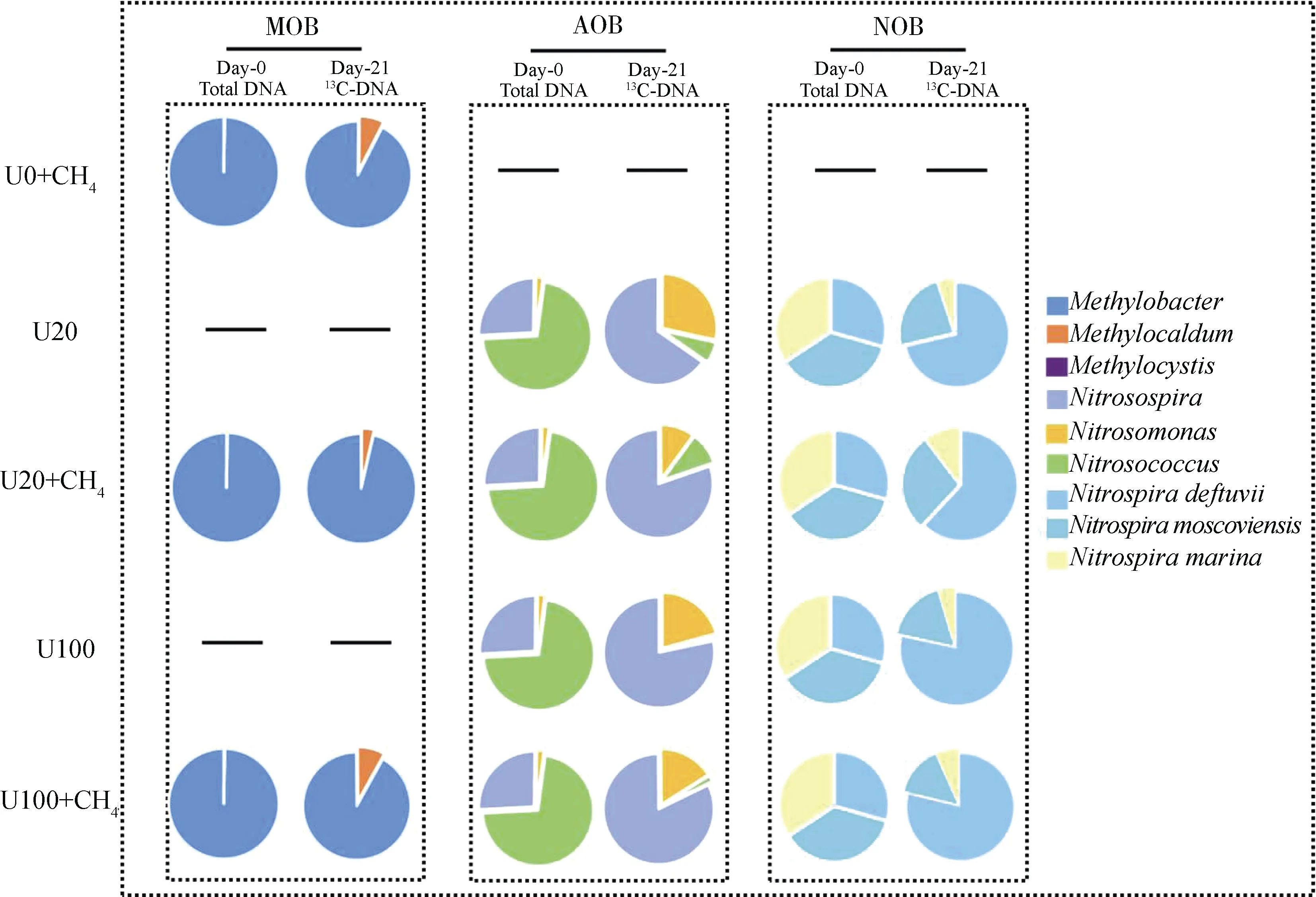
圖5 活性甲烷氧化微生物和硝化微生物的群落組成分析Fig. 5 Proportional changes of active methanotrophs and nitrifying phylotypes in SIP microcosms after an incubation period of 21 days
2.5 活性甲烷氧化微生物和硝化微生物的共線性網絡
為明確活性甲烷氧化微生物和硝化微生物的相互作用,將重層MOB、AOB 和NOB 的高豐度OTU構建共線性網絡圖(圖6)。結果發現,MOB 與AOB 存在顯著的負相關關系,此外,MOB 還與NOB 存在顯著的負相關關系,AOB 與NOB 存在顯著正相關關系。
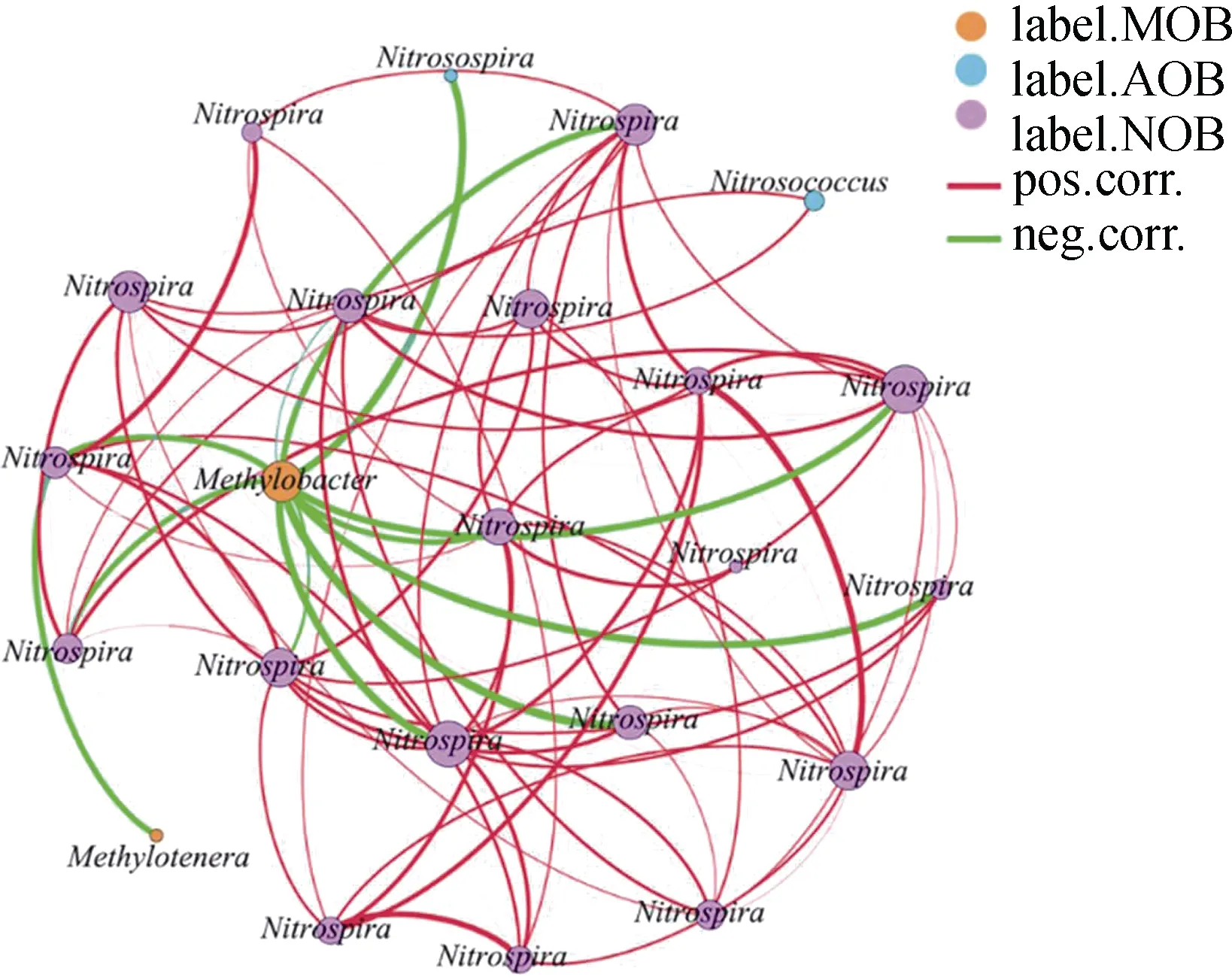
圖6 活性MOB、AOB 和NOB 的共線性網絡圖Fig. 6 Network analysis of co-occurring phylotypes of active MOB,AOB and NOB in microcosms
3 討 論
3.1 氮素水平對甲烷氧化活性和活性甲烷氧化微生物的影響
本世紀初,Stein 等提出銨鹽對MOB 刺激還是抑制取決于銨態氮與甲烷態碳的比例(N-CH)。然而,Bodelier和Zheng等發現N-CH比分別為200 和0.11 時,尿素添加均顯著刺激甲烷氧化。而本實驗中U100(100 μg·g干土)處理的N-CH比僅為0.70,但是顯著抑制了甲烷氧化。同樣,Zhang等通過四年田間試驗也發現100 kg·hm·a(約158 μg·g干土)的氮添加顯著抑制甲烷氧化。Zhao等推測氮肥對甲烷氧化的刺激或抑制作用可能取決于主要甲烷氧化菌的種系型,比如,Zheng 等發現水稻土中添加尿素后,活性甲烷氧化微生物以為主,而未施加尿素的處理以為主。在本草原土的試驗中,無論是否添加尿素以及添加尿素多少,均是以為主。因此,有關氮素水平對甲烷氧化的機理尚不明確,未來關于氮素對甲烷氧化影響的研究可考慮多種類型土壤的綜合分析。
3.2 甲烷對凈硝化活性和活性硝化微生物的影響
類似于對MOB 的分析,本實驗對氨氧化功能微生物在0 d 和21 d 的總DNA 分別進行定量分析。結果顯示(圖3b),低濃度和高濃度氮添加均會刺激AOB 豐度的增長,而抑制AOA 的繁殖,說明AOB 在硝化作用過程中具有重要作用。DNA-SIP 結果發現AOB 在各處理中被C 標記,而AOA 在各種處理中均未表現出明顯的被標記跡象(圖4a),說明AOB 同化了來自CO的C,進一步證明AOB是驅動該草原土壤硝化作用的活性硝化微生物。
不同浮力密度范圍的AOB分布,以及AOB 和NOB 相對豐度的分布進一步表明低氮水平下,甲烷添加顯著抑制了活性硝化微生物同化CO(圖4a,圖4c)。Zheng 等在水稻土的研究發現,100 μg·g干土的尿素處理,甲烷添加顯著抑制硝化活性和活性硝化微生物。而本研究發現,在草原土壤中,添加20 μg·g干土的尿素,甲烷即顯著抑制活性硝化微生物同化CO,降低硝化活性,而100 μg·g干土的尿素處理下,硝化活性和活性硝化微生物對甲烷添加無響應。無機氮對于生物量的形成是至關重要的,尤其是對于那些缺乏固定分子態N 的甲烷氧化菌來說。說明甲烷對硝化活性的抑制作用可能只出現在氨氧化底物不足時,氨氧化微生物和甲烷氧化微生物競爭N 源,一部分N 作為N源被甲烷氧化菌同化,但是當N 源濃度較高時,甲烷氧化微生物反被抑制(圖2,圖4),喪失了與氨氧化微生物競爭的能力。結合上述氮水平對甲烷氧化的影響,氮素添加體現出對甲烷氧化的刺激效應時,甲烷的存在就會抑制氮素轉化(硝化作用);氮素添加體現出對甲烷氧化的抑制效應時,甲烷的存在對氮素轉化過程(硝化作用)沒有顯著影響,說明甲烷氧化和硝化過程的相互作用關系受氮素水平的調控。
3.3 活性甲烷氧化和硝化微生物及相互作用
群落組成分析發現,活性MOB 群落結構比較單一,以Type Ia 的甲基桿菌屬()為主,占90%以上,還有一些Type Ib 的甲基暖菌屬()和極少比例的 Type II 型的。一方面,研究發現Type Ia 型甲烷氧化菌在濕地、水稻土、冰下水系等多種環境中均作為主要的甲烷氧化微生物;另一方面,通過菌株分離分析發現,高濃度甲烷條件下的活性遠高于低濃度甲烷環境。此外,在本試驗中,尿素施入并沒有改變草原土壤中活性甲烷氧化微生物的群落組成,這一結果與Zhao 等在水稻土中的發現一致,說明草原土和水稻土中氮有效性不是決定甲烷氧化微生物組成的關鍵因子。本試驗發現活性硝化微生物以AOB 和NOB 為主,這一結果與之前在草原土壤、水稻土壤和森林土壤中的研究結果一致。
尿素對甲烷氧化菌群落的低促高抑主要反應在MOB,菌株生長需要氮源,但其不具備固氮能力,因此,氮施入可促進生長繁殖。但隨著施氮量增加,硝化活性增強,硝化作用產生的NHOH 和NO以及硝化產物NO對 MOB 均存在毒害作用;活性AOB 和NOB 比例隨施氮量增加而增加,導致MOB 與AOB/NOB 在同一生境中競爭O、NH-N 和生存空間,網絡分析結果也發現與活性硝化微生物(AOB 和NOB)存在顯著負相關關系。結合低氮條件下,甲烷氧化增強而硝化被抑制;高氮水平下,硝化增強而甲烷氧化被抑制,說明草原土壤中甲烷氧化和硝化之間存在氮引發的MOB 與AOB/NOB 之間的競爭性相互作用關系。
4 結 論
草原土壤甲烷氧化活性及微生物功能活性在低濃度氮素水平下顯著增加,而在高濃度氮素水平下顯著降低,呈現出“低促高抑”現象。N 對甲烷氧化的影響(刺激或抑制)可能是土壤條件和主要微生物種系型共同作用的結果。DNA-SIP 結果發現,活性甲烷氧化微生物以為主,活性硝化微生物以AOB 和NOB 為主。而網絡分析結果發現MOB 和AOB/NOB 之間存在顯著負相關關系,說明甲烷氧化和硝化微生物之間存在競爭性相互作用。以上結果表明甲烷氧化和硝化過程的相互作用受氮素水平的調控。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
