高直鏈淀粉禾谷類作物種質創新與利用研究進展
2022-06-09 08:47:50徐妙云邢利娟楊明雨張凌萱王磊劉悅萍
生物技術通報 2022年4期
關鍵詞:水稻
徐妙云 邢利娟 楊明雨 張凌萱 王磊 劉悅萍
(1.中國農業科學院生物技術研究所 農業部農業基因組學重點實驗室(北京),北京100081;2.北京農學院生物與資源環境學院,北京102206)
隨著居民生活水平的提高,因營養失衡引發的慢性病高發問題日益嚴重,《2018全球營養報告》[1]指出,全球有1/5的死亡與飲食有關。該報告首次強調了飲食在消除營養不良方面的重要性。全球194個國家和地區每年因營養性疾病損失3.5萬億美元,而超重和肥胖花費5 000億美元。中國青少年雖然營養不良發生率大幅下降,但超重和肥胖的比例大幅增加。同時中國成人糖尿病、超重和肥胖比例也大幅升高。在接下來的幾年里,需要引導人們培養健康的生活習慣,以期在2025年達到超重、肥胖、“三高”的零增長率目標[2]。因此從食物源頭供給上進行改變,依據營養需求安排農產品供給,開發具有營養和保健功能的食品尤為重要。
淀粉(starch)作為世界上絕大多數人口的主食,在人類生活中起著舉足重輕的作用。根據多糖鏈的形式,淀粉分為直鏈淀粉(amylose)和支鏈淀粉(amylopectin)。根據其在人體中的消化吸收方式不同,淀粉可分為可消化淀粉(digestible starch)、慢消化淀粉(slowly digestible starch)和抗消化淀粉(resistant starch,RS)。抗消化淀粉又稱為抗性淀粉,是指在健康人體的小腸中不能被分解吸收,而在大腸中可被酵母菌發酵分解的淀粉。RS有良好的生理功能,可應用于高膳食纖維、低血糖指數功能性食品的加工和開發。RS含量與直鏈淀粉所占的比例顯著相關[3-4]。因為直鏈淀粉具有很好的凝膠強度,回升值較高,可以形成塑性較好的薄膜,作為重要的工業原料,用途廣泛,涉及到各個領域,如醫療、造紙、包裝、石油、環保、光纖、服裝等行業。同時,直鏈淀粉持水力低、吸水性差、可膨化、糊化溫度高、熱量低等特點,適合用于食品直接加工或作為食品添加劑[5]。就玉米而言,全世界淀粉年產量為3 600萬t,其中80%以上是玉米淀粉,但是普通玉米的直鏈淀粉含量20%-25%,從普通玉米提取直鏈淀粉成本很高;對水稻和小麥而言,通過提高淀粉中直鏈淀粉的含量來提高RS的比例,進而可以開發適合慢性病人的食物,因此加強高直鏈淀粉玉米和水稻、小麥育種具有重要的社會效益和經濟效益。本文將對參與玉米和水稻等禾谷類作物胚乳中淀粉合成代謝的酶類、淀粉的生物合成途徑、直鏈淀粉提高的途徑,以及目前已經選育成功的高RS水稻、小麥和高直鏈淀粉玉米的推廣情況等進行綜述,并展望該領域今后可能的發展。
1 淀粉概述
淀粉以半結晶顆粒的形式存在于谷物或者豆類種子胚乳或植物的各種器官中。目前已知其常見的儲藏器官主要是種子(水稻、小麥、玉米、高梁、大麥、燕麥等),塊莖(馬鈴薯、菊苣、山藥等),塊根(甘薯、木薯等),未成熟的果實(香蕉和芒果),以及葉片(擬南芥、藻類等)。淀粉顆粒形狀有圓形、橢圓形、扁豆形和多角形等,大小范圍在1-100 μm之間。淀粉是由數百上千的單糖分子連接聚合而成的多聚體,是一種多糖。直鏈淀粉是D-葡萄糖基以α-(1,4)糖苷鍵連接的多糖鏈,分子中有200個左右葡萄糖基。雖然直鏈淀粉是線性的,但是其依然存在部分分支,大分子量的直鏈淀粉分子可能具有多達10個或更多分支,不過分支一般相對較短。支鏈淀粉分子中除有α-(1,4)糖苷鍵的糖鏈外,還有α-(1,6)糖苷鍵連接的分支,分子中含300-400個葡萄糖基。淀粉由15%-25%的線性直鏈淀粉、高度分枝和組織化的支鏈淀粉組成[6]。
淀粉合成主要在植物葉片的葉綠體和儲藏組織的淀粉體中進行。在葉綠體中,白天通過光合作用,將CO2固定為淀粉,作為臨時性儲藏物,夜間將淀粉分解,以蔗糖的形式運輸到其它各個組織中。在儲藏器官中蔗糖重新轉化為淀粉,淀粉粒作為長期儲藏形式積累在淀粉體中,大部分在種子萌發和幼苗生長時期被分解,提供萌發和生長所需的能量。
2 參與禾谷類作物胚乳中淀粉合成代謝的主要酶類
葉綠體和淀粉體均是淀粉合成的主要場所,前者通過光合作用合成淀粉,淀粉合成初期以臨時性淀粉形式存儲在葉片中,隨后淀粉降解產生的蔗糖經韌皮部運輸至胚乳。在胞液中經蔗糖合成酶(sucrose synthase)催化分解為果糖和UDP-葡萄糖(UDPG),UDPG轉化為6-磷酸葡萄糖(Glu-6-P)或1-磷酸葡萄糖(Glu-1-P)。Glu-6-P或Glu-1-P轉運進入淀粉體中,經ADP葡萄糖焦磷酸酶(ADP glucose pyrophosphorylase,AGPase)作用生成合成淀粉的底物腺苷二磷酸葡萄糖(adenosine diphosphate glucose,ADPG)。
淀粉的合成是一個復雜的生化調控過程,需要多種酶共同參與,主要有五大類酶,分別是AGPase、淀粉合成酶(starch synthase,SS)(包括顆粒束縛淀粉合成酶(granule-bound starch synthase,GBSS)和可溶性淀粉合成酶(soluble starch synthase,SSS))、分支酶(starch branching enzyme,SBE)和脫分支酶(debranching enzyme,DBE)。各種酶既發揮獨特的作用,部分酶彼此之間又相互作用,并存在功能冗余現象。雖然禾谷類作物胚乳中淀粉合成途徑都主要是這五大類酶分工合作,但在進化過程中,不同作物中每種酶又不盡相同,功能也各異,這里以玉米和水稻胚乳中迄今發現的酶類為例進行闡述,并根據目前的研究,列出了水稻和玉米淀粉合成的異同(圖1)。
2.1 ADP葡萄糖焦磷酸酶AGPase
AGPase催化淀粉合成途徑中第一個關鍵的調控步驟,植物AGPase是同源四聚體,由兩個大亞基(large subunit,LSU)和兩個小亞基(small subunit,SSU)組成[7]。此處,水稻中發現有Pho1以溫度依賴型的方式參與淀粉的合成,pho1水稻突變體中聚合度(degree of polymerization,DP)為11的短鏈比例高,而DP13-21中等長度的鏈比例低,另外,在Glu-1-P存在的情況下,Pho1可以將DP4-6的麥芽低聚糖合成長鏈[8],所以,在水稻的淀粉合成途徑中,Pho1在長鏈糖原的合成中發揮重要作用,但是Pho1基因僅在籽粒發育早期顯著高表達。而玉米和大麥中還沒有發現pho1突變體(圖1)。
2.2 淀粉合成酶SS
淀粉合成酶的功能是將葡萄糖殘基加到引物的非還原端延長葡聚糖的線性糖鏈。到現在為止,玉米胚乳中鑒定了多個淀粉合成酶異構體,有GBSS,SSI,SSII,SSIII 和 SSIV[9]( 圖1)。 水 稻有2個顆粒結合型淀粉合成酶異構體(GBSSI和GBSSII),和10個可溶性的淀粉合成酶異構體,1個SSI,3個SSII異構體(SSIIa/SSII-3,SSIIb/SSII-2和SSIIc/SSII-1),2個SSIII 異構體(SSIIIa/SSIII-2和SSIIIb/SSIII-1),以及 2個 SSIV 異構體(SSIVa/SSIV-1和 SSIVb/SSIV-2)[10-12](圖1)。GBSS 與淀粉粒結合緊密,主要負責直鏈淀粉的合成。水稻、玉米和小麥中,Waxy(wx)基因編碼GBSSI,wx突變體胚乳總淀粉含量變化均不明顯,但AC含量顯著減少或者完全缺失[13-15]。研究表明GBSS除參與直鏈淀粉的合成外,也參與到支鏈淀粉中超長鏈(extra long chains,ELC)的合成[16]。SS或是溶于細胞質基質,或是呈半溶解半偶聯于淀粉粒的狀態,參與支鏈淀粉的合成。每一個亞型的 SS 在支鏈淀粉的合成中有著各自獨特的功能。一般認為SSI可參與合成較短的葡糖鏈(DP8-12),沒有異構體,SSI缺失突變體的種子和淀粉顆粒大小、形態均沒有明顯變化,說明有其他SS可以互補SSI的功能[17];SSII負責中等長度的支鏈合成(DP13-25),玉米sugary(su2)突變體缺失SSII,導致中等長度的鏈減少而短鏈顯著增加[18],但是玉米和水稻SSIIa突變體的淀粉含量和淀粉粒形態沒有顯著變化[18-19];SSIII可能參與長鏈的合成,并且與SSII存在一定相互作用,也有可能補償部分SSIV的作用,水稻SSIIIa缺失突變體淀粉粒排列稀疏,胚乳粉質化,白心[20-21],而玉米突變體也由于淀粉顆粒形態異常而呈現胚乳玻璃狀無光澤的表型[22],說明支鏈淀粉短鏈的變化對淀粉顆粒形態沒有顯著影響;SSIV 可能參與淀粉粒的起始合成,控制淀粉粒的數目,水稻中有兩個SSIV基因SSIVa和SSIVb在灌漿階段持續表達,說明SSIV也參與了淀粉的合成過程,但目前還沒有禾谷類SSIV突變體被鑒定。
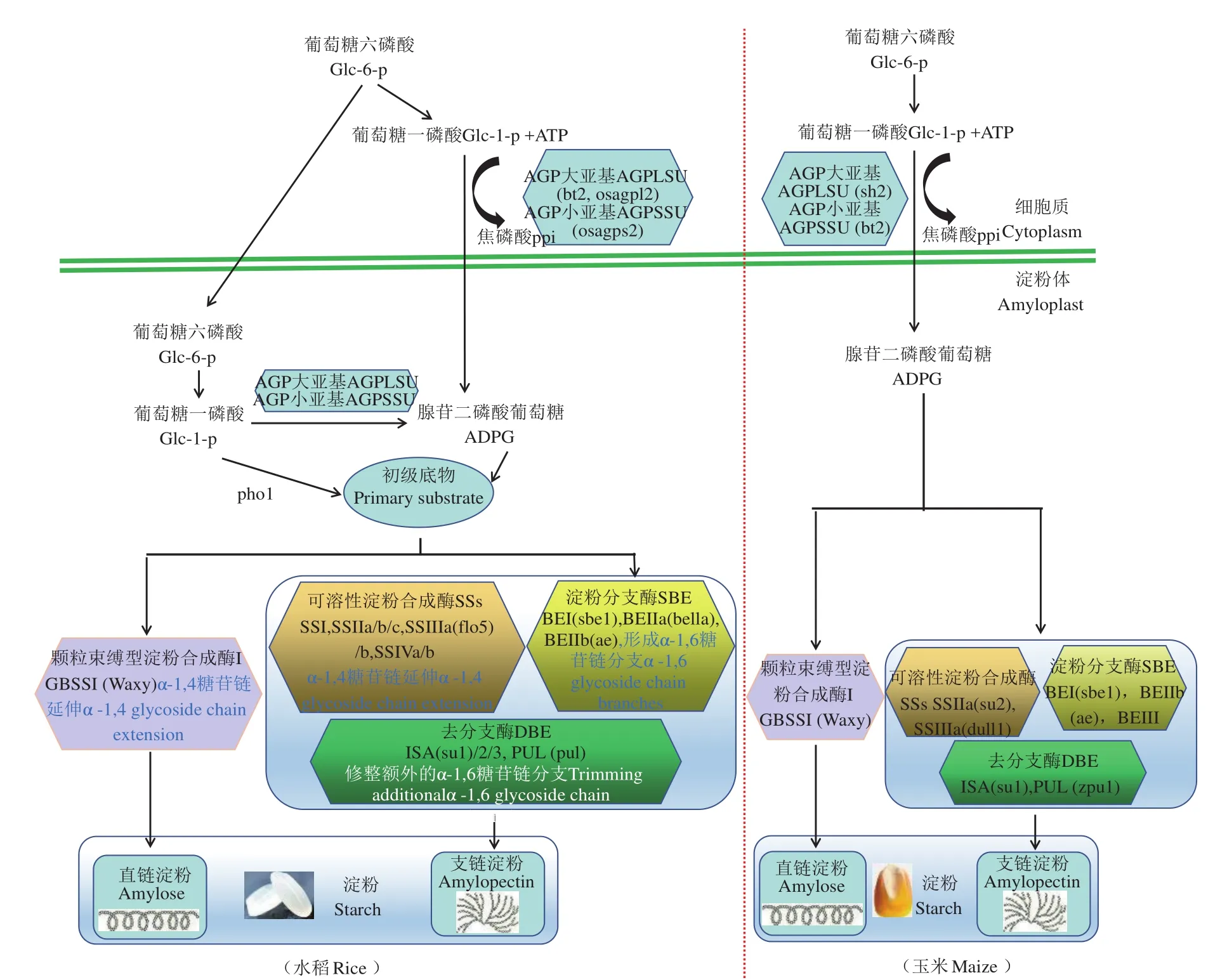
圖1 水稻和玉米中直鏈和支鏈淀粉的合成途徑Fig.1 Synthesis pathways of amylose and amylopectin in rice and maize
2.3 分支酶SBE
SBE將現有的α-1,4 糖苷鍵切斷,然后將含6個或更多的葡萄糖殘基的糖鏈轉移至另外的或同一葡萄糖鏈上的 C6 位置上,可以引入直鏈與支鏈的分支點。目前在水稻和玉米中各發現了3個SBE基因(圖1),SBE基因分為兩個亞型:SBEI和SBEII,這兩類基因編碼的分支酶在支鏈淀粉合成中有著獨特的作用,SBEI傾向于產生DP大于16的長鏈,而SBEII則傾向于產生DP小于12的短鏈。雙子葉植物只有一種SBEII,而單子葉則有兩種異構體SBEIIa和SBEIIb。SBEIIa在植物所有組織中都表達,而SBEIIb只在胚乳中特異表達。玉米和水稻SBEIIa突變體中支鏈淀粉的鏈分布模式沒有明顯變化,而玉米SBEIIb突變體中直鏈淀粉顯著增加[23],說明SBEIIb特異的參與支鏈淀粉中短鏈的形成,而SBEIIa可能有部分冗余功能。最新對sbeIIb水稻功能缺失突變體的研究發現,sbeIIb水稻除了直鏈淀粉含量從19.6%提高到27.4%,抗性淀粉含量從0.2%提高到17.2%外,許多編碼AGPase,可溶性淀粉合成酶和其他淀粉分支酶異構體的基因均上調表達,而編碼顆粒束縛淀粉合成酶、去分支酶、普魯蘭酶和淀粉磷酸化酶下調表達。同時,胚乳中糖、脂肪酸、氨基酸和植物甾醇含量均增加,表明淀粉合成途徑對其他初級和次級代謝物的積累都產生了較大影響[24]。
2.4 去分支酶DBE
去分支酶也稱為α-1,6-葡聚糖水解酶,它的作用是清除錯誤的分支點,使得支鏈淀粉形成易于結晶的螺旋結構,是決定淀粉結構形成的一個關鍵酶。目前在水稻中報道了4個DBE基因,玉米2個DBE基因(圖1)。DBE分為兩類:異淀粉酶(isoamylase,ISA)和普魯蘭酶(pullulanase,PUL)。ISA主要對可溶性的植物糖原和支鏈淀粉進行去分支,而PUL主要作用于普魯蘭糖和支鏈淀粉,但不作用于植物糖原。目前鑒定到植物中至少有3個ISA基因,但只有1個PUL基因[25]。
3 提高禾谷類作物中直鏈淀粉含量(amylose content,AC)的途徑
目前提高禾谷類作物胚乳中AC主要是通過對淀粉合成途徑中3個環節的關鍵酶基因進行改良來實現:一是提高AGPase的活性,從而增加底物ADPG的濃度[26-31];二是提高GBSSI的表達,但因糖鏈的還原性末端有限,并且直鏈淀粉與支鏈淀粉二者之間存在底物競爭,導致AC達到一定水平后就不再上升[25-27];三是降低SBEs,SSIIa,或者參與支鏈淀粉合成的其他酶類[28-30]。
3.1 通過提高AGPase的活性來增加胚乳中直鏈淀粉含量
水稻中,AGPase基因家族包括兩個SSU基因,OsAGPS1和 OsAGPS2,4個 LSU基 因,OsAGPL1,OsAGPL2,OsAGPL3 和 OsAGPL4[26-28]。玉米中迄今為止,鑒定了分別編碼胚乳特異性的LSU和SSU亞基 shrunken2 和 brittle2 基因[29-31]。過表達熱穩定的玉米LSU(sh2r6hs)能夠使水稻胚乳的淀粉含量提高23%[28]。同樣,轉玉米sh2r6hs基因的小麥種子中AGPase酶活性提高,每株種子重量平均提高38%,生物量提高31%[32],同時促進光合作用和碳代謝[33]。在小麥胚乳中特異性表達TaLSU1基因可以顯著提高種子中AGPase酶活性、胚乳中淀粉重量、穗粒數和單粒重[34]。
3.2 通過降低參與支鏈淀粉合成酶的活性來提高直鏈淀粉含量
SSIIa降低對胚乳中直鏈淀粉含量的改變因物種而異,例如SSIIa功能缺失的玉米突變體sugary2(su2)中直鏈淀粉含量可以從常規的25%-30%提高到40%-50%[18],但是水稻中影響不大,SSIIa功能缺失的粳稻突變體胚乳中直鏈淀粉含量提高4%,與野生秈稻中含量相當[35]。而SSIIa功能缺失的小麥突變體籽粒皺縮,直鏈淀粉和抗性淀粉含量均提高,同時淀粉的可消化性降低[36-37]。分支酶活性降低或者完全喪失而產生的高直鏈淀粉水稻、小麥或者玉米株系,實際是因為支鏈淀粉的結構發生了較大變化,主要以長鏈分支為主[38-40]。支鏈淀粉的長鏈分支比例越高,直鏈淀粉含量越高[41]。
與降低SSIIa活性相比較,失活SBE的活性可以大幅度提高水稻、小麥和玉米中直鏈淀粉的含量。禾谷類作物中主要有3類SBE,分別為SBEI,SBEIIa和SBEIIb。研究表明,降低SBEI活性,對淀粉的成分和籽粒表型沒有明顯影響,只會引起籽粒中支鏈淀粉精細結構的微小變化[42-43]。SBEIIa基因缺失的水稻和玉米胚乳直鏈淀粉含量和支鏈淀粉的精細結構均沒有明顯的變化,但葉片中同化產物受到極大影響[10,44]。這可能是因為SBEIIa基因主要在葉片中特異性表達,因而不適合作為降低籽粒中支鏈淀粉含量的候選基因。而SBEIIb缺失(一般稱為ae)突變體中直鏈淀粉含量能達到61.7%-67.7%,增加幅度35%[45]。水稻無論是粳稻還是秈稻,其ae突變體中直鏈淀粉含量均高于野生型,但是增加幅度最高15%,遠遠低于玉米ae突變體[38]。當SBEIIb與SBEI 在秈稻中組合突變后直鏈淀粉含量提高到了60%,顯著高于SBEIIb單突變[46]。與水稻和玉米相反,抑制SBEIIb基因表達對小麥和大麥籽粒中直鏈淀粉含量影響不大[47],而SBEIIa基因功能缺失的小麥和大麥突變體籽粒中直鏈淀粉含量分別提高到55%和38%[48-49]。
另外一種提高直鏈淀粉含量的有效方法是將SBEII單個突變與其他基因突變組合,例如缺失SSIIIa的水稻中直鏈淀粉含量為30.7%,將該突變引入SBEIIb缺失突變的背景中,雙突變體的直鏈淀粉含量達到45%[50]。最新的研究報道了GBSSI基因野生型與SSIIa和SSIIIa等位基因發生突變的組合能使水稻籽粒中直鏈淀粉的含量提高17%[51]。
4 高直鏈淀粉禾谷類作物種質培育與推廣應用
高直鏈淀粉水稻和小麥的應用價值,除了可作為食品添加劑之外,更重要的是直鏈淀粉含量的增加使水稻和小麥籽粒中抗性淀粉RS的含量提高。RS因為具有“控制餐后血糖及防止糖尿病、降血脂和控制體重、有利于腸道健康、抗消化及降低食物的熱效應、促進鈣鎂鋅離子的吸收”等重要生理功能,備受國內外營養專家和功能食品專家的重視,研制高RS谷物作物來防止人類慢性病的發展,成為了國內外育種和功能食品研究的新興領域。市售水稻品種RS含量基本低于3%。2006年,浙江大學培育了首個高RS早秈稻突變體新品種:浙輻201,RS含量達3.6%[52]。該團隊以主推雜交水稻恢復系R7954為起始材料,創制了突變體RS111,其熱米飯中RS含量可達7%[53]。在此基礎上,浙江綠巨人生物技術有限公司與浙江大學團隊合作,開發了RS為8%以上的功能型水稻,2007年11月正式命名為“宜糖米”。同時,云南省農科院生物所與浙江大學團隊合作開發了RS高達10%的功能型水稻功米3號,于2008年獲得了新品種權,商品名為“適糖米”,并且根據不同人群的需求,開發了“適糖米普通型”和“適糖米加強型”。宜糖米和適糖米均為秈稻,目前在市場上都有銷售。2010年,上海農科院利用花藥培養技術和常規育種技術,在國內首次選育出較普通水稻品種RS含量高10倍以上的粳稻新品系:降糖稻1號,其RS含量超過13%。近年來,該團隊以降糖稻1號為供體,利用分子育種技術,成功選育了功能性和高產優質兼顧的新高RS含量的水稻品系優糖稻2號和優糖稻3號。該系列商品名為“優糖米”,目前也在市場有銷售。
小麥高抗性淀粉種質創新方面,2016年加利福尼亞大學戴維斯分校將SBEIIa基因的六倍體小麥Lassik EMS突變體和SBEIIb基因的Lassik EMS突變體進行雜交,獲得了1個直鏈淀粉和抗性淀粉均大幅度提高的突變組合[54]。2020年,中國農業科學院作物科學研究所利用基因編輯技術,定點敲除冬小麥品種鄭麥7698和春小麥品種Bobwhite中的SBEIIa基因,分別獲得了高抗性淀粉的冬、春小麥新種質[39],為培育營養功能型小麥新品種提供了新途徑。
玉米方面,在20世紀70年代,用于商業生產高直鏈淀粉玉米雜交種有兩種類型:C1assⅥ(含直鏈淀粉 50%)和 C1assⅦ(含直鏈淀粉 70%-80%)。到20世紀80年代時,Custom 農場種子公司推出直鏈淀粉含量超過 94%的 C1assⅧ和 C1assⅨ玉米單交種。我國以前沒有高直鏈淀粉玉米品種,也沒有高直鏈淀粉玉米種植和淀粉生產。經過多年的努力,中國農業大學國家玉米改良中心在國內率先培育出AC 達50%左右的高直鏈淀粉玉米中農大401。
5 展望
高直鏈淀粉水稻的選育工作雖然目前已經取得了一些不錯的進展,也成功推廣了幾個商業化的產品,但是還存在產量低、堊白率高、加工性能和口感差、目標性狀單一等問題,在今后的研究中可以在考慮改善農藝性狀、提高加工性能的同時,向多目標復合性狀方面發展。而我國高抗性淀粉小麥的種質創新工作剛剛起步,今后可以進一步利用基因工程技術和傳統育種技術相結合,加快高抗性淀粉小麥、大麥的品種選育。
全世界淀粉年產量為3 600萬t,其中80%以上是玉米淀粉,因此,加強高淀粉玉米育種具有重要的社會效益和經濟效益。直鏈淀粉是重要的工業原料,用途廣泛,涉及到各個領域,如食品、塑料、醫療等行業。但是普通玉米的直鏈淀粉含量20%-25%,從普通玉米提取直鏈淀粉成本很高。而相對于水稻,玉米的高直鏈淀粉選育工作要遠遠落后。目前,我國所需的直鏈淀粉主要依賴進口,價格昂貴。美國高直鏈淀粉玉米雜交品種的直鏈淀粉含量規定在50%以上。ae 基因的發現是玉米高直鏈淀粉育種的重要里程碑。該基因純合體玉米胚乳中直鏈淀粉的含量從 25%增加到 70%。采用常規育種技術選育高直鏈淀粉玉米遇到的主要問題是玉米淀粉總含量減少,籽粒含水分量增高,最終產量降低。接下來的工作中要更注重于挖掘參與直鏈淀粉含量調控和修飾的基因,并闡明其分子機制。隨著基因工程技術,以及各種淀粉特性分析技術的發展,相信不久的將來,我國高直鏈淀粉玉米的選育工作將會有所突破。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
