水稻鋅吸收、轉運、累積機理研究進展
2022-06-09 08:47:50薛欣月于雪然劉曉剛馬嘉欣田蕾李培富
生物技術通報 2022年4期
薛欣月 于雪然 劉曉剛 馬嘉欣 田蕾 李培富
(寧夏優勢特色作物現代分子育種重點實驗室 寧夏大學農學院,銀川 750021)
鋅是人體必需的微量元素之一,主要分布于人體的肌肉、血液和骨骼中[1],參與人體多種新陳代謝過程、包括DNA聚合酶等多種酶的合成、細胞分解、能量和其他物質的代謝,其不足阻礙人體的生長發育[2],鋅對人體健康具有十分重要的調控作用。目前,我國部分地區的兒童及青少年存在不同程度的鋅營養缺乏癥,但直接服用高鋅含量藥物不利于人體健康[2]。研究表明植物體中的鋅可以更安全、高效地被人體吸收,因此鋅在植物體中的轉運機理受到研究者的高度重視[3]。水稻作為主要糧食作物,提高水稻籽粒鋅含量,將會有效緩解人類鋅缺乏的困境。研究水稻體內鋅元素吸收、轉運和累積機理,可以為富鋅水稻的創制奠定理論基礎。
1 鋅在水稻體內的生理功能
1.1 生物體內多種酶的組分和活化劑
鋅在生物體內主要以二價陽離子形態(Zn2+)存在,且是多種酶的組成成分。研究表明,動物、植物、微生物中至少已鑒定出300多種酶的活性與功能受到鋅元素的調節[4]。其中,參與水稻生理功能的鋅酶(zinc enzymes)主要分為含鋅金屬酶和需鋅酶兩大類(表1)。

表1 鋅酶及其可能功能Table 1 Zinc enzymes and their possible functions
鋅是水稻體內多種含鋅酶的組成成分,在含鋅酶中主要發揮的作用包括催化作用、結構功能、輔酶、共催化功能等。碳酸酐酶存在于水稻葉綠體外膜,參與光合作用[7],當水稻植株內鋅元素缺乏程度加劇時,葉片中碳酸酐酶的活性降低,阻礙了水稻植株的光合作用,水稻產量將會大幅降低[5]。鋅是合成谷氨酸過程中必不可少的元素,是谷氨酸脫氫酶(GDH)的主要成分,谷氨酸脫氨酶普遍存在于水稻線粒體內,在水稻碳氮代謝中具有重要作用[8]。鋅是醛縮酶的激活劑,醛縮酶主要存在于細胞質內,參與水稻光合作用碳代謝過程。銅鋅超氧化物歧化酶(Cu/Zn-SOD)是1種重要的含鋅金屬抗氧化酶,主要存在于水稻細胞質和葉綠體中[9],其中含有Cu2+和Zn2+兩個金屬輔基,銅鋅超氧化物歧化酶對水稻抗氧化、抗逆、抗衰老具有重要意義,Sen等[13]研究表明在水稻中具有充足的Cu/Zn-SOD時,能顯著提高水稻的抗鹽、抗旱及抗寒性等。
鋅是水稻體內過氧化氫酶(catalase)、核酮糖-1,5-二磷酸羧化酶(rubisco)、過氧化物酶(peroxidase)等多種需鋅酶的活化因子。過氧化氫酶分布于水稻葉綠體中,參與光呼吸、共生性氮固定,當鋅營養缺乏時,CAT活性降低,容易導致生物膜的過氧化損傷,引起有機物和無機物從根細胞滲漏[7]。核酮糖-1,5-二磷酸羧化酶是光合作用碳反應中重要的羧化酶,當水稻中鋅營養適當時,有利于光合色素的形成,Rubisco活性增強,保證光合作用正常進行,促進水稻種子萌發、根系發育[10]。過氧化物酶可促進水稻光合作用、呼吸作用,研究表明,不同濃度Zn2+對水稻體內的POD活性影響不同[12]。
1.2 參與生長素的合成
鋅元素能促進水稻體內吲哚和絲氨酸合成色氨酸,而色氨酸是生長素合成的必要條件,因此鋅間接影響生長素的形成。鋅缺乏會導致水稻中生長素含量降低,從而使水稻植株矮小[14]。水稻生長旺盛的部位都含大量的鋅和生長素[15-17],缺鋅會使植株中生長素含量降低,影響植株生長發育速度,蛋白質合成受阻,生物量明顯下降,結實率下降、空秕率上升,從而影響產量和品質[18]。
1.3 參與光合作用
Rubisco是水稻體內重要的需鋅酶,存在于水稻葉綠體中,是水稻光呼吸以及光合作用中決定碳同化速率的關鍵酶[21]。植株體內鋅含量過低時Rubisco活性降低,導致ATP中的化學能轉化為葡萄糖的過程受到抑制[10]。1,6-二磷酸果糖酶主要存在于葉綠體中,參與光合產物的轉運,1,6-二磷酸果糖酶是葉綠體中裂解六碳糖的關鍵酶,在植物缺鋅時該酶活性顯著降低,導致光合產物向細胞質中的轉運受到抑制[7]。
1.4 鋅與水稻病害
鋅的抗病性可能與鋅在維持膜的結構和功能方面的重要作用有關,對根系生物膜的保護作用尤為突出[22-24]。當水稻缺鋅時,植株對病原體的抗性減弱,體內的天門冬酰胺和谷氨酰胺含量增加,加速了病原體的侵染和生長,導致水稻病害加重。此外,缺鋅水稻根際碳水化合物和游離氨基酸滲出量增多,致使病原菌快速繁殖[25]。
水稻缺鋅引發水稻縮苗病,往往在土壤有效鋅含量低或氨鋅比例失調時發病。輕度缺鋅時,葉片出現褐色斑點,葉片失綠變白;缺鋅嚴重時,植株生長受到抑制,新葉變小、植株矮小,致使水稻不能抽穗,明顯矮縮[26-27]。
2 水稻鋅的生物強化
水稻是世界上最重要的農作物之一,在全球大部分地區都有種植,是人類重要的主食之一。據估計,人類消耗的熱量中有19%來自大米[28]。然而,稻米中的鋅等礦物質含量低,因此鋅是人類飲食中最常缺乏的營養元素[29]。在谷物中,大米的礦物質含量和遺傳變異性都是最低的[30],考慮到大米的廣泛消費和低營養品質,水稻的生物強化被認為是向人類提供鋅含量增加谷物的最佳解決方案之一。生物強化是指使植物在其可食部分(如籽粒、葉子和根)積累更高濃度的可利用營養元素[30-31]。人們可通過增加稻米中鋅的絕對含量來改善人體鋅缺乏[32]。但稻米中存在植酸[33]、纖維素等因素,導致膳食中鋅的生物有效性低[34]。研究表明在水稻中,植酸與二價離子(如 Zn2+、Ca2+、Mg2+、Fe2+)形成絡合物,礦物的生物有效性顯著降低[35-36],所以要通過提高水稻籽粒有效鋅以及稻米中鋅的絕對含量來進行生物強化。
3 鋅與水稻產量及品質的關系
鋅在水稻的生長發育中具有不可替代性。鋅是水稻多種酶的構成元素,也參與光合作用、激素合成等多種重要的生理生化反應,在水稻的不同生育時期發揮著重要作用,從而影響水稻的產量和品質。
根據現有工藝流程,將硝化工藝的循環泵出口管線分成兩路,第一路繼續將好氧池底部的活性污泥循環至好氧池頂部,不影響硝化工藝的正常運行;第二路將好氧池底部的活性污泥循環至厭氧池,使其進行反硝化反應。并可根據需要調節第一、二兩路的閥門開度,控制進入好氧池、厭氧池的循環污泥比例,使中水系統處于最佳的工作狀態。
3.1 鋅與水稻產量
祁明等[37]研究發現,在石灰性土壤中水稻易發生缺鋅現象,水稻分蘗期缺鋅非蛋白氮含量顯著增加,蛋白氮總量減少。水稻內雙氨基和多氨基的氨基酸含量明顯降低,導致蛋白質的合成受阻。王孟蘭[38]研究表明,在缺鋅的土壤上施用鋅肥,可有效促進氮素吸收,通過增加單位面積穗數和穗粒數增加水稻產量,但施鋅量過高會抑制水稻對鋅的吸收,并導致減產,水稻產量隨施鋅量的增加而先遞增后減少。麥日桂[39]研究表明,在水稻生長期間,增施鋅肥能有效改善水稻的農藝性狀,以達到增產提質目標。
3.2 鋅與水稻品質
施用鋅肥可改善稻米品質,如提升稻米的食味值和香味,鋅可增強水稻植株的光合作用,促進糖類代謝,改善稻米口感[40]。張麗等[41]研究發現,基施鋅肥或基施+噴施鋅肥可有效增加稻米的蛋白質含量,合理噴施鋅肥可有效增加水稻籽粒中鋅含量的積累,改善水稻籽粒營養品質,如降低稻米碳水化合物含量等。袁少文等[42]研究表明,稻米中鋅含量與噴施鋅肥濃度呈正相關,噴施濃度為2.5%時,稻米中鋅含量最高。
4 水稻不同營養器官鋅吸收途徑
水稻植株對鋅元素的吸收主要通過兩種方式進行。一方面通過水稻根系通過土壤中有效鋅的吸收及鋅肥施用進行吸收,另一方面通過鋅的葉面噴施鋅肥的方式進行鋅吸收。
4.1 水稻根系鋅吸收途徑
水稻根系鋅吸收途徑主要有兩種,分別是質外體和共質體途徑,共質體途徑為主要方式[43]。質外體途徑為自由擴散,無能量消耗,共質體途徑為主動運輸,消耗能量。Zn2+通過被動和主動運輸由細胞膜進入根系細胞,在根中Zn2+從木質部薄壁細胞裝載到木質部[44]。
4.2 水稻葉片鋅吸收途徑
水稻葉片鋅吸收途徑主要是葉面噴施鋅肥。葉片對鋅的吸收主要通過氣孔或表皮細胞進入葉片表皮。但是由于葉片表面張力作用,純水很難自由進入氣孔,所以要借助外部力量,在葉面鋅肥中加入表面活性劑可以降低葉片表面的張力,增加水稻葉片對鋅肥的吸收量[45-46]。但有些表面活性劑會對水稻植株產生毒害作用,影響植株光合作用,所以要注意表面活性劑的用量。Zn2+最后大部分積累在葉、莖和籽粒中,而不是根部[47]。
5 水稻植株鋅吸收、轉運的影響因素
水稻植株在進行鋅的吸收轉運過程中會受到內在因素和外在因素的影響,制約水稻植株對鋅的吸收轉運。內在因素包括水稻品種多樣性、水稻生育時期的影響和水稻籽粒鋅含量相關QTLs的影響。外在因素包括土壤理化性質和農藝措施的影響[32]。
5.1 內在因素
5.1.1 水稻品種多樣性 不同水稻品種對鋅的吸收積累量存在顯著差異。Yang等[48]對285份水稻品種的籽粒礦質元素含量進行測定,發現籽粒中鋅含量秈稻>粳稻,紅米>白米。研究表明云南稻核心種質的鋅含量,地方品種明顯高于改良品種,地方稻核心種質的鋅含量在秈粳、光殼與否及有無芒之間差異不顯著,但水陸、軟米與普通米、紫米與紅米間的鋅含量差異極顯著,以陸稻鋅含量最高,軟米則最低[49]。Gregorio等[50]對 1 138 份不同來源的水稻品種籽粒鋅含量進行測定發現,鋅元素在不同種質間的變異范圍為15.3-58.4 mg/kg。
5.1.2 水稻生育時期 張標金[51]研究表明,不同水稻品種在分蘗盛期對鋅的吸收能力均為最強,因為營養器官發育最關鍵的時期為水稻分蘗期,植株需大量的營養供生殖生長所需,此時根系活力增強,大量的礦質元素儲存在根部。張濟龍等[52]發現植株灌漿期鋅含量低于成熟期,尤其是根和莖中的鋅含量,這可能是由于鋅在水稻根系中的積累濃度已達到峰值,而鋅在水稻根中的積累主要在水稻灌漿期進行。但當鋅濃度低于0.1 mg/L時,植株在灌漿期對鋅的吸收量與成熟期相近。
5.1.3 水稻籽粒鋅含量相關QTLs 鋅在水稻籽粒中的積累受多基因控制,不同水稻品種籽粒鋅含量的差異也受不同基因控制,國內外學者在水稻每條染色體上都發現了控制水稻籽粒鋅含量的QTL(表2)。

表2 水稻籽粒鋅含量相關QTL簡表Table 2 A brief list of QTLs related to zinc content in rice grains
雖然近年在有關水稻籽粒鋅含量的QTL定位的研究取得了一定進展,但由于水稻控制鋅含量的遺傳機制較為復雜,所以對相關基因的精細定位和克隆研究一直進展較慢。這些研究不僅為眾多學者在水稻籽粒鋅含量相關基因研究方面提供了經驗,也為進一步闡明控制水稻籽粒鋅含量基因的功能分析和相關目標基因的克隆奠定了基礎,使通過分子標記輔助選擇指導富鋅水稻品種成為了可能。
5.2 外在因素
5.2.1 土壤理化性質 土壤中的有效鋅可以反映土壤的供鋅能力[68],有效鋅受成土母質、土壤pH值等眾多因素的影響。水稻植株對鋅的吸收量因土壤質地的不同而變化,水稻土和紅壤等類型的土壤有效鋅含量均較高,草甸風沙土、石灰性紫色土和灌耕土等的有效鋅含量較低[69]。植株根系對鋅的吸收速率在砂土中明顯高于壤土及黏土。除此以外,水稻中的鋅元素來源于土壤有效鋅和土壤速效鋅,水稻植株可以直接吸收和利用土壤中的鋅元素。速效鋅可以衡量土壤的供鋅能力,當其含量偏低時,水稻缺鋅癥狀加重[70]。
5.2.2 農藝措施 施用鋅肥被認為是一個水稻增產優質的有效措施。增施鋅肥可以促進水稻生殖生長,增加粒重及產量。研究表明葉面噴灑鋅肥能提高果實及種子中鋅的積累量[17]。王晨等[71]研究表明施用硫酸鋅可有效提高水稻籽粒鋅含量,施鋅能提高水稻不同營養器官的鋅濃度,也可以通過施用氨基酸鋅復合物來增加鋅在籽粒中積累。
吳春勇等[23]認為水稻盛花期噴灑CTK可促進鋅在劍葉中的積累。研究表明增加施氮量可促進水稻對鋅的吸收與累積,氮肥與鋅肥同時施用對水稻生長具有協助作用[72],增施鋅肥能有效增加水稻籽粒的氮濃度和粗蛋白含量,其含量增加與施鋅量呈正相關[73]。
6 鋅轉運蛋白
鋅是植物中重要蛋白結構的輔助因子和活化劑。水稻中有大量的轉運蛋白參與鋅的吸收、轉運和再轉運,對水稻產量和營養品質都有重要影響[74]。水稻體內Zn2+的轉運與鋅轉運蛋白家族密不可分。研究學者在多種高等植物中發現了鋅轉運蛋白家族,如YSL蛋白家族、ZIP蛋白家族、NRAMP蛋白家族(自然抗性巨噬細胞蛋白家族)、CDF蛋白家族、P1b-ATP酶、Mg2+/H+的反向交換轉運蛋白家族、三磷酸結合盒轉運蛋白等[74-76]。鋅轉運蛋白包括鋅吸收蛋白以及鋅排出蛋白兩大類[77]。
6.1 鋅吸收蛋白種類
鋅吸收蛋白包括YSL蛋白家族、ZIP蛋白家族、NRAMP蛋白家族。
6.1.1 YSL蛋白家族 目前在擬南芥、玉米、小麥、水稻、大麥中都有發現YSL蛋白家族(yellow stripelike protein,YSL)的存在。YSL蛋白家族主要參與Fe2+、Mn2+、Cu2+等重金屬離子的轉運。但還未在水稻基因組中發現有關Zn2+吸收轉運的的蛋白基因[78]。
6.1.2 ZIP蛋白家族 金屬穩態對植物的生長、發育和對環境脅迫的適應至關重要,主要由各種金屬轉運體控制。植物ZIP(Zn-regulated transporter,Iron-regulated transporter-like Protein)家族蛋白屬于負責吸收和分配必需和非必需金屬的完整膜轉運蛋白。參與鋅吸收的蛋白為鋅調控轉運體(Zinc regulated transporter,ZRT),研究學者在擬南芥、水稻、苜蓿等植物中鑒定出多個ZIP基因[79-80]。已知水稻ZIP蛋白基因家族主要有9個鋅調控轉運蛋白基因,分別為OsZIP1、OsZIP2、OsZIP3、OsZIP4、OsZIP5、OsZIP6、OsZIP7、OsZIP8、OsIRT1,其表達部位、表達誘導條件及功能如表3所示。在水稻中,OsZIP4、OsZIP5、OsZIP7和OsZIP8在地上部和根中對鋅的分配中起著重要作用,這些轉運蛋白可用于提高鋅的吸收或分配,改善水稻生長。
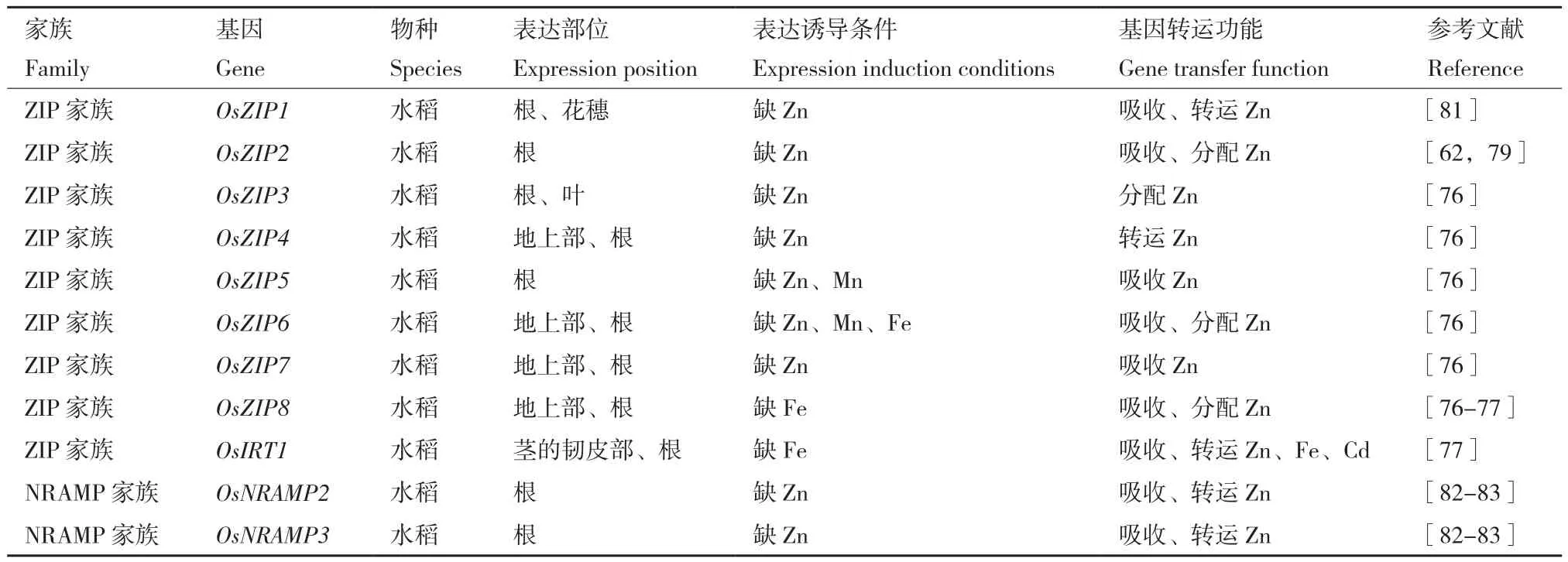
表3 鋅吸收蛋白種類、功能及表達特征Table 3 Types, functions and expression characteristics of zinc absorption proteins
6.1.3 NRAMP蛋白家族 NRAMP蛋白家族為膜蛋白家族,廣泛存在于各種生物體內,主要負責部分二價金屬陽離子的吸收。孫麗娟等[82]研究表明,水稻中OsNRAMP2和OsNRAMP3參與鋅的轉運,其表達部位均位于水稻根部。圖1為現階段已發現水稻中11個鋅轉運蛋白成員的氨基酸序列比對,其親緣關系如圖2所示。
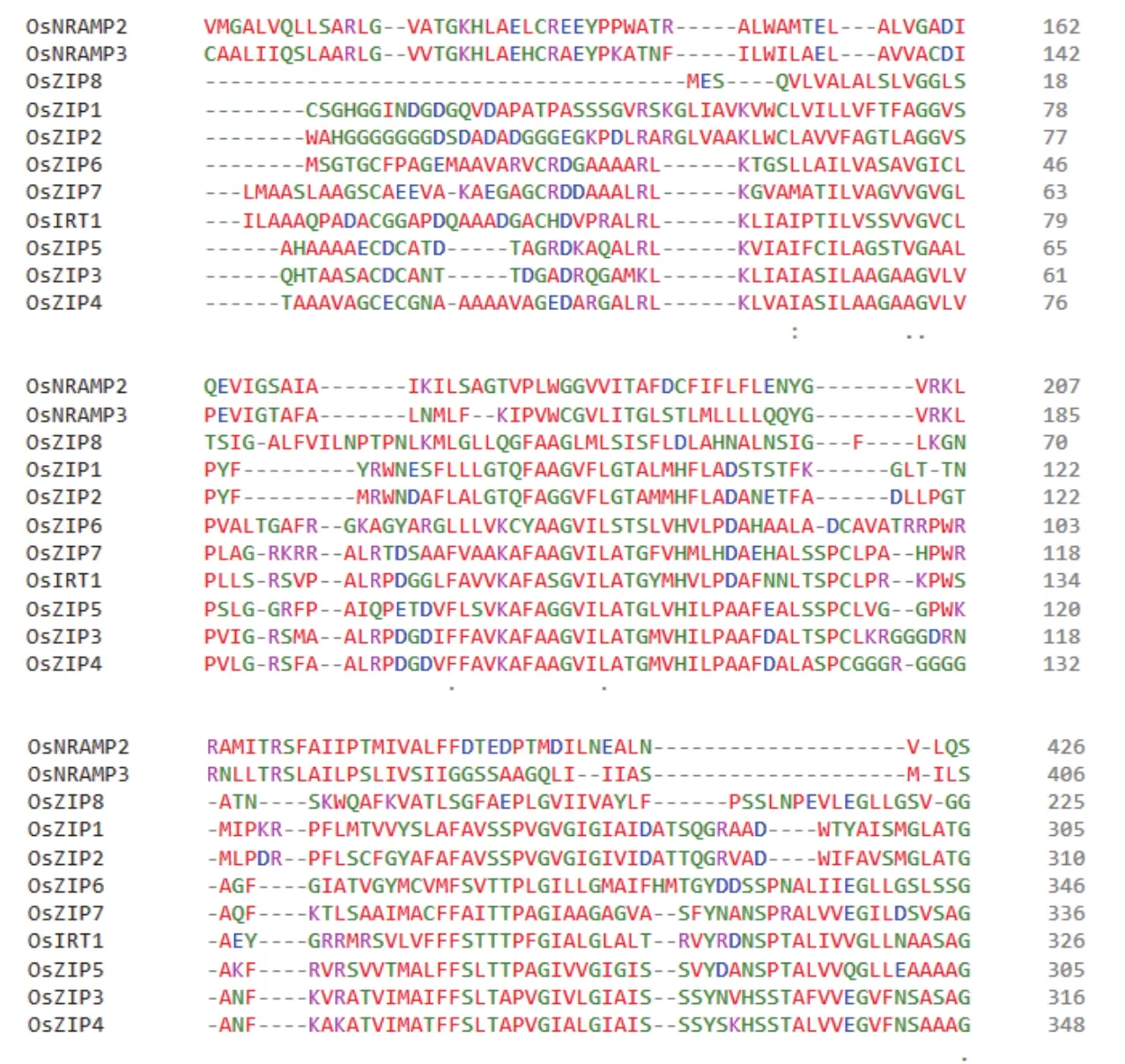
圖1 鋅吸收蛋白氨基酸序列比對Fig.1 Amino acid sequence alignment of zinc uptake proteins
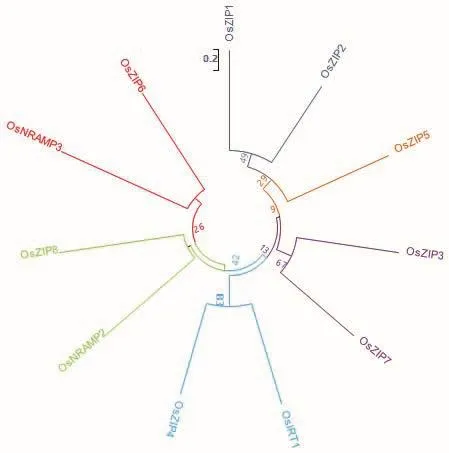
圖2 鋅吸收蛋白進化樹Fig.2 Zinc-absorbing protein evolutionary tree
6.2 鋅排出蛋白種類
當前發現的鋅排出蛋白包括CDF蛋白家族、P1B型ATPases、Mg2+/H+的反向交換轉運蛋白家族、三磷酸結合盒轉運蛋白[76]。但目前還未在Mg2+/H+的反向交換轉運蛋白家族中發現有關水稻Zn2+吸收轉運的的蛋白基因。
6.2.1 CDF蛋白家族 CDF家族蛋白主要參與鋅、鎘和鎳離子的轉運,其作用是將過量的鋅離子排出細胞,避免過量的鋅對細胞產生毒害[83]。目前已發現有關水稻Zn2+吸收轉運的的蛋白基因有OsMTP1和OZT1。OZT1在Zn、Cd脅迫下于水稻各部位處表達,參與鋅和鎘的轉運[78]。
6.2.2 P1B型ATPases 研究學者在水稻基因組中發現9個基因(OsHMA1-9)[84],其中有3個基因為鋅調控轉運蛋白基因,為OsHMA1、OsHMA2、OsHMA3,分別作用于水稻地上部、根、葉,基因轉運功能如表4所示。OsHMA2在營養生長期主要在根系成熟區表達,在生殖期主要在節間達,它的表達不因鋅的有效性而改變。OsHMA2有助于鋅和鎘的轉運,特別是對發育中的組織[85]。
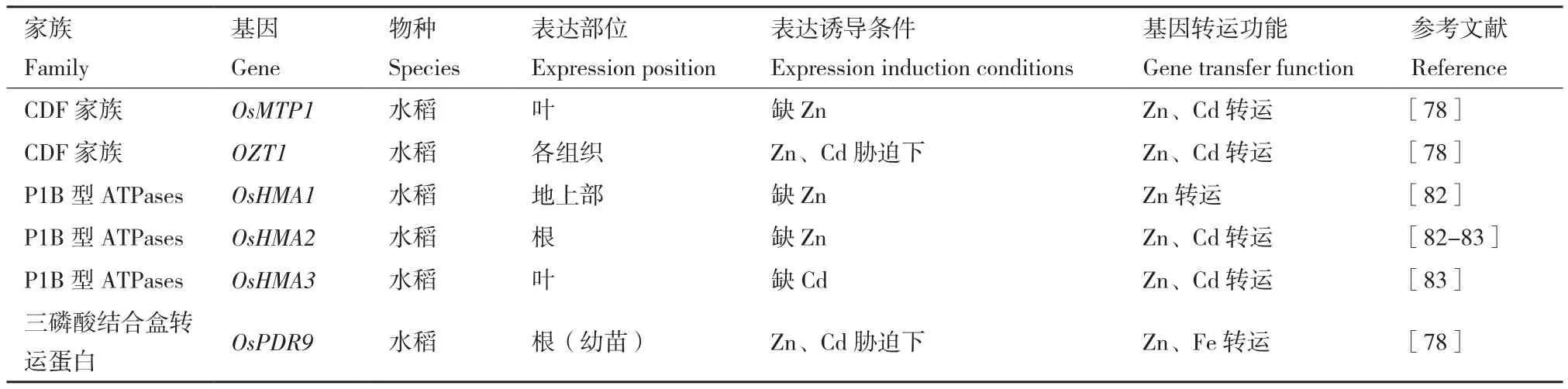
表4 鋅排出蛋白種類、功能及表達特征Table 4 Types, functions and expression characteristics of zinc excretion proteins
6.2.3 三磷酸結合盒轉運蛋白 OsPDR9是在水稻中發現的1種參與鋅鐵轉運的三磷酸結合盒蛋白,在Zn和Cd誘導下,OsPDR9在水稻幼苗的根中表達[79]。圖3為現階段已發現水稻中6個鋅排出蛋白成員的氨基酸序列比對,其親緣關系如圖4所示。

圖3 鋅排出蛋白氨基酸序列比對Fig.3 Zinc excretion protein amino acid sequence alignment
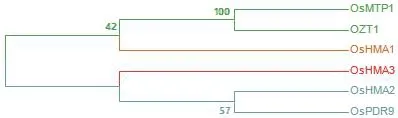
圖4 鋅排出蛋白進化樹Fig.4 Zinc excretion protein evolutionary tree
6.3 鋅轉運相關蛋白基因克隆
現已有大量研究表明鋅轉運相關蛋白可參與鋅的吸收轉運及分配,深入研究其分子機制及其基因克隆,可為人們培育富鋅水稻奠定理論基礎。因此,研究學者對鋅轉運相關蛋白的基因克隆進行了深入研究。目前已克隆的鋅轉運相關蛋白基因有 OsZIP1、OsZIP3、OsZIP4、OsZIP5、OsZIP6、OsZIP8、OsIRT1、OsNRAMP3、OsMTP1、OsHMA2、OsHMA3、OsPDR9,基因分布如圖5所示。
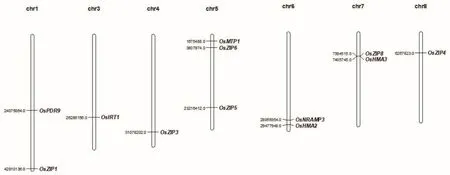
圖5 已克隆鋅轉運蛋白基因分布圖Fig.5 Distribution map of cloned zinc transporter
OsZIP1被認為是一種鋅吸收轉運體,也是一種限制水稻鋅過量積累的金屬外排轉運蛋白,其表達是在缺鋅的情況下誘導的[81]。OsZIP3主要在水稻根、葉處表達,在根中的表達受缺鋅誘導[86]。OsZIP4、OsZIP6、OsZIP8、在水稻地上部和根中表達,其表達是在缺鋅、鐵的情況下誘導的[76-77]。OsIRT1缺鐵時在水稻莖的韌皮部處表達[77]。OsHMA2、OsHMA3在缺鐵、鎘時分別在水稻根、葉處表達,都參與鋅和鎘的轉運[82-83]。OsPDR9在鋅、鎘脅迫下在根及幼苗處表達[78]。
鋅被水稻吸收后在根部積累,提高鋅向莖葉轉運效率成為關鍵。因此,對未克隆的相關基因(OsZIP2、OsZIP7、OsNRAMP2、OZT1、OsHMA1)進行功能分析將成為今后的研究重點。
7 展望
鋅對促進水稻生長發育,保障優質高產等方面具有重要作用,水稻所需的鋅主要從土壤中攝取,但由于我國水稻土資源豐富,類型復雜,不同種稻土中鋅含量分布差異性較大,且不同基因型水稻對鋅元素的吸收積累能力也不同,導致水稻對鋅的吸收水平和籽粒富鋅效果欠佳。因此今后的研究可以從以下幾個方面加強:
目前,專家學者對于植物營養與肥料的研究已經實現從定性方面深入到定量方面,根據不同水稻土的含鋅量和水稻的營養特點,制定測土施肥和植株診斷施肥方案,通過施肥定量的投入鋅營養,調節水稻生長和發育[87]。數字模型的應用可以滿足不同環境、不同生育期內水稻鋅營養的需求,做到定量調控,提高鋅肥利用效率,避免“奢侈吸收”和無效累積[88]。
2004年以來,中國作物營養強化項目(Harvest plus-China)一直將通過作物營養強化提高主要糧食作物中的微量營養元素含量(鐵、鋅、維生素A等)作為重要研究目標。但要實現這一目標,應從以下三個方面進行開展:一是集中人力、物力資源對現有水稻種質的鋅吸收積累能力進行評價,為水稻資源的合理應用提供理論依據;二是利用現代分子生物技術選育不改變外觀、口感和食味特性的鋅營養高效型的水稻品種,提高水稻對鋅的吸收積累,實現生物強化;三是通過提高稻米中鋅的絕對含量和降低營養抑制物質(植酸)的水平,提高水稻鋅的生物有效性,改善人類的鋅營養[36]。
隨著分子生物學的發展,越來越多參與水稻體內鋅吸收和解毒作用的蛋白被相繼發現,并對大多數鋅轉運蛋白家族成員的結構、功能、細胞定位和器官表達特異性等方面展開深入研究[77-80]。但仍存在以下問題:一是研究目標單一化,對多個鋅吸收轉運蛋白協同調控的研究較少。二是目前對水稻籽粒鋅積累調控基因的研究過少,已有研究表明鋅轉運體基因的過表達,會使地上部的Zn2+向根部遷移,從而導致鋅在籽粒富集減少,為了提高水稻的鋅含量水平,若通過更換一個特異性的啟動子,或發現一個組織特異性表達的基因來實現相關蛋白基因在籽粒中進行異位表達,達到籽粒富集鋅的目的,將是今后研究鋅轉運蛋白的方向之一。三是發現的鋅轉運蛋白成員中克隆報道不多,對未克隆的相關基因進行功能分析,豐富水稻中鋅元素吸收、轉運和分配機制的基因調控網絡,并為富鋅水稻分子育種研究提供理論依據。
隨著第二代測序技術的發展和鑒定技術的進步,全基因組關聯分析(genome-wide association study,GWAS)因關聯精度高,研究周期短必將成為發掘與水稻鋅含量相關遺傳變異位點的有效方法,獲取與營養相關的關鍵位點將成為可能,也為系統全面的認知鋅營養信號通路奠定了基礎[89-90]。通過GWAS分析和轉錄組分析,充分挖掘富鋅水稻種質資源,有助于了解水稻對相關性狀適應性的途徑和機制,也有助于設計和制定有效的育種策略,培育相關的富鋅水稻品種。這些結果將為在育種中提高水稻籽粒鋅含量和探索其遺傳機理提供線索[91-92]。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
文苑(2020年6期)2020-06-22 08:41:52
科技傳播(2019年22期)2020-01-14 03:06:54
文苑(2019年22期)2019-12-07 05:29:00
