河南新育小麥品種多酚氧化酶活性基因的檢測與分布
2022-06-24 15:12:05葛君任德超孟自力李愛霞陳杰
江蘇農業科學 2022年11期
葛君 任德超 孟自力 李愛霞 陳杰


摘要:為了解多酚氧化酶活性基因在河南新育小麥品種中的分布情況,選用123份河南新育小麥品種為試驗材料,利用功能標記PPO18、PPO16和PPO29對供試材料的Ppo-A1和Ppo-D1位點等位基因進行分子檢測。結果表明,在Ppo-A1位點上共檢測到2種等位基因Ppo-A1a和Ppo-A1b,分布頻率分別為63.4%和36.6%,以與高多酚氧化酶活性相關的等位基因Ppo-A1a分布為主;在Ppo-D1位點上共檢測到2種等位基因Ppo-D1a和Ppo-D1b,分布頻率分別為78.9%和21.1%,以與低多酚氧化酶活性相關的等位基因Ppo-D1a分布為主。在Ppo-A1和Ppo-D1這2個位點上,共檢測到4種等位基因組合Ppo-A1a/Ppo-D1a、Ppo-A1a/Ppo-D1b、Ppo-A1b/Ppo-D1a和 Ppo-A1b/Ppo-D1b,分布頻率分別為52.8%、10.6%、26.0%和10.6%,以與中等多酚氧化酶活性相關的等位基因組合Ppo-A1a/Ppo-D1a分布為主。此外,本研究篩選出的偃高161、商麥188和厚德麥971等32份攜帶等位基因組合Ppo-A1b/Ppo-D1a(與低多酚氧化酶活性相關)的小麥材料,可以為一線育種人員進行小麥品質色澤改良提供參考信息。
關鍵詞:小麥;多酚氧化酶;分子檢測;功能標記;等位基因
中圖分類號:S512.103 文獻標志碼:A
文章編號:1002-1302(2022)11-0037-06
收稿日期:2022-03-07
基金項目:國家現代農業產業技術體系建設專項(編號:CARS-03-31)。
作者簡介:葛 君(1981—),女,河南商丘人,助理研究員,主要從事小麥遺傳育種與栽培研究。E-mail:y2013g2015@163.com。
通信作者:李愛霞,高級農藝師,主要從事小麥遺傳育種與栽培研究。E-mail:spsnlkxymzl@163.com。
小麥面粉在加工過程中容易產生褐變,這不僅影響面制食品的外觀品相,而且還會對其營養價值造成一定影響。小麥籽粒中的多酚氧化酶是引起小麥面粉褐變的主要因素[1-3]。調控多酚氧化酶活性的基因主要位于小麥第2同源染色體上[4-7],Raman等在小麥2AL染色體上檢測到的主效QTL基因位點可以解釋82%~84%的表型變異[6];張立平等在小麥2DL染色體上檢測到的主效QTL基因位點可以解釋25.1%~29.1%的表型變異[7]。隨后,Sun等克隆出位于2AL染色體上的Ppo-A1基因,并開發設計了分子功能標記PPO18用于檢測等位基因Ppo-A1a和Ppo-A1b,其中等位基因Ppo-A1a與高多酚氧化酶活性相關,等位基因Ppo-A1b與低多酚氧化酶活性相關[8]。He等克隆出位于2DL染色體上的Ppo-D1基因,并開發設計了分子功能標記PPO16和PPO29用于檢測等位基因 Ppo-D1a和Ppo-D1b,其中等位基因Ppo-D1a與低多酚氧化酶活性相關,等位基因Ppo-D1b與高多酚氧化酶活性相關[9]。標記PPO18、PPO16和PPO29的準確性和可靠性又先后在我國不同麥區的材料中得到了驗證[10-12],隨后國內學者利用上述標記又在新疆、陜西、甘肅、黑龍江、四川和貴州小麥材料中進行了檢測應用[13-18]。
河南是我國小麥重要的生產省份之一,選育低多酚氧化酶活性的小麥品種有利于小麥面粉品質的色澤改良。本研究選用123份河南新育小麥品種為試驗材料,利用功能標記PPO18、PPO16和PPO29對供試材料的Ppo-A1和Ppo-D1位點等位基因進行分子檢測,以了解多酚氧化酶活性基因在河南新育成小麥品種中的分布情況,進而篩選出攜帶低多酚氧化酶活性基因的小麥資源材料,從而為河南省一線育種人員進行小麥品質色澤改良提供參考信息。
1 材料與方法
1.1 供試材料
供試材料為參加2020—2021年度河南省區域試驗的小麥品種,共計123份(表1)。這些材料經過2年品種比較試驗晉級而來,可以很客觀地反映河南當前的小麥育種情況。
1.2 基因組DNA提取
提取基因組DNA所用試劑的配制以及提取均參考袁謙等[19]和陳杰等[20]的方法進行。
1.3 分子標記檢測
選用標記PPO18對供試材料Ppo-A1位點的等位基因檢測,擴增出685 bp條帶的材料記為Ppo-A1a類型等位基因,擴增出876 bp條帶的材料記為Ppo-A1b類型等位基因;選用標記PPO16和PPO29對供試材料Ppo-D1位點的等位基因檢測,擴增出713 bp條帶的材料記為Ppo-D1a類型等位基因,擴增出490 bp條帶的材料記為Ppo-D1b類型等位基因。所選用標記的引物信息見表2。
利用ABI 9700型PCR儀器(美國伯樂公司生產)進行擴增,擴增產物用1.8%的瓊脂糖凝膠電泳分離,用0.6%的溴化乙錠染色,最后用AlphaImager HP型凝膠成像系統(美國Protein Simple公司生產)掃描成像保存。
1.4 數據分析
用Excel 2003軟件進行基礎數據處理和表格制作。208899EF-BB0E-4B04-BFD8-DA6257A58E44
2 結果與分析
2.1 Ppo-A1位點等位變異的分子檢測
針對Ppo-A1位點,利用標記PPO18對123份供試材料進行分子檢測。檢測結果(圖1)顯示,中農867、輪選128和苑豐16等78份材料可以擴增出685 bp的條帶,記為Ppo-A1a類型等位基因,分布頻率為63.4%;百農227、豫農526和鄭麥189等45份材料可以擴增出876 bp的條帶,記為Ppo-A1b類型等位基因,分布頻率為36.6%。從檢測結果分析可知:在123份供試材料的Ppo-A1位點上,以與高多酚氧化酶活性相關的等位基因Ppo-A1a分布為主。
2.2 Ppo-D1位點等位變異的分子檢測
針對Ppo-D1位點,利用標記PPO16和PPO29對123份供試材料進行分子檢測(圖2、圖3)。檢測結果顯示,育麥6號、育麥699和中農539等97份材料可以擴增出713 bp的條帶,記為Ppo-D1a類型等位基因,分布頻率為78.9%;偃高167、溫育919和天民188等26份材料可以擴增出490 bp的條帶,記為Ppo-D1b類型等位基因,分布頻率為21.1%。從檢測結果分析可知,在123份供試材料的Ppo-D1位點上,以與低多酚氧化酶活性相關的等位基因Ppo-D1a分布為主。
2.3 多酚氧化酶活性基因在河南小麥材料中的分布
在123份供試材料Ppo-A1和Ppo-D1位點上共檢測到4種類型的等位基因組合(表1),與低多酚氧化酶活性相關的等位基因組合Ppo-A1b/Ppo-D1a共檢測到32份,分布頻率為26%;與高多酚氧化酶活性相關的等位基因組合Ppo-A1a/Ppo-D1b共檢測到13份,分布頻率為10.6%;與中等多酚氧化酶活性相關的等位基因組合Ppo-A1a/Ppo-D1a和Ppo-A1b/Ppo-D1b分別檢測到65份和13份,分布頻率分別為52.8%和10.6%。從以上內容分析可知,在123份供試材料的Ppo-A1和Ppo-D1位點上,以與中等多酚氧化酶活性相關的等位基因組合Ppo-A1a/Ppo-D1a分布為主。
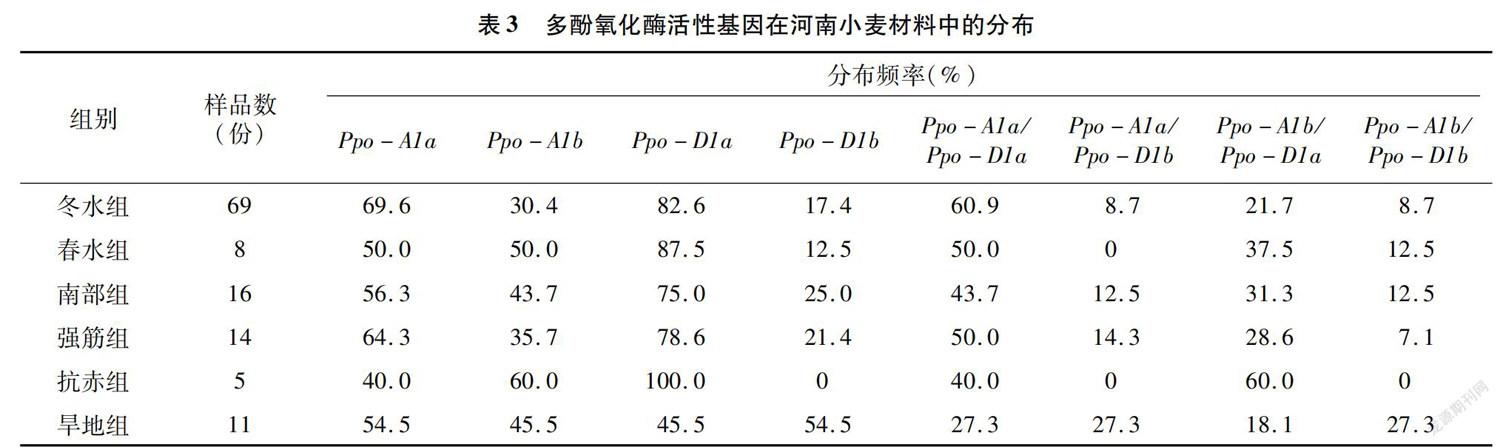
進一步分析不同類型河南小麥材料的多酚氧化酶基因的分布(表3)可知,與低多酚氧化酶活性相關的等位基因Ppo-A1b在來源于冬水組、南部組、強筋組、旱地組小麥材料中的分布頻率分別為30.4%、43.7%、35.7 %和45.5%,均低于與高多酚氧化酶活性相關的等位基因Ppo-A1a,而在抗赤組小麥材料中則反之。與低多酚氧化酶活性相關的等位基因Ppo-D1a在來源于冬水組、春水組、南部組、強筋組、抗赤組小麥材料中的分布頻率分別為82.6%、87.5%、75.0%、78.6 %和100.0%,均高于與高多酚氧化酶活性相關的等位基因Ppo-D1b,而在旱地組小麥材料中則反之。與低多酚氧化酶活性相關的等位基因組合Ppo-A1b/Ppo-D1a在來源于不同類型河南小麥材料中的分布頻率的大小順序為抗赤組(60.0%)>春水組(37.5%)>南部組(31.3%)>強筋組(28.6%)>冬水組(21.7%)>旱地組(18.1%)。從以上內容分析可知:與低多酚氧化酶活性相關的等位基因組合Ppo-A1b/Ppo-D1a在不同類型河南小麥材料中(除抗赤組)的分布頻率仍然較低。
3 討論與結論
了解多酚氧化酶活性基因的分布情況,可以為多酚氧化酶活性的遺傳改良提供參考信息。諸位國內學者研究表明,等位基因Ppo-A1a、Ppo-A1b、Ppo-D1a和Ppo-D1b在貴州小麥中的分布頻率分別為90.37%、9.63%、15.56%和84.44%,在四川小麥中的分布頻率分別為47.6%、52.4%、74.3%和25.7%,在黑龍江小麥中的分布頻率分別為60.8%、36.0%、48.0%和52.0%,在甘肅小麥中的分布頻率分別為49.04%、50.96%、50.96%和49.04%,在陜西小麥中的分布頻率分別為47.1%、52.9%、30.4%和69.6%,在新疆小麥中的分布頻率分別為69.0%、31.0%、13.1%和86.9%[13-18]。本研究表明,等位基因Ppo-A1a、Ppo-A1b、Ppo-D1a和Ppo-D1b在123份河南新育小麥品種中的分布頻率分別為63.4%、36.6%、78.9%和21.1%。綜合以上數據分析可知,與低多酚氧化酶活性相關的等位基因Ppo-A1b和Ppo-D1a在我國不同省份之間的分布頻率差異很大,等位基因Ppo-A1b在四川、甘肅和陜西小麥中的分布頻率均高于50%,而在貴州、黑龍江、新疆和河南小麥中的分布頻率均低于50%;等位基因Ppo-D1a在四川、甘肅和河南小麥中的分布頻率均高于50%,而在貴州、黑龍江、陜西和新疆小麥中的分布頻率均低于50%。今后在育種過程中應當對等位基因Ppo-A1b和 Ppo-D1a施加選擇壓力,淘汰攜帶等位基因Ppo-A1a和Ppo-D1b且綜合農藝性狀差的材料,選擇攜帶等位基因Ppo-A1b和Ppo-D1a且綜合農藝性狀好的材料,以促進低多酚氧化酶活性小麥新品種的選育。
分子功能標記的開發及應用是小麥分子育種領域的研究熱點。分子功能標記是根據不同位點等位基因內部的功能區開發設計,該功能區之間序列多態性的差異直接與所控制的性狀表型相關。因此,利用此項技術可大大提高分子標記輔助育種的準確性和效率。本研究選用的3個功能標記PPO18、PPO16和PPO29穩定性好,擴增條帶清晰,可以快速準確地檢測出Ppo-A1和Ppo-D1位點的等位基因變異情況,可直接作為多酚氧化酶活性基因分子輔助選擇的有效工具。此外,本研究篩選出了偃高161、商麥188和厚德麥971等32份攜帶等位基因組合Ppo-A1b/Ppo-D1a的小麥材料,在綜合考慮抗病性和產量等農藝性狀的前提下,可作為選育低多酚氧化酶活性品種的重點資源,在配置育種組合中加以利用。208899EF-BB0E-4B04-BFD8-DA6257A58E44
參考文獻:
[1]Simeone R,Pasqualone A,Clodoveo M L,et al. Genetic mapping of polyphenol oxidase in tetraploid wheat[J]. Cellular & Molecular Biology Letters,2002,7(2B):763-769.
[2]Baik B K,Czuchajowska Z,Pomeranz Y. Discoloration of dough for oriental noodles[J]. Cereal Chemistry,1995,72(2):198-205.
[3]Fuerst E P,Anderson J V,Morris C F. Delineating the role of polyphenol oxidase in the darkening of alkaline wheat noodles[J]. Journal of Agricultural and Food Chemistry,2006,54(6):2378-2384.
[4]Jimenez M,Dubcovsky J. Chromosome location of genes affecting polyphenol oxidase activity in seeds of common and durum wheat[J]. Plant Breeding,1999,118(5):395-398.
[5]Demeke T,Morris C,Campbell K G,et al. Wheat polyphenol oxidase:distribution and genetic mapping in three inbred line populations[J]. Crop Science,2001,41:1750-1757.
[6]Raman R,Raman H,Johnstone K,et al. Genetic and in silico comparative mapping of the polyphenol oxidase gene in bread wheat (Triticum aestivum L.)[J]. Functional & Integrative Genomics,2005,5(4):185-200.
[7]張立平,葛秀秀,何中虎,等. 普通小麥多酚氧化酶活性的QTL分析[J]. 作物學報,2005,31(1):7-10.
[8]Sun D J,He Z H,Xia X C,et al. A novel STS marker for polyphenol oxidase activity in bread wheat[J]. Molecular Breeding,2005,16(3):209-218.
[9]He X Y,He Z H,Zhang L P,et al. Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat[J]. Theoretical and Applied Genetics,2007,115(1):47-58.
[10]肖永貴,何心堯,劉建軍,等. 中國冬小麥品種多酚氧化酶活性基因等位變異檢測及其分布規律研究[J]. 中國農業科學,2008,41(4):954-960.
[11]左愛輝,陳 鋒,尚曉麗,等. 河南小麥新品種(系)的多酚氧化酶基因等位變異[J]. 麥類作物學報,2012,32(6):1072-1077.
[12]陳 杰,陳 鋒,詹克慧,等. 黃淮麥區小麥子粒多酚氧化酶活性基因等位變異的分子檢測[J]. 植物遺傳資源學報,2013,14(5):900-907.
[13]晏 權,任明見,李振華,等. 貴州小麥品種(系)籽粒低PPO活性種質資源篩選[J]. 南方農業學報,2020,51(3):512-519.
[14]李式昭,鄭建敏,伍 玲,等. 四川小麥品種淀粉特性、多酚氧化酶活性和黃色素含量相關基因的分子標記鑒定[J]. 分子植物育種,2015,13(5):982-993.
[15]楊淑萍,張宏紀,張舉梅,等. 俄引與黑龍江春小麥多酚氧化酶活性基因類型的比較[J]. 核農學報,2015,29(12):2268-2275.
[16]曹 東,張雪婷,王世紅,等. 104份甘肅小麥品種脂肪氧化酶和多酚氧化酶活性基因等位變異的檢測[J]. 麥類作物學報,2014,34(4):467-473.
[17]付曉潔,葉 石,吳學闖,等. 多酚氧化酶活性基因等位變異在陜西小麥品種中的分布[J]. 麥類作物學報,2009,29(2):236-240.
[18]王 亮,穆培源,徐紅軍,等. 新疆小麥品種中多酚氧化酶(PPO)活性基因等位變異的分布[J]. 麥類作物學報,2008,28(5):766-771.
[19]袁 謙,張 鋒,張中州,等. 國審小麥品種漯麥18重要功能基因的KASP標記檢測[J]. 江蘇農業科學,2021,49(24):56-59.
[20]陳 杰,張星宇,張福彥,等. 黃淮麥區(南片)小麥新品系脂肪氧化酶活性分析及其等位基因檢測[J]. 麥類作物學報,2021,41(10):1219-1227.208899EF-BB0E-4B04-BFD8-DA6257A58E44
