2016 和2020 年長(zhǎng)江口及鄰近海域浮游動(dòng)物群落結(jié)構(gòu)夏季年際變化及與環(huán)境因子的關(guān)系
2022-07-02 09:21:02胡亞麗王健鑫許永久蔣日進(jìn)朱文斌張洪亮
關(guān)鍵詞:優(yōu)勢(shì)差異
胡亞麗,王健鑫,許永久,鄭 基,蔣日進(jìn),朱文斌,張洪亮,郭 愛,梁 君
(1.浙江海洋大學(xué)水產(chǎn)學(xué)院,浙江舟山 316022;2.浙江海洋大學(xué)海洋科學(xué)與技術(shù)學(xué)院,浙江舟山 316022;3.浙江海洋大學(xué)海洋與漁業(yè)研究所,浙江省海洋水產(chǎn)研究所,浙江舟山 316021)
舟山地處長(zhǎng)江口和杭州灣,長(zhǎng)江和錢塘江等河流攜帶大量的陸源物質(zhì)進(jìn)入該海域,使得該海域初級(jí)生產(chǎn)力豐富,為浮游動(dòng)物提供豐富的餌料來(lái)源[1]。同時(shí),該地區(qū)受沿岸徑流、臺(tái)灣暖流與長(zhǎng)江沖淡水的多重影響,水文環(huán)境復(fù)雜多樣,每年夏季豐水期,長(zhǎng)江沖淡水勢(shì)力增強(qiáng),隨之帶來(lái)的半咸水或淡水種對(duì)于浮游動(dòng)物群落結(jié)構(gòu)亦具有顯著影響[2]。關(guān)于舟山近海浮游動(dòng)物數(shù)量變化的研究較多[3-10]對(duì)2000-2003 年長(zhǎng)江口鄰近海域春季和夏季浮游動(dòng)物群落特征的變動(dòng)趨勢(shì)的研究,表明鹽度是影響浮游動(dòng)物群落的主要環(huán)境因子;王慧君、蔣巧麗等[4-5]研究了舟山近海春季、秋季不同水域浮游動(dòng)物群落結(jié)構(gòu)的差異及其影響因素。
沈海琪等[2]分析了長(zhǎng)江口內(nèi)外浮游動(dòng)物群落結(jié)構(gòu)的晝夜差異,發(fā)現(xiàn)溫度、葉綠素a 的變化會(huì)影響到浮表層浮游動(dòng)物的群落結(jié)構(gòu)。胡亞麗等[10]研究了2019 年春、秋季浮游動(dòng)物群落結(jié)構(gòu)與環(huán)境因子的關(guān)系,得出鹽度是影響空間分布差異主要環(huán)境因子,溫度、葉綠素a 是影響季節(jié)分布差異的主要環(huán)境因子。然而受各航次調(diào)查所限,前述研究較少針對(duì)上升流、海洋鋒等物理海洋學(xué)因素對(duì)浮游動(dòng)物群落結(jié)構(gòu)的影響展開研究。夏季上升流是長(zhǎng)江口鄰近海域較為典型一種物理過程,其所帶來(lái)的低溫、高初級(jí)生產(chǎn)力水團(tuán)會(huì)對(duì)周邊的生物群落結(jié)構(gòu)產(chǎn)生重要影響。
本文結(jié)合2016 和2020 年夏季長(zhǎng)江口及其鄰近海域浮游動(dòng)物和水文調(diào)查,研究2 個(gè)不同年份上升流強(qiáng)弱及其對(duì)浮游動(dòng)物群落結(jié)構(gòu)組成,分析夏季長(zhǎng)江沖淡水量的變化對(duì)浮游動(dòng)物群落結(jié)構(gòu)的影響,從而為研究舟山近海浮游動(dòng)物生態(tài)系統(tǒng)動(dòng)力學(xué)提供一定的參考,也為舟山近海漁場(chǎng)生態(tài)修復(fù)提供理論基礎(chǔ)。
1 材料與方法
1.1 樣品采集與分析
2016 年8 月20-26 日(夏季)和2020 年8 月10-17 日(夏季),分別搭乘“浙嵊漁10301 號(hào)”和“浙漁科2號(hào)”開展浮游動(dòng)物生態(tài)和水文調(diào)查,2016 年的夏季調(diào)查區(qū)域?yàn)?0°-33° N,122°-127° E,調(diào)查站位為16個(gè)。2020 年的調(diào)查區(qū)域?yàn)?0°-32° N,122°-126° E,調(diào)查站位為19 個(gè)(圖1)。2016 和2020 年夏季浮游動(dòng)物樣品采集和樣品處理均按照《海洋調(diào)查規(guī)范海洋動(dòng)物調(diào)查》(GB/T 12763.6-2007)中的規(guī)定進(jìn)行,樣品采集使用淺水I 型浮游動(dòng)物網(wǎng)(口徑50 cm、篩絹CQ 14、網(wǎng)長(zhǎng)145 cm、孔徑0.505 mm),所獲樣品均5 %甲醛溶液固定保存后,帶回實(shí)驗(yàn)室內(nèi)通過顯微鏡分類、鑒定和計(jì)數(shù)。2016 年采集方法為距海面10 m 處向上斜平拖2 min,2020 年采集方法為表層5 m 左右平拖10 min。將2016 年和2020 年的浮游動(dòng)物數(shù)量轉(zhuǎn)換成單位體積尾數(shù)(浮游動(dòng)物豐度),進(jìn)行數(shù)據(jù)標(biāo)準(zhǔn)化,使得2 個(gè)不同年份的浮游動(dòng)物數(shù)據(jù)具有可比性和一致性。

圖1 2016(圖a)和2020 年(圖b)夏季浮游生物調(diào)查站位Fig.1 2016 (figure a) and 2020 (figure b) summer plankton survey stations
為了更好地識(shí)別長(zhǎng)江口及鄰近海域的上升流,本文使用了大尺度的衛(wèi)星遙感數(shù)據(jù),2016 和2020 年海水表層溫度(SST)與葉綠素a(Chl a)數(shù)據(jù)從OceanColor 官方網(wǎng)(http://www.oceancolor.org/)下載獲得,應(yīng)用WimSoft 軟件(www.wimsoft.com)計(jì)算該海域調(diào)查期間的平均值。
1.2 統(tǒng)計(jì)分析
1.2.1 群落結(jié)構(gòu)參數(shù)計(jì)算
(1)浮游動(dòng)物數(shù)據(jù)標(biāo)準(zhǔn)化
由于浮游動(dòng)物數(shù)據(jù)的差異性比較大,為了減少極值對(duì)數(shù)據(jù)的影響,將所有站位每種浮游動(dòng)物數(shù)量進(jìn)行標(biāo)準(zhǔn)化處理,公式如下[11]:
式中:Xi為原始豐度;Yi為轉(zhuǎn)換后數(shù)值。
(2)浮游動(dòng)物的豐度(N)和優(yōu)勢(shì)種(Y)豐度利用采樣時(shí)的濾水體積來(lái)計(jì)算,以每立方米水體中的個(gè)數(shù)(個(gè)·m-3)來(lái)表示[12]。豐度(N)計(jì)算公式為:
式中:N 為單位體積海水中浮游動(dòng)物的個(gè)體數(shù),ind.·m-3;V 為采集浮游動(dòng)物樣品中過濾海水體積,m3;n 為樣品鑒定計(jì)數(shù)獲得的浮游動(dòng)物個(gè)體數(shù)。2016 年體積V1計(jì)算公式為:V1=πr2×L1,L1=0.5 m·s-1×120 s=60 m,V1=11.775;2020 年體積V2計(jì)算公式為:V2=πr2×L2,L2=1.29 m·s-1×600 s=771 m,其中L1為2016 年浮游生物網(wǎng)拖行距離,L2為2020 年浮游生物網(wǎng)拖行距離,r 為網(wǎng)口半徑0.25 m。
優(yōu)勢(shì)種的計(jì)算公式為:

式中:N 為各采樣站位所有種類的豐度,ind.·m-3;ni為第i 種的豐度,ind.·m-3;fi為該種在各站位出現(xiàn)的頻率。以YI 0.02 為優(yōu)勢(shì)種的劃分標(biāo)準(zhǔn)[13-14]。
(3)浮游動(dòng)物及其優(yōu)勢(shì)種的更替率[15]計(jì)算公式

式中:a 和b 分別為2 個(gè)季節(jié)優(yōu)勢(shì)種或者浮游動(dòng)物種類的個(gè)數(shù);c 為2 個(gè)季節(jié)共同的優(yōu)勢(shì)種數(shù)或共同的浮游動(dòng)物種數(shù)。
1.2.2 統(tǒng)計(jì)分析
利用SPSS,根據(jù)年(year)、經(jīng)度(lon)、緯度(lat)、海水表層溫度(SST)、葉綠素a(Chl a),將2016 和2020 年的站位進(jìn)行聚類;采樣時(shí)間分年際尺度和晝夜尺度,晝夜根據(jù)調(diào)查時(shí)間點(diǎn)確定,06:00-18:00 為白天,18:00 到次日06:00 為夜間[2]。利用Primer 7.0 的SIMPER 方法計(jì)算不同年份和不同棲息地浮游動(dòng)物群落結(jié)構(gòu)的貢獻(xiàn)種和貢獻(xiàn)率,使用Primer 7.0 中的PERMANOVA(非參數(shù)方差分析)方法研究時(shí)間(年際尺度、晝夜尺度)和空間(近岸和近海)對(duì)2 年浮游動(dòng)物的群落結(jié)構(gòu)、多樣性、均勻度、豐富度的差異,時(shí)間尺度為隨機(jī)因子,空間尺度為固定因子。利用Canoco 5.0 軟件中的CCA 分析浮游動(dòng)物各類群以及優(yōu)勢(shì)種結(jié)構(gòu)特征年際變化與環(huán)境因子的關(guān)系。2 年的表層溫度、鹽度和葉綠素a 的等值線圖均采用Sufer13 軟件繪制。
2 結(jié)果
2.1 2016 和2020 年夏季水文環(huán)境
從遙感數(shù)據(jù)可以看出,從長(zhǎng)江口到舟山近岸再到舟山近海海水表層溫度呈現(xiàn)出高-低-高的趨勢(shì),研究區(qū)范圍內(nèi),空間變化表現(xiàn)為2016 年海水表層溫度變化范圍為29.4~31.5 ℃,最高值分布外海站位,最低值分布在近岸站位,平均值為30.4 ℃(圖2a);2020 年溫度的空間分布與2016 年相似,從長(zhǎng)江口到舟山近岸再到舟山近海也呈現(xiàn)出高-低-高的趨勢(shì),研究區(qū)范圍內(nèi)溫度為24.5~29.8 ℃,最高值分布在近海站位,最低值分布在近岸長(zhǎng)江口偏南站位,平均值為27.7 ℃(圖2b)。年際變化上表現(xiàn)出2020 年的平均溫度比2016 年低3 ℃左右。
2016 年和2020 年遙感葉綠素都呈現(xiàn)出由近岸到近海逐漸降低的趨勢(shì)。研究區(qū)域內(nèi),空間變化表現(xiàn)為2016 年葉綠素的范圍值為0.012~3.02,最高值分布在長(zhǎng)江口附近站位,最低值分布在長(zhǎng)江口外側(cè)(經(jīng)度124°E 附近),平均值為0.75(圖2c);2020 年葉綠素的范圍值為0.03~2.50,最高值和2016 年葉綠素a 最高值分布范圍相似,為長(zhǎng)江口附近站位,最低值分布范圍為研究區(qū)域南面(緯度30°N 附近),平均值為0.59(圖2d)。年際變化表現(xiàn)出2020 年葉綠素a 平均值比2016 年低0.2 左右。

圖2 2016 和2020 年研究區(qū)域環(huán)境Fig.2 Study regional environment in 2016 and 2020
2.2 2016 年和2020 年優(yōu)勢(shì)種類及浮游動(dòng)物種類組成
2.2.1 浮游動(dòng)物種類組成及優(yōu)勢(shì)種
2016 和2020 年共鑒定浮游動(dòng)物14 類147 種(表2),其中2016 年鑒定浮游動(dòng)物91 種,2020 年鑒定浮游動(dòng)物89 種,2 年的共有種33 種,約占2 年物種數(shù)的1/5,5 年間物種更替率為77.6%。2016 年夏季橈足類為第一優(yōu)勢(shì)類群,共鑒定44 種,占浮游動(dòng)物總種數(shù)的48.4%;浮游幼體11 種,占浮游動(dòng)物總種數(shù)的12.1%;水母類10 種,占浮游動(dòng)物總種數(shù)的11.0%。2020 年夏季橈足類為第一優(yōu)勢(shì)類群,共鑒定37 種,占浮游動(dòng)物總種數(shù)的41.6%,水母類為第二優(yōu)勢(shì)類群,共鑒定18 種,占浮游動(dòng)物總種數(shù)的20.2%;浮游幼體為第三優(yōu)勢(shì)類群,共鑒定8 種,占浮游動(dòng)物總種數(shù)的9.0%。

表2 2016 年和2020 年舟山近海浮游動(dòng)物種類組成Tab.2 The composition of zooplankton species in the coastal waters of Zhoushan in 2016 and 2020
優(yōu)勢(shì)度的分析結(jié)果顯示,2016 年共有8 個(gè)物種屬于優(yōu)勢(shì)種,其中前3 位是微刺哲水蚤Canthocalanus pauper、錐形寬水蚤Temora turbinata、普通波水蚤Undinula vulgaris,2020 年共有7 個(gè)物種屬于優(yōu)勢(shì)種,其中前3 位是雙生水母Diphyes chamissonis、長(zhǎng)尾類溞狀幼體、尖刺唇角水蚤L(zhǎng)abidocera acuta(表1)。

表1 2016 和2020 年浮游動(dòng)物優(yōu)勢(shì)種(類)及優(yōu)勢(shì)度Tab.1 Dominant species and dominance of zooplankton in 2016 and 2020

續(xù)表
2.3 浮游動(dòng)物群落結(jié)構(gòu)年際差異及其影響因素
2.3.1 浮游動(dòng)物年際差異的貢獻(xiàn)種和貢獻(xiàn)率
2016 和2020 年浮游動(dòng)物豐度分別為922.78 ind.·m-3和364.66 ind.·m-3。2016 年主要貢獻(xiàn)類群(>12%)為橈足類(38.07%)、浮游幼蟲(13.83%)和毛顎類(13.29%),2020 年主要為浮游幼蟲(17.73%)、橈足類(17.06%)和水母類(16.45%)。
2016 年和2020 年站位根據(jù)經(jīng)度(lon)、緯度(lat)、海水表層溫度(SST)、葉綠素a(Chla)環(huán)境因子通過聚類分析分為3 組(圖3),2016 年浮游動(dòng)物和2020 年浮游動(dòng)物群落結(jié)構(gòu)組成年際變化上存在顯著差異(ANOSIM,P<0.05),物種平均相異性為90.76 %,2 年浮游動(dòng)物年際差異的主要貢獻(xiàn)種為拿卡箭蟲(3.95%)、短尾類溞狀幼體(3.19%)、雙生水母(3.14%)、微刺哲水蚤(3.13%)、長(zhǎng)尾類溞狀幼體(3.08%);2016年近海和2020 年近海物種的平均相異性為89.66%,主要貢獻(xiàn)種為海樽(4.05%)、拿卡箭蟲(3.36%)、雙生水母(3.48%)、短尾類溞狀幼體(3.21%)、中型瑩蝦(3.07%)(表3);2020 年近岸浮游動(dòng)物和2020 年近海浮游動(dòng)物空間變化上存在顯著差異(ANOSIM,P<0.05),平均相異性為72.02%,空間差異的主要貢獻(xiàn)種為雙生水母(5.07%)、海樽(4.85%)、尖刺唇角水蚤(3.64%)、中型瑩蝦(3.43%)、強(qiáng)真哲水蚤(3.26%),這些貢獻(xiàn)種大部分都是來(lái)自2 年各自的優(yōu)勢(shì)類群和優(yōu)勢(shì)物種。
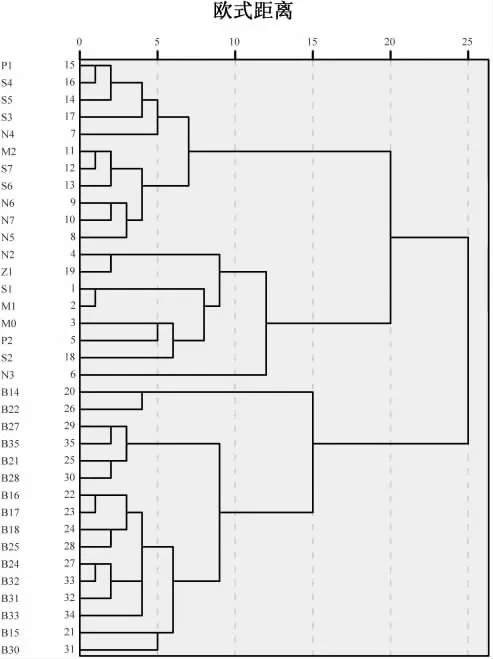
圖3 2016 和2020 年夏季站位聚類圖Fig.3 The cluster map of the stations in the summer of 2016 and 2020

表3 2016 和2020 年浮游動(dòng)物年際及空間差異的貢獻(xiàn)種和貢獻(xiàn)率Tab.3 Contributing species and contribution rates of interannual and spatial differences in zooplankton in 2016 and 2020
2.3.2 浮游動(dòng)物群落組成時(shí)間(晝夜、年份)及空間差異
2016 與2020 年晝、夜間采樣站位分別為8、8 個(gè)與14、5 個(gè),2016 年晝間采樣站位平均豐度為69.73 ind.·m-3,夜間采樣站位平均豐度為42.73 ind.·m-3,晝間平均豐度高于夜間,2020 年晝間平均豐度為32.16 ind.·m-3,夜間平均豐度為182.89 ind.·m-3,與2016 年相反,2020 年晝間平均豐度明顯低于夜間平均豐度。浮游動(dòng)物晝夜豐度有明顯差異,但浮游動(dòng)物晝夜群落結(jié)構(gòu)沒有明顯差異(PERMANOVA,P>0.05)。
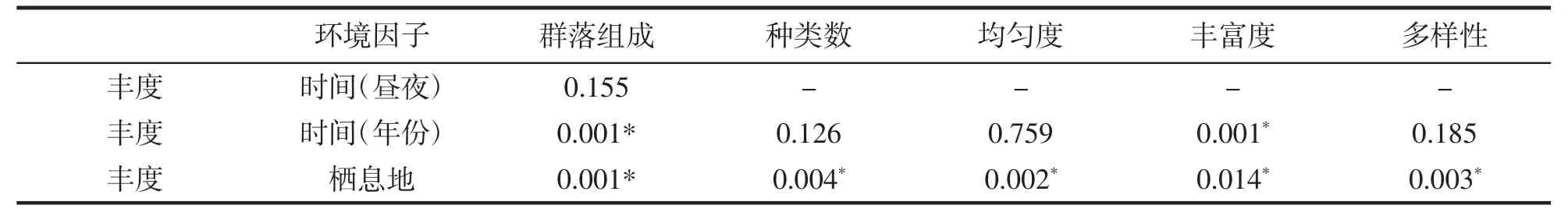
注:豐度經(jīng)過lg(數(shù)量+1)轉(zhuǎn)換,各指數(shù)豐度之間距離使用Bray-Curtis 相似性指數(shù);P 值經(jīng)過999 次迭代計(jì)算而來(lái),*極顯著差異[16].
表4 結(jié)果顯示,空間作為隨機(jī)因子,時(shí)間作為固定因子,年份和棲息地的差異對(duì)群落結(jié)構(gòu)組成的影響極顯著(P<0.01),年份差異對(duì)豐富度影響顯著,棲息地差異對(duì)種類數(shù)、均勻度、豐富度和多樣性影響較為顯著。
2.3.3 各類群浮游動(dòng)物群落結(jié)構(gòu)年際變化與環(huán)境因子的關(guān)系
2016、2020 兩年夏季各類群浮游動(dòng)物群落結(jié)構(gòu)與環(huán)境因子的CCA 分析顯示,前2 軸與環(huán)境因子之間相關(guān)性均顯著(圖4a、4b)。2 年的浮游動(dòng)物優(yōu)勢(shì)種與環(huán)境因子CCA 分析顯示,前4 軸與環(huán)境因子之間的相關(guān)性顯著(F=5.5,P<0.05),第一CCA 軸(CCA 1,特征值:0.59)和第二CCA 軸(CCA 2,特征值:0.48)表現(xiàn)出較高的物種-環(huán)境相關(guān)性,CCA 1 和CCA 2 解釋了物種累計(jì)方差百分比的88.6%和物種-環(huán)境累計(jì)方差百分比的95.8%(圖4d),Chl a、SST、Lon、Lat 均聚集在CCA 1 的負(fù)軸(圖4d)。2016 年站位和2020 年站位分別分布在CCA 1 的負(fù)軸和正軸,因此CCA 1 主要反應(yīng)了年際的差異。2020 年近岸站位和2020 年近海站位分別分布在CCA 2 的正軸和負(fù)軸,因此CCA 2 正負(fù)軸兩邊呈現(xiàn)了近岸站位和近海站位的差異(圖4d)。這與2016 和2020 各類群浮游動(dòng)物與環(huán)境因子的CCA 分析圖結(jié)果一致(圖4c),錐形寬水蚤聚集在CCA 1 的負(fù)軸,與海水表層溫度(SST)呈現(xiàn)明顯的正相關(guān)性,說明海水表層溫度(SST)是影響其分布的水域的主要因素,長(zhǎng)尾類溞狀幼體分布在CCA 1 的正軸,CCA 2 軸的正軸,與海水表層溫度(SST)呈現(xiàn)負(fù)相關(guān)性,微刺哲水蚤和普通哲水蚤聚聚在CCA 2 的正軸,說明高Chl a 濃度是影響其分布的主要因素。而長(zhǎng)尾類溞狀幼體和雙生水母聚集在CCA 1 的正軸,與Chl a 呈現(xiàn)負(fù)相關(guān),這些分布特征可能反映出浮游動(dòng)物對(duì)某一環(huán)境因子的偏好和其生態(tài)習(xí)性。
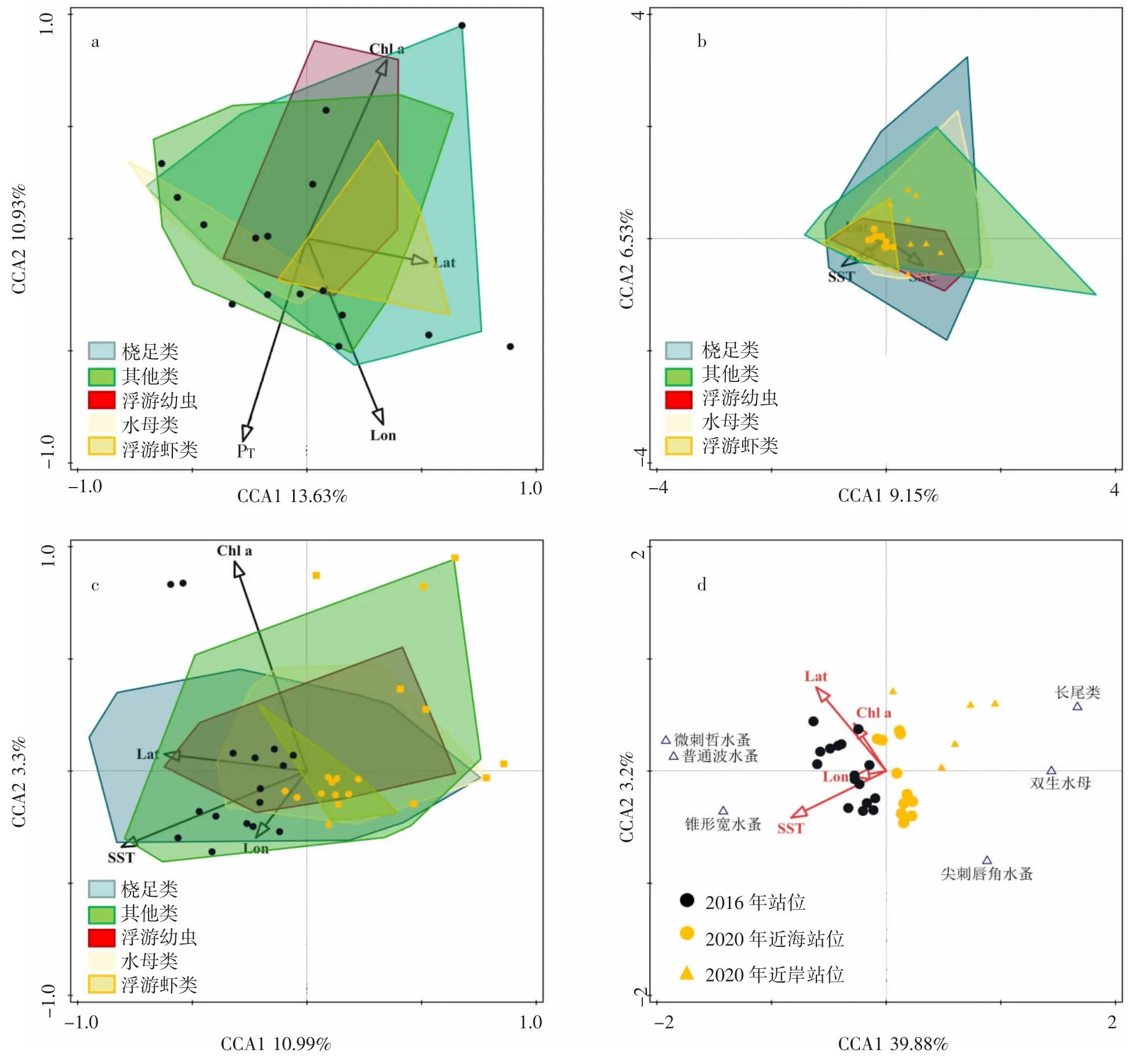
圖4 浮游動(dòng)物與環(huán)境因子典范對(duì)應(yīng)分析Fig.4 Canonical correspondence analysis of dominant species of zooplankton and environmental factors
3 討論
3.1 長(zhǎng)江口沖淡水對(duì)浮游動(dòng)物群落組成的影響
長(zhǎng)江口上升流強(qiáng)弱一般通過上升流與非上升流區(qū)溫度差值來(lái)表示,從圖2 中可以看出2016 年上升流與非上升流區(qū)溫度差值有6 ℃左右,2020 年則相差4 ℃左右,因此2016 年的整體溫度偏高,上升流較強(qiáng),2020 年的整體溫度稍低,上升流則相對(duì)較弱。兩年溫度差異對(duì)浮游動(dòng)物群落組成造成一定程度的影響,2016 年夏季和2020 年夏季浮游動(dòng)物的群落組成存在顯著差異(PERMANOVA,P<0.01),2016 年微刺哲水蚤數(shù)量較多,在B17 站位出現(xiàn)最高值,微刺哲水蚤為高溫高鹽種類,所適的水溫一般高于20 ℃,鹽度范圍在31~33 之間[17],這與2016 年溫度較高的環(huán)境狀況相吻合。而2020 年第一優(yōu)勢(shì)種則為雙生水母,這種演替方式可能與餌料競(jìng)爭(zhēng)或者捕食者等生物因素有關(guān),同樣受到環(huán)境因子的強(qiáng)烈影響[2]。2016 年沖淡水影響較強(qiáng),范圍偏北,臺(tái)灣暖流勢(shì)力入侵到長(zhǎng)江口,2020 年沖淡水較弱,影響范圍偏南,臺(tái)灣暖流影響勢(shì)力減弱,僅影響到舟山群島本島以南。因此2020 年的優(yōu)勢(shì)種為舟山近海常見種,在舟山近海區(qū)域分布較多[18-20]。高倩等[21]在2002-2003 年長(zhǎng)江口夏季的浮游水母的研究中也認(rèn)為雙生水母為當(dāng)年夏季的優(yōu)勢(shì)種,覃濤等[22]在對(duì)2006 和2007 年舟山近海的浮游動(dòng)物的研究結(jié)果中則認(rèn)為微刺哲水蚤為該海域的優(yōu)勢(shì)種,而楊杰青[23]在2013 年夏季舟山近岸海域浮游動(dòng)物的調(diào)查研究中并未得出微刺哲水蚤為優(yōu)勢(shì)種的結(jié)果。根據(jù)何青青[24]在2002-2013 年舟山近海夏季上升流時(shí)空特征的研究結(jié)果,認(rèn)為2013 年舟山近海的上升流并不顯著,海水表層溫度平均值為26 ℃左右。因此長(zhǎng)江口舟山近海海域這種優(yōu)勢(shì)種的年際演替受上升流、沖淡水和不同水團(tuán)的綜合交互影響。
3.2 棲息地、晝夜變化對(duì)浮游動(dòng)物優(yōu)勢(shì)種數(shù)量動(dòng)態(tài)變化的影響
2020 年近岸與近海站位浮游動(dòng)物群落組成和豐度均有顯著差異(P<0.05),近岸站位人類活動(dòng)頻繁[25],并且受到長(zhǎng)江沖淡水的影響,多為中華哲水蚤、中型瑩蝦等廣溫廣鹽種。近海受到臺(tái)灣暖流等外海水團(tuán)的影響,溫度和鹽度的值都逐漸升高,則多分布為尖刺唇角水蚤等高溫高鹽種和海樽、小哲水蚤等暖水種、大洋廣布種。且大多分布在123° E 以東高溫高鹽區(qū)域,這種明顯的種類分布差異也體現(xiàn)了不同棲息地對(duì)浮游動(dòng)物的生存空間的影響。
2016 和2020 年浮游動(dòng)物群落組成晝夜分布未有顯著差異(P>0.05),然而本文發(fā)現(xiàn)浮游動(dòng)物晝夜豐度存在明顯差異,2016 年白天浮游動(dòng)物豐度高于夜間,這一結(jié)果的可能是2016 年水深相對(duì)2020 年較深,由于夏季長(zhǎng)江口不同深度水溫的不同和浮游植物的分層,造成浮游動(dòng)物出現(xiàn)明顯的垂直分布[26],浮游動(dòng)物聚集在不同水深層,朱駿華[27]和陳輝煌[28]等在各自的研究中發(fā)現(xiàn)不同水層的浮游動(dòng)物豐度會(huì)有顯著差異,夏季浮游動(dòng)物的垂直分布差異較為顯著。2016 年采樣的浮游動(dòng)物可能為較深水層采集而來(lái)。2020 年夜間浮游動(dòng)物豐度遠(yuǎn)高于晝間浮游動(dòng)物豐度,這與KIBIRIGE,et al[29]的研究結(jié)果相一致,這一結(jié)果的可能原因是一種逃避捕食的生存策略,夜晚水體表層的攝食壓力比白天低[30],具體的浮游動(dòng)物垂直晝夜分布機(jī)制,還需要結(jié)合其他年份的調(diào)查數(shù)據(jù),進(jìn)一步研究驗(yàn)證。
致謝:感謝“浙嵊漁10301 號(hào)”和“浙漁科2 號(hào)”全體船員在出海采樣過程中的幫助,感謝A18 資環(huán)姜淇俊、吳麗航、徐紀(jì)曉、李立華、楊麗萍等同學(xué)參與樣品鑒定等部分實(shí)驗(yàn),謹(jǐn)致謝忱。
猜你喜歡
我愛學(xué)·笑話與口才(2025年3期)2025-02-24 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
趣味(語(yǔ)文)(2020年3期)2020-07-27 01:42:46
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國(guó)衛(wèi)生(2015年2期)2015-11-12 13:13:54
鄭州大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
中國(guó)火炬(2014年11期)2014-07-25 10:31:58
