斑鱾dmrt1的克隆及其表達(dá)分析
2022-07-04 09:20:00溫思民朱春華李廣麗
海洋漁業(yè) 2022年2期
溫思民,翟 毅,黃 洋,2,朱春華,2,4,李廣麗,2,3
(1.廣東海洋大學(xué)水產(chǎn)學(xué)院,廣東湛江 524088;2.廣東省名特優(yōu)魚(yú)類(lèi)生殖調(diào)控與繁育工程技術(shù)研究中心,廣東湛江 524088;3.廣東省水產(chǎn)經(jīng)濟(jì)動(dòng)物病原生物學(xué)及流行病學(xué)重點(diǎn)實(shí)驗(yàn)室,廣東湛江 524088;4.廣東省海水養(yǎng)殖生物育種工程實(shí)驗(yàn)室,廣東湛江 524088)
dmrt (doublesex and mab-3 related transcription factors)基因家族是一類(lèi)與果蠅(Drosophila melanogaste)性別決定基因 dsx(doublesex)和秀麗隱桿線蟲(chóng)(Caenorhabditis elegans)性別決定基因mab-3(male abnormal-3)同源的基因[1-2]。該基因家族成員的主要特征是所編碼的蛋白質(zhì)包含一個(gè)能與DNA序列結(jié)合,且高度保守的DM 結(jié)構(gòu)域(doublesex and male aberrant-3 relative domain),這些蛋白質(zhì)可通過(guò)其鋅指結(jié)構(gòu)與特定的DNA序列結(jié)合,調(diào)控下游基因的表達(dá),在性別決定與分化、胚胎發(fā)育、組織器官形成等生物學(xué)過(guò)程起著重要作用[3-4]。作為dmrt基因家族的一員,dmrt1基因編碼的蛋白質(zhì)具有典型的DM結(jié)構(gòu)域,該基因在無(wú)脊椎動(dòng)物和脊椎動(dòng)物中均存在,是參與性別決定的最古老的基因,具有進(jìn)化保守性。dmrt1基因廣泛存在于哺乳動(dòng)物、鳥(niǎo)類(lèi)、爬行類(lèi)和魚(yú)類(lèi)中,參與動(dòng)物性別決定與分化,在雄性性腺的表達(dá)水平高于雌性[5]。在人類(lèi)中,DMRT1基因在男性胚胎的生殖嵴中特異表達(dá),在女性中不表達(dá),該基因在維持睪丸的細(xì)胞形態(tài)和功能方面發(fā)揮重要作用,缺失會(huì)導(dǎo)致發(fā)生睪丸向卵巢的轉(zhuǎn)分化[6]。在小鼠中,Dmrt1基因在XX和XY早期胚胎的生殖嵴表達(dá),但在性別決定后逐漸轉(zhuǎn)變?yōu)橹辉诓G丸中特異表達(dá),敲除Dmrt1基因的XY雄性小鼠會(huì)導(dǎo)致睪丸分化受阻,最終喪失生殖細(xì)胞[7],而XX雌性小鼠Dmrt1基因的突變雖然會(huì)減少鼠胚卵巢中原始卵泡的形成,但不影響其生育能力[8]。在魚(yú)類(lèi)中,dmrt1基因最早在尼羅羅非魚(yú)(Oreochromis niloticus)和虹鱒(Oncorhynchus mykiss)中得到鑒定,并且僅在這兩個(gè)物種的精巢中表達(dá),表明該基因是重要的雄性相關(guān)基因[9-10]。XY雄性尼羅羅非魚(yú)dmrt1功能缺陷會(huì)導(dǎo)致其精巢退化,表現(xiàn)為輸精管畸形、精原細(xì)胞退化甚至完全喪失生殖細(xì)胞[11]。MATSUDA 等[12]和NANDA 等[13]在2002年分別獨(dú)立在青鳉(Oryzias latipes)中發(fā)現(xiàn)了魚(yú)類(lèi)的第一個(gè)雄性性別決定基因,即Y染色體上的DM結(jié)構(gòu)域基因(DM-domain gene on the Y chromosome,dmy),也稱(chēng)Y染色體性別決定區(qū)dmrt1的重復(fù)拷貝基因dmrt1bY,dmy基因的自然突變會(huì)導(dǎo)致XY青鳉發(fā)生由雄性向雌性的性逆轉(zhuǎn),而在XX雌性青鳉中過(guò)表達(dá)dmy會(huì)誘導(dǎo)發(fā)生卵巢向精巢的轉(zhuǎn)分化。此外,敲除常染色體dmrt1基因的XY青鳉會(huì)發(fā)育為雌性并具有正常生育能力,而通過(guò)挽救實(shí)驗(yàn)過(guò)表達(dá)的dmrt1,可使其發(fā)育為正常雄性[14]。與青鳉類(lèi)似,敲除dmrt1的XY雄性斑馬魚(yú)(Danio rerio)大多數(shù)發(fā)育為具有生育能力的雌性,少數(shù)成為不育的雄性,表現(xiàn)為精巢發(fā)育受阻,最終喪失生殖細(xì)胞[15]。以上研究表明,dmrt1基因參與脊椎動(dòng)物性別分化和精巢的功能調(diào)控。
斑鱾(Girella punctata),又稱(chēng)瓜子鱲,俗名黑毛魚(yú),隸屬于鱸形目(Perciformes),鱸亞目(Percoidei),鱾科(Girellidae),鱾屬,是近海暖水性巖礁魚(yú)類(lèi),食性為雜食性,主要以藻類(lèi)和小型無(wú)脊椎動(dòng)物為食,體長(zhǎng)最長(zhǎng)可達(dá)50 cm。斑鱾屬雌雄異體魚(yú)類(lèi),雌雄魚(yú)表型基本相同,不易分辨,但雌雄性腺發(fā)育不一致,雄魚(yú)需2年性成熟,雌魚(yú)需3年性成熟,該魚(yú)的產(chǎn)卵期在2—6月,棲息于巖礁、石礫底質(zhì)的近岸水域,主要分布于中國(guó)的東海、南海和臺(tái)灣島附近海域,以及朝鮮、日本和菲律賓沿海[16-18]。斑鱾?cè)赓|(zhì)潔白鮮嫩、營(yíng)養(yǎng)豐富,經(jīng)濟(jì)價(jià)值極高,是名貴高檔的經(jīng)濟(jì)魚(yú)類(lèi),深受消費(fèi)者青睞,具有十分廣闊的市場(chǎng)前景。但目前斑鱾尚不能進(jìn)行規(guī)模化人工繁殖,市場(chǎng)消費(fèi)的成魚(yú)及海水網(wǎng)箱養(yǎng)殖的斑鱾苗種均主要來(lái)自海捕,這在一定程度上制約了斑鱾規(guī)模化養(yǎng)殖的發(fā)展。dmrt1基因作為魚(yú)類(lèi)的重要生殖發(fā)育調(diào)控基因,在斑鱾中尚未見(jiàn)相關(guān)研究報(bào)道。本研究以斑鱾為研究對(duì)象,克隆其dmrt1基因,運(yùn)用生物信息學(xué)技術(shù)預(yù)測(cè)其Dmrt1蛋白的性質(zhì)與結(jié)構(gòu),并分析dmrt1基因在斑鱾雌雄性腺和胚胎發(fā)育過(guò)程中的表達(dá)模式,以期為斑鱾生殖發(fā)育調(diào)控及性別分化研究提供基礎(chǔ)資料。
1 材料與方法
1.1 實(shí)驗(yàn)動(dòng)物及樣品采集
斑鱾來(lái)自珠海育成魚(yú)苗養(yǎng)殖有限公司,體質(zhì)量230~347 g,體長(zhǎng)23.9~28.2 cm,3雌3雄。麻醉后取部分性腺組織液氮速凍后-80℃保存,用于提取總RNA及后續(xù)基因克隆;剩余部分性腺組織用Bouin’s液固定,乙醇脫水,二甲苯透明,石蠟包埋、切片,蘇木精-伊紅(HE)染色后用于性腺組織學(xué)觀察。斑鱾胚胎樣品取自中國(guó)臺(tái)灣省臺(tái)南市屏東縣東港鎮(zhèn)的中國(guó)臺(tái)灣水產(chǎn)試驗(yàn)所東港分所,受精卵在溫度為(26.0±0.5)℃、鹽度為31的海水中培育。用120目篩絹網(wǎng)撈取受精卵,用體視顯微鏡(Thermo,XTL-2400,美國(guó))進(jìn)行胚胎發(fā)育觀察及圖片采集,并取不同發(fā)育時(shí)期胚胎樣品各30顆液氮速凍后-80℃保存,用于總RNA提取。
1.2 總RNA提取和cDNA合成
采用Trizol試劑盒(Invitrogen,15596018,美國(guó))分別提取胚胎和性腺的總RNA。用1%瓊脂糖凝膠電泳檢測(cè)RNA的完整性,并用Nanodrop 2000超微量核酸蛋白測(cè)定儀(Thermo Scientific,美國(guó))檢測(cè)RNA的濃度和質(zhì)量。RNA質(zhì)量檢測(cè)合格后,按照PrimeScriptTMRTreagent Kit with gDNA Eraser(TaKaRa,RR047A,日本)說(shuō)明書(shū)合成cDNA。
1.3 斑鱾dmrt1 ORF的克隆與序列分析
從NCBI數(shù)據(jù)庫(kù)中獲得黃金鱸(Perca flavescens)和白梭吻鱸(Sander lucioperca)等鱸形目物種dmrt1 cDNA序列,在本實(shí)驗(yàn)室斑鱾性腺轉(zhuǎn)錄組數(shù)據(jù)庫(kù)(BioProject ID:PRJNA752004)中進(jìn)行生物信息學(xué)比對(duì)分析,找到斑鱾dmrt1可能轉(zhuǎn)錄本(7 204 bp),用Primer Premier 5.0軟件根據(jù)轉(zhuǎn)錄本序列設(shè)計(jì)特異性引物對(duì)該基因ORF進(jìn)行克隆驗(yàn)證,引物由上海生工生物技術(shù)公司合成(表1)。采用2×PCR MIX(P2014,東盛生物科技公司)進(jìn)行基因擴(kuò)增。PCR反應(yīng)程序:94℃預(yù)變性5 min;94℃30 s,54℃30 s,72℃1 min,32個(gè)循環(huán);72℃延伸10 min。將得到的PCR產(chǎn)物進(jìn)行1.5%瓊脂糖凝膠電泳檢測(cè),用DNA凝膠回收試劑盒(N1072,東盛生物科技公司)對(duì)目的條帶進(jìn)行切膠回收、連接、亞克隆、挑選陽(yáng)性克隆送至上海生工生物有限公司測(cè)序。
使用NCBI的ORF finder(https://www.ncbi.nlm.nih.gov/orffinder/)在線預(yù)測(cè)斑鱾dmrt1開(kāi)放閱讀框(open reading frame,ORF);使用在線分析軟件ExPASy Proteomics Server(http://ca.expasy.org)推導(dǎo)其氨基酸序列、分子量計(jì)算值和理論等電點(diǎn)等;使用SignalP 5.0 Server(http://www.cbs.dtu.dk/services/SignalP)預(yù)測(cè)其信號(hào)肽序列;使用TMHMM Server 2.0(http://www.cbs.dtu.dk/services/TMHMM)預(yù)測(cè)其跨膜結(jié)構(gòu)域;使用SoftBerry-Psite(http://linux1.softberry.com)預(yù)測(cè)其氨基酸序列中的功能位點(diǎn)分布;使用SMART(http://smart.embl-heidelberg.de)分析其蛋白質(zhì)結(jié)構(gòu)功能域;使用PSORT II Prediction(http://psort.hgc.jp/form2.html)預(yù)測(cè)其亞細(xì)胞定位;使用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)預(yù)測(cè)Dmrt1蛋白的二級(jí)結(jié)構(gòu);運(yùn)用ExPASy的SWISSMODEL(http://www.swissmodel.expasy.org)進(jìn)行建模分析;采用NCBI在線序列比對(duì)(http://blast.ncbi.nlm.nih.gov/Blast.cgi)進(jìn)行序列同源性比對(duì)和相似性分析;運(yùn)用DNAMAN 9進(jìn)行蛋白序列的多重比對(duì)分析;采用MEGA X軟件以鄰位相連法(neighbor-joining)構(gòu)建各物種Dmrt1系統(tǒng)進(jìn)化樹(shù)[19]。
1.4 實(shí)時(shí)熒光定量PCR檢測(cè)
采用實(shí)時(shí)熒光定量PCR(qRT-PCR)技術(shù)檢測(cè)dmrt1在雌雄性腺和不同胚胎發(fā)育時(shí)期的表達(dá)。操作步驟參照SYBR?Green Real Time PCR Master Mix試劑盒(TOYOBO,QPK-201,日本)說(shuō)明書(shū)進(jìn)行。反應(yīng)體系為:SYBR?Green Real Time PCR Master Mix 10μL,cDNA 2μL,10μmol·L-1上下游引物各0.8μL,再用超純水補(bǔ)齊至20μL。設(shè)置反應(yīng)程序:95℃預(yù)變性5 min;95℃30 s,54℃20 s,72℃20 s,40個(gè)循環(huán);72℃延伸5min。以β-actin為內(nèi)參基因,引物見(jiàn)表1。用2-ΔΔCT方法計(jì)算dmrt1在斑鱾雌雄性腺和不同胚胎發(fā)育時(shí)期中的相對(duì)表達(dá)量。
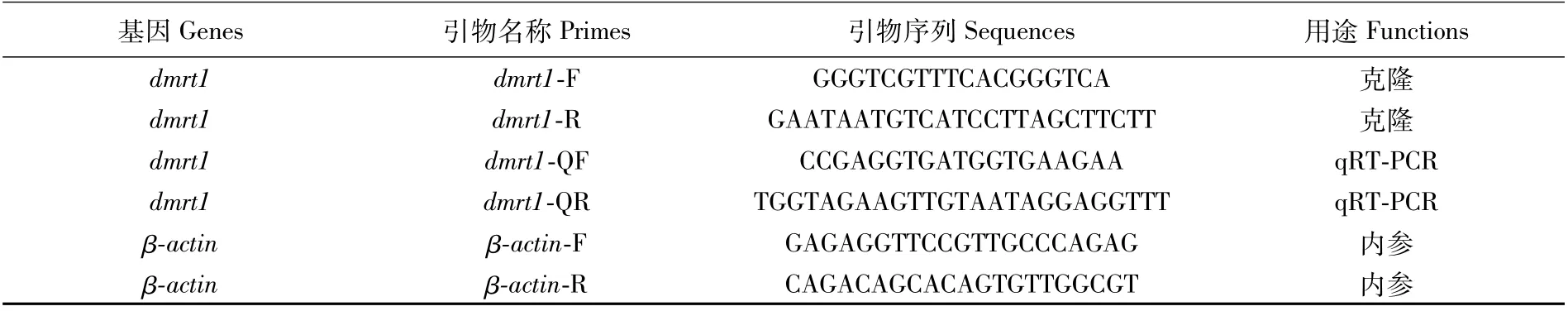
表1 PCR相關(guān)引物序列Tab.1 Primers used for PCR
1.5 數(shù)據(jù)處理
用SPSS 25.0對(duì)dmrt1在斑鱾雌雄性腺的表達(dá)數(shù)據(jù)進(jìn)行獨(dú)立樣本t檢驗(yàn)分析,對(duì)dmrt1在斑鱾不同胚胎發(fā)育時(shí)期的表達(dá)數(shù)據(jù)進(jìn)行單因素方差分析和Duncan多重比較分析,數(shù)據(jù)用平均值±標(biāo)準(zhǔn)誤差(mean±SE)表示,n=3,當(dāng)P<0.05時(shí)表示組間差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果與分析
2.1 斑鱾dmrt1 ORF的克隆及生物信息學(xué)分析
克隆得到的斑鱾dmrt1 ORF全長(zhǎng)為894 bp,編碼297個(gè)氨基酸(圖1)。對(duì)斑鱾Dmrt1蛋白進(jìn)行預(yù)測(cè)分析,結(jié)果表明該蛋白原子總數(shù)為4 414,分子結(jié)構(gòu)式為C1374H2154N406O457S23,理論分子量為32.41 ku,理論等電點(diǎn)(pI)為7.01;不穩(wěn)定系數(shù)64.19,大于閾值40,性質(zhì)不穩(wěn)定;脂溶指數(shù)為53.60,總平均親水性(GRAVY)為-0.638,屬親水性蛋白,位于細(xì)胞核,不存在信號(hào)肽與跨膜區(qū),含有4個(gè)N-糖基化位點(diǎn)、1個(gè)蛋白激酶C磷酸化位點(diǎn)、3個(gè)酪蛋白激酶II磷酸化位點(diǎn)、6個(gè)N-肉豆蔻酰化位點(diǎn)、2個(gè)異戊二烯基結(jié)合位點(diǎn)和4個(gè)微體C末端靶信號(hào)位點(diǎn)(圖1)。在蛋白N端27~80位具有一個(gè)與果蠅Dxs和線蟲(chóng)Mab-3蛋白相似的DM結(jié)構(gòu)域,用SOPMA軟件預(yù)測(cè)Dmrt1蛋白的二級(jí)結(jié)構(gòu),結(jié)果顯示,無(wú)規(guī)則卷曲所占的比例最高,占64.65%,延 伸 鏈 占14.14%,α-螺 旋 占17.51%,蛋白質(zhì)的二級(jí)結(jié)構(gòu)中有超過(guò)半數(shù)的氨基酸參與形成無(wú)規(guī)則卷曲結(jié)構(gòu),說(shuō)明該蛋白質(zhì)整體處于一個(gè)以無(wú)規(guī)則卷曲為主的二級(jí)結(jié)構(gòu)中。將Dmrt1氨基酸序列提交至SWISS-MODEL軟件,自動(dòng)搜索同源蛋白作模板,得到以人類(lèi)DMRT1的DM結(jié)構(gòu)域模型(PDB ID:4yj0.1.A)為模板構(gòu)建的斑鱾Dmrt1的DM 結(jié)構(gòu)域模型(圖2),相似性為87.10%,這表明了Dmrt1的DM結(jié)構(gòu)域在進(jìn)化上的保守性。

圖1 斑鱾dmrt1的核苷酸序列及其編碼的氨基酸序列Fig.1 Nucleic acid and amino acid sequences of dmrt1 gene in G.punctata
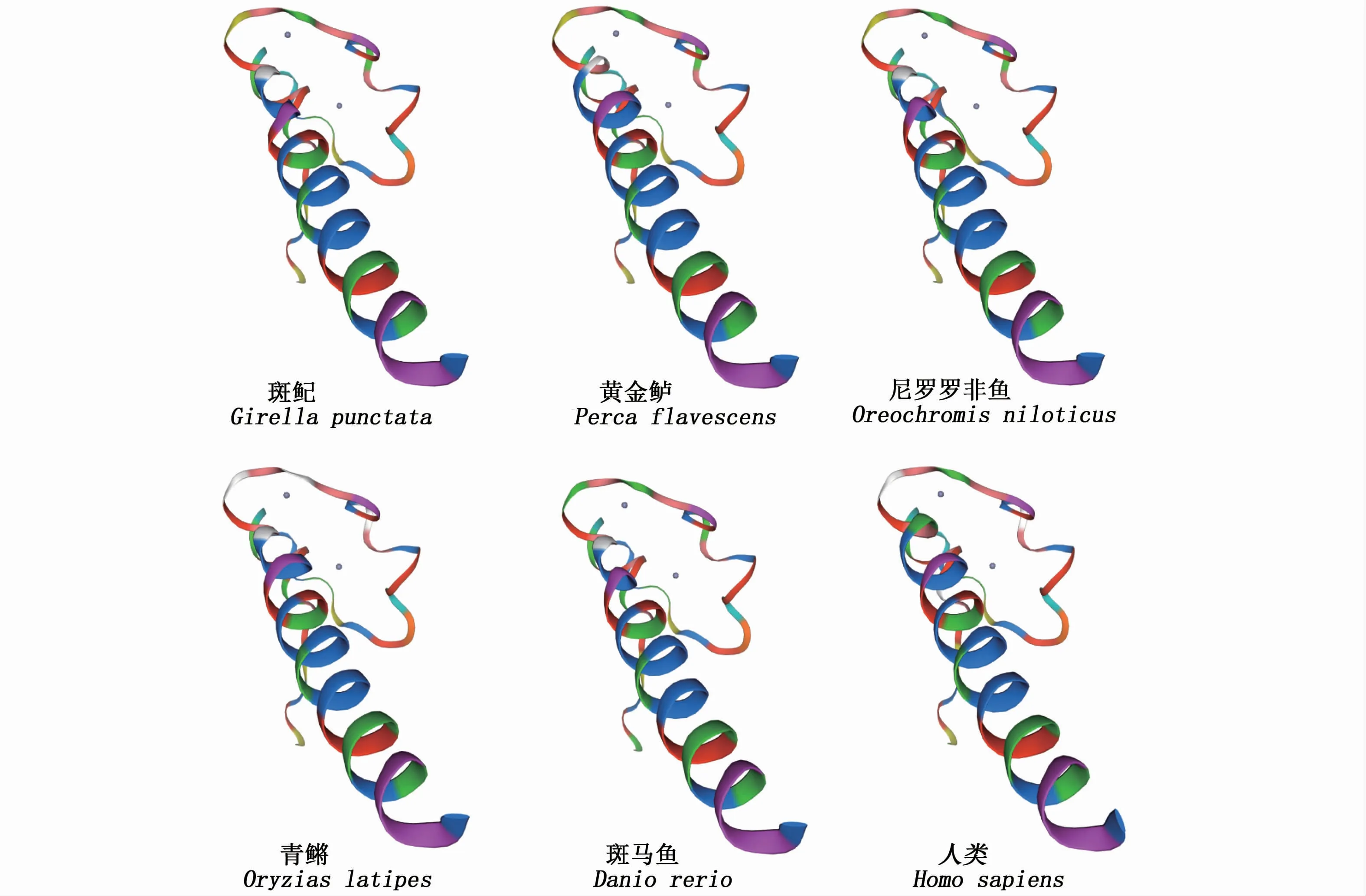
圖2 不同魚(yú)類(lèi)和人類(lèi)的Dmrt1/DMRT1蛋白的DM 結(jié)構(gòu)域模型Fig.2 DM domain model of Dmrt1/DMRT1 protein in different fishes and Homo sapiens
2.2 斑鱾Dmrt1的同源性及進(jìn)化分析
將斑鱾Dmrt1蛋白序列與其他物種的蛋白序列進(jìn)行同源比對(duì)分析,斑鱾Dmrt1與黃金鱸的同源性最高,為85.71%,與人類(lèi)的同源性最低,為32.89%(表2)。多重序列比對(duì)分析發(fā)現(xiàn),斑鱾和其他脊椎動(dòng)物的Dmrt1均含有保守的DM結(jié)構(gòu)域(圖3),斑鱾Dmrt1的DM結(jié)構(gòu)域與黃金鱸、白梭吻鱸、橙胸鏢鱸(Etheostoma obama)和歐洲海鱸(Dicentrarchus labrax)的同源性為96.30%,與斑馬魚(yú)的為90.74%,與人類(lèi)的為87.10%,DM結(jié)構(gòu)域的高度同源性表明了其在進(jìn)化上的保守性。

圖3 斑鱾與其他物種的Dmrt1的蛋白序列多重比對(duì)Fig.3 Comparison pf Dmrt1 protein alignment of in G.punctata and other species

表2 斑鱾Dmrt1與其他物種的同源性Tab.2 Homology between Dmrt1 in G.punctata and other species
在斑鱾和其他脊椎動(dòng)物的Dmrt1蛋白序列系統(tǒng)進(jìn)化樹(shù)中,斑鱾與黃金鱸、白梭吻鱸等鱸形目魚(yú)類(lèi)聚為一支,與斑馬魚(yú)、人類(lèi)和小鼠等分離,這與傳統(tǒng)分類(lèi)地位一致(圖4)。
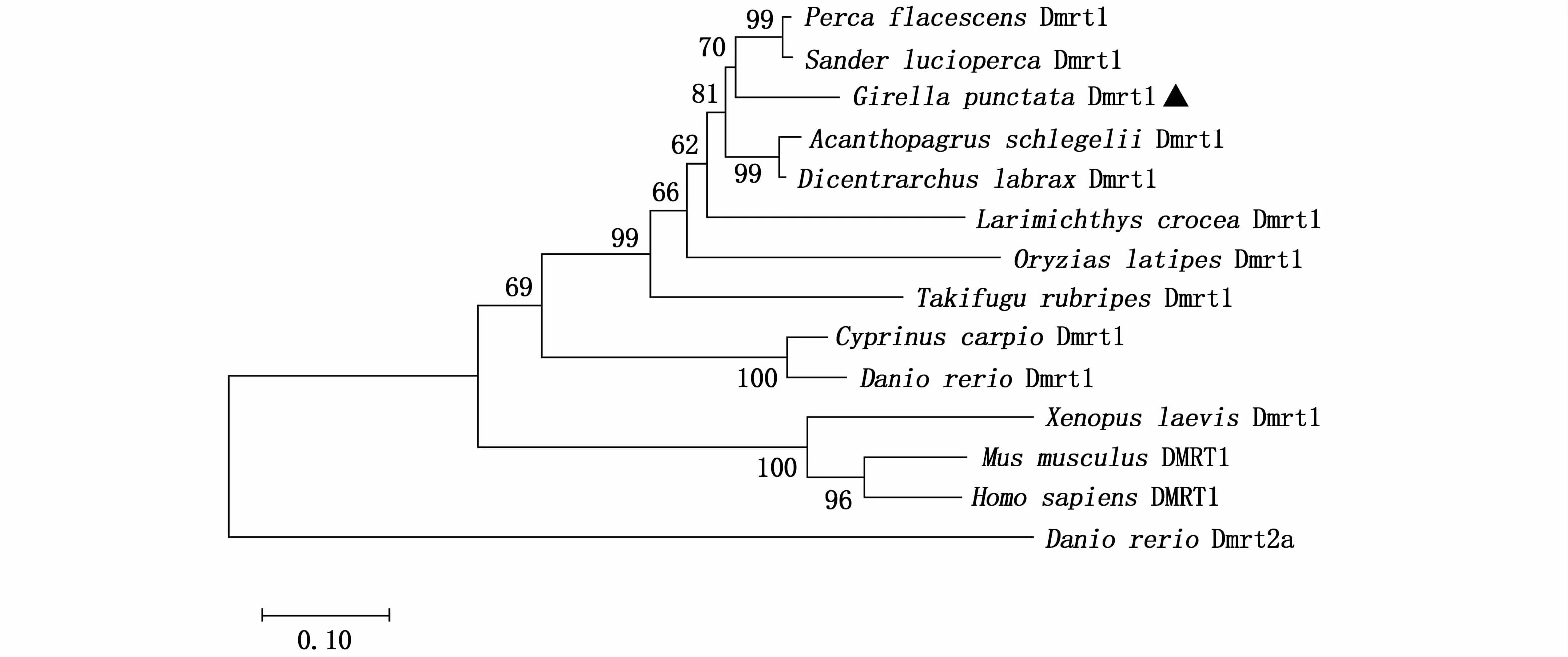
圖4 不同脊椎動(dòng)物的Dmrt1系統(tǒng)發(fā)生分析Fig.4 Phylogenetic analysis of Dmrt1 in vertebrates
2.3 斑鱾雌雄性腺的組織學(xué)觀察
雌雄性腺組織切片HE染色結(jié)果表明,該時(shí)期斑鱾精巢均處于III期,精原細(xì)胞呈近圓形或橢圓形,細(xì)胞中央有一大圓核,核透亮,核仁被蘇木精染成深藍(lán)色,胞質(zhì)被伊紅染成淡紅色;精母細(xì)胞由精原細(xì)胞增殖分裂產(chǎn)生,數(shù)量較多,染色程度較精原細(xì)胞深。斑鱾卵巢均處于II期,卵母細(xì)胞的形狀不規(guī)則,呈近圓形、多邊形或梨形等,細(xì)胞核偏位,占卵母細(xì)胞的比例較大(圖5)。
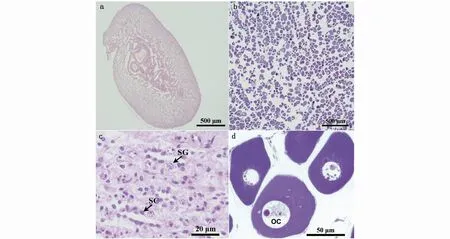
圖5 斑鱾雌雄性腺的組織切片觀察Fig.5 Histological observation of male and female gonads of G.punctata
2.4 dmrt1在斑鱾成體雌雄性腺的表達(dá)
qRT-PCR數(shù)據(jù)分析顯示,dmrt1在II期卵巢有微弱表達(dá),在III期精巢中有較高表達(dá),雄性的表達(dá)水平顯著高于雌性(P<0.05)(圖6)。
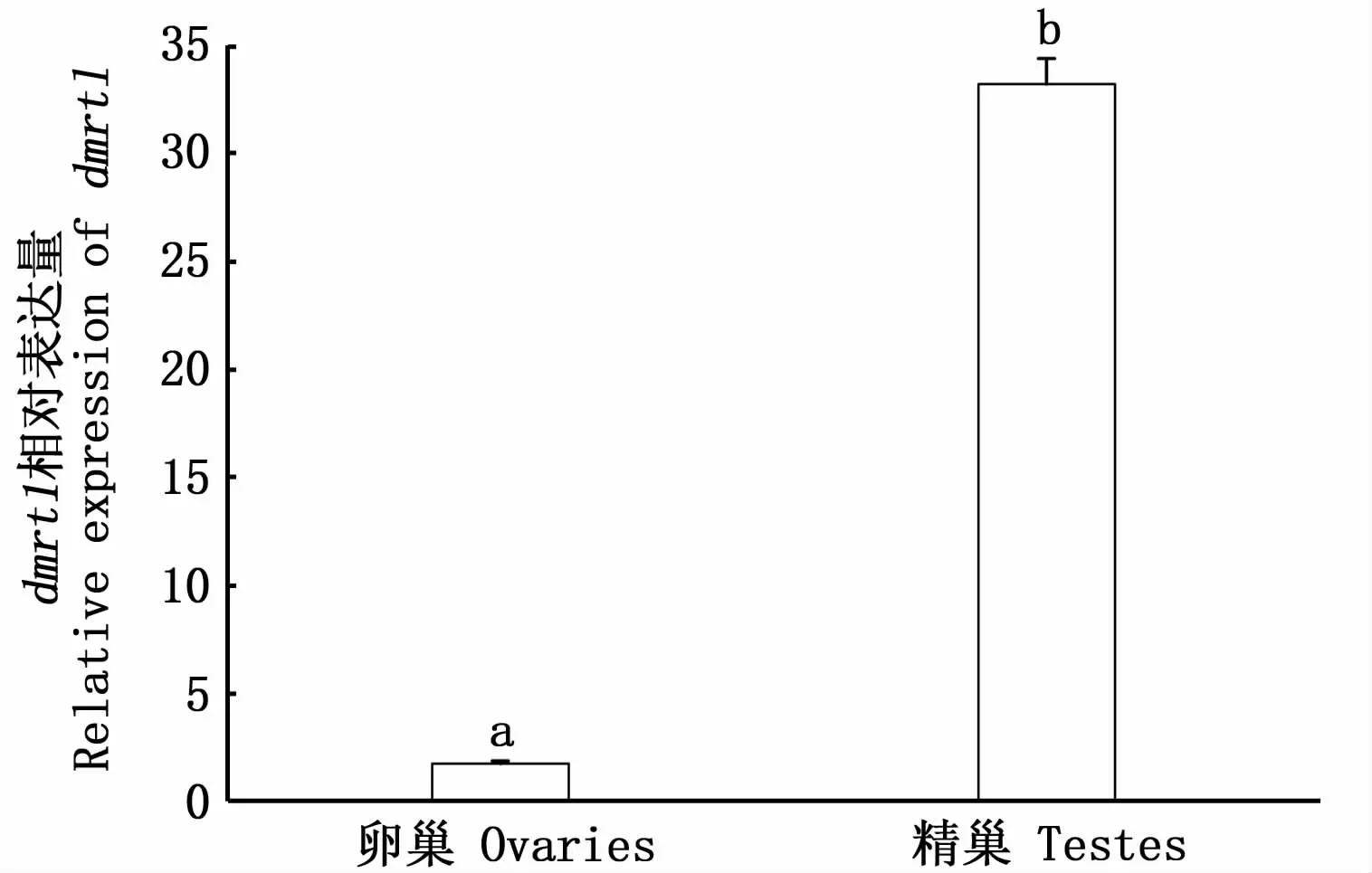
圖6 dmrt1在斑鱾雌雄性腺的相對(duì)表達(dá)Fig.6 Relative expression of dmrt1 in male and female gonads of G.punctata
2.5 斑鱾胚胎發(fā)育觀察
斑鱾受精卵屬端黃卵,為無(wú)色透明的圓球形,受精卵直徑為(973.92±21.46)μm(n=36),具有1個(gè)油球,油球直徑為(221.55±11.68)μm。斑鱾受精卵在溫度為(26.0±0.5)℃、鹽度為31的海水中培育,從受精到超過(guò)50%的仔魚(yú)孵出,整個(gè)胚胎發(fā)育過(guò)程歷時(shí)24 h 36 min,胚胎發(fā)育各個(gè)階段及主要發(fā)育特征見(jiàn)表3和圖7。
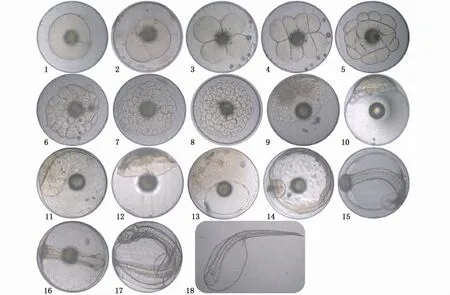
圖7 斑鱾胚胎發(fā)育Fig.7 Embryonic development of G.punctata
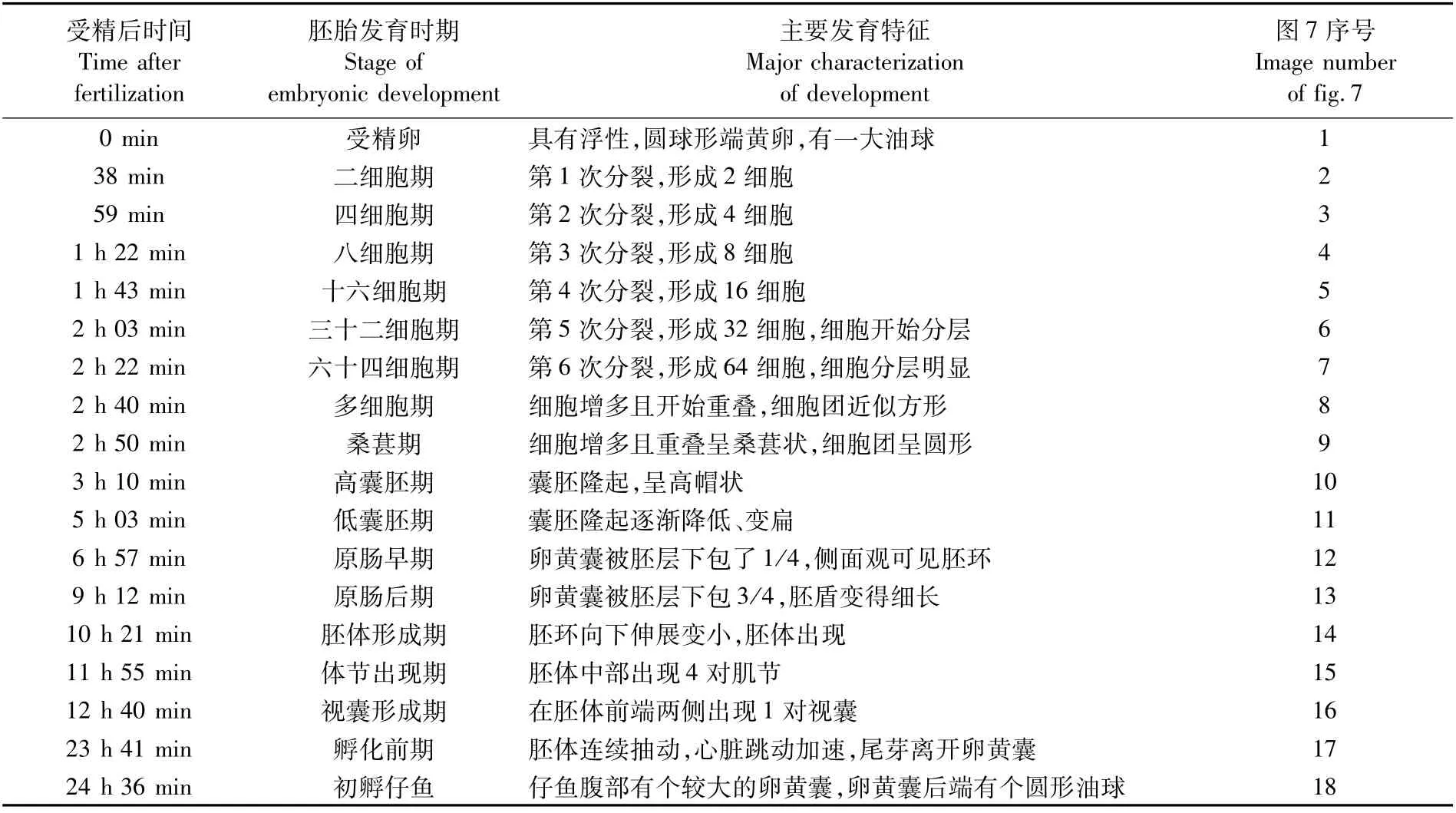
表3 斑鱾胚胎發(fā)育時(shí)序Tab.3 Embryonic development of G.punctata
2.6 dmrt1在斑鱾胚胎不同發(fā)育時(shí)期的表達(dá)
qRT-PCR數(shù)據(jù)分析顯示,dmrt1表達(dá)水平在斑鱾胚胎發(fā)育過(guò)程中呈先上升后降低的趨勢(shì),在桑葚期表達(dá)水平最高,囊胚期開(kāi)始降低,原腸期后表達(dá)水平急劇下降,至初孵期幾乎不表達(dá)(圖8)。
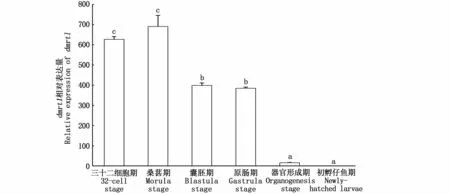
圖8 dmrt1在斑鱾胚胎不同發(fā)育時(shí)期的相對(duì)表達(dá)Fig.8 Relative expression of dmrt1 in embryo at different development stages in G.punctata
3 討論
本研究克隆的斑鱾dmrt1基因的ORF全長(zhǎng)為894 bp,編碼297個(gè)氨基酸。運(yùn)用生物信息學(xué)方法預(yù)測(cè)分析斑鱾Dmrt1蛋白的結(jié)構(gòu)與性質(zhì),結(jié)果顯示,該蛋白無(wú)N端信號(hào)肽和跨膜區(qū)結(jié)構(gòu),位于細(xì)胞核,屬不穩(wěn)定性親水蛋白,預(yù)測(cè)結(jié)果與其結(jié)合特定DNA序列調(diào)控下游基因表達(dá)的特性相符。斑鱾Dmrt1蛋白的蛋白結(jié)構(gòu)、細(xì)胞定位、穩(wěn)定性和疏水性等多項(xiàng)預(yù)測(cè)結(jié)果與奧里亞羅非魚(yú)(Oreochromis aureus)、胡 子 鲇(Clarias fuscus)Dmrt1的預(yù)測(cè)結(jié)果相似[20-21]。通過(guò)與其他脊椎動(dòng)物Dmrt1多重蛋白比對(duì)發(fā)現(xiàn),斑鱾Dmrt1存在較為保守的DM結(jié)構(gòu)域序列,這預(yù)示著斑鱾Dmrt1具有與其他脊椎動(dòng)物Dmrt1相似的功能。而斑鱾Dmrt1除DM結(jié)構(gòu)域外的序列與其他脊椎動(dòng)物的同源性差異較大,表明Dmrt1蛋白在不同進(jìn)化地位的物種間仍然具有一定差異。
dmrt1基因是dmrt基因家族中研究最廣泛的性別調(diào)控相關(guān)基因,廣泛存在于各種魚(yú)類(lèi)中,但在不同物種間存在時(shí)序和組織表達(dá)差異。在魚(yú)類(lèi)dmrt1基因性別相關(guān)研究中發(fā)現(xiàn),dmrt1是具有性別二態(tài)性的雄性高表達(dá)基因,即其在精巢中的表達(dá)量要遠(yuǎn)遠(yuǎn)高于在卵巢及其他組織中的表達(dá)量,有些魚(yú)類(lèi)的dmrt1基因甚至只在精巢中特異性表達(dá)。如斑馬魚(yú)[22]性腺切片原位雜交表明,dmrt1在精巢和卵巢發(fā)育的生殖細(xì)胞中均有表達(dá),且Northern印跡雜交結(jié)果顯示,其在精巢的表達(dá)遠(yuǎn)高于卵巢;虹鱒[10]Northern印跡雜交結(jié)果顯示,dmrt1在其整個(gè)精子發(fā)生過(guò)程中持續(xù)表達(dá),且通過(guò)RT-qPCR技術(shù)檢測(cè)到dmrt1在其卵巢中的微弱表達(dá)。銀漢魚(yú)(Odontesthes bonariensis)[23]、紅 鰭 東 方 鲀(Takifugu rubripes)[24]和 香 魚(yú)(Plecoglossus altivelis)[25]等魚(yú)類(lèi)dmrt1研究結(jié)果與斑馬魚(yú)類(lèi)似。此外,在成體青鳉[26]、尼羅羅非魚(yú)[27]和黑鯛(Acanthopagrus schlegeli)[28]等魚(yú)類(lèi)中,dmrt1僅在精巢中特異性表達(dá),在其他組織中不表達(dá)。而在本實(shí)驗(yàn)中,斑鱾dmrt1在雌雄性腺中的表達(dá)模式與前者一致,其在雄性III期精巢中有較高表達(dá),在II期卵巢中微弱表達(dá),這預(yù)示著其可能不僅與精巢發(fā)育有關(guān),還可能在卵巢發(fā)育過(guò)程中起作用。
胚胎發(fā)育階段,在斜帶石斑魚(yú)(Epinephelus coioides)[29]、稀有鮈鯽(Gobiocypris rarus)[30]、黃顙魚(yú)(Pelteobagrus fulvidraco)[31]、香魚(yú)[25]和河川沙塘鱧(Odontobutis potamophila)[32]等魚(yú)類(lèi)研究中發(fā)現(xiàn),dmrt1基因從受精卵裂到出膜孵化整個(gè)胚胎發(fā)育過(guò)程均有表達(dá),表達(dá)水平隨著胚胎發(fā)育呈現(xiàn)出先升后降的趨勢(shì),但不同物種到達(dá)最高表達(dá)的時(shí)期有所差異,如斜帶石斑魚(yú)和稀有鮈鯽的dmrt1的表達(dá)水平在囊胚期達(dá)到最高值,而黃顙魚(yú)、黃河鯉(Cyprinus carpio)[33]、香魚(yú)和河川沙塘鱧在原腸期達(dá)到峰值。此外,有些魚(yú)類(lèi)在胚胎發(fā)育階段 dmrt1 基因不表達(dá),如大口黑鱸(Micropterus salmoides)[34],在孵化后40 d才開(kāi)始出現(xiàn)dmrt1的表達(dá)。在本研究中,斑鱾dmrt1在胚胎發(fā)育的卵裂期到原腸期有較高表達(dá),而原始生殖細(xì)胞(PGCs)最早發(fā)現(xiàn)于原腸胚時(shí)期,推測(cè)dmrt1在胚胎發(fā)育前期的高表達(dá)與原始生殖細(xì)胞的形成有關(guān)。此外dmrt1在斑鱾初孵仔魚(yú)期幾乎不表達(dá),這與已有研究結(jié)果不同,有可能是一種新的表達(dá)模式,仍有待進(jìn)一步研究。
綜上所述,本研究克隆的斑鱾dmrt1基因的序列,其所編碼的蛋白具有典型的Dmrt1結(jié)構(gòu)特征,在進(jìn)化上與鱸形目魚(yú)類(lèi)的Dmrt1同源性高。在雌雄性腺和胚胎發(fā)育過(guò)程中的表達(dá)模式表明,斑鱾dmrt1基因可能參與了性腺分化和早期胚胎發(fā)育。
