急冷與無水雙重脅迫下南美白對蝦存活變化及防御系統響應規律
2022-07-29 02:40:42吳嘉鑫徐德峰孫力軍秦小明范秀萍
食品科學 2022年13期
關鍵詞:氧化應激
陳 晗,吳嘉鑫,徐德峰,孫力軍,秦小明,范秀萍
(廣東海洋大學食品科技學院,廣東 湛江 524088)
南美白對蝦對外界環境脅迫反應較為強烈,捕撈和運輸過程中的不利因素常會造成其大量應激性傷殘和死亡,因此降低應激反應從而提高存活質量已成為南美白對蝦保活流通的關注熱點。傳統的有水保活運輸因需要大量水而成本高昂,運輸途中的對蝦排泄物嚴重影響機體存活狀況。近年來快速發展的無水保活運輸由于消除了運輸過程對水的需求,且裝載量更大、綜合成本大幅縮減,因而受到業界的廣泛關注。但對蝦生長環境溫度通常在20 ℃以上,捕撈后短時間快速冷應激和運輸過程中長時間的無水暴露會影響對蝦生理代謝和存活質量。氧化應激和免疫防御失衡通常是生物細胞在應答極端環境脅迫時的基本表現,相關研究表明,魚、貝類在無水保活過程中不利環境因子可觸發其氧化應激,使其先天免疫系統失衡,甚至造成組織損傷。研究發現,12 ℃低溫和空氣暴露3~5 min可顯著改變南美白對蝦的抗氧化防御系統,而關于產業運輸實際中急冷并長時間無水脅迫如何影響南美白對蝦氧化和免疫系統鮮見報道。因此,有必要結合產業實際探究急冷及無水雙重脅迫誘導南美白對蝦死亡的氧化和免疫防御失衡機制。
此外,肝胰腺是對蝦生理代謝的重要器官之一,也是響應外界脅迫的關鍵器官。鑒于肝胰腺組織在對蝦機體響應環境脅迫中的作用,可將其作為無水保活流通過程中南美白對蝦冰溫損傷效應評價的靶組織。無水保活流通過程中的低溫和空氣暴露聯合應激對南美白對蝦肝胰腺抗氧化和免疫系統的影響尚不清楚。因此,本研究通過模擬南美白對蝦無水保活運輸條件,考察急冷及無水脅迫下對蝦肝胰腺氧化應激和非特異性免疫指標變化,解析雙重脅迫影響機體存活的生理防御基礎,以期為優化對蝦保活運輸條件提供參考。
1 材料與方法
1.1 材料與試劑
10 kg鮮活南美白對蝦()購自廣東省湛江市霞山水產品批發市場,質量(16.23±2.12)g、體長(7.36±1.42)cm,市場出售的對蝦均在凌晨4點左右起塘后快速放入盛有新鮮海水的運輸容器內,由裝有控溫和持續供氧裝置的配送車輛在4 h內運輸至批發市場,剔除死亡個體后放于盛有新鮮海水的水族缸或鐵盆內持續泵入新鮮空氣。市場選購實驗用蝦時,用容積20 L的聚乙烯袋將10 kg對蝦均勻分成5 袋,袋中注入新鮮海水10 L,用氧泵充分打氧后扎緊袋口2 h內運送到廣東海洋大學水產品保活實驗室,之后迅速將對蝦放于溫度25 ℃、鹽度35‰的約1 m新鮮海水中暫養6 h,以緩解和消除捕撈和運輸操作時蝦的應激反應,然后剔除死亡個體,挑選無機械傷、活動正常、狀態良好的對蝦進行實驗。
酚氧化酶(phenol oxidase,PO)試劑盒 江蘇酶免實業有限公司;活性氧(reactive oxygen species,ROS)試劑盒、丙二醛(malondialdehyde,MDA)試劑盒、超氧化物歧化酶(superoxide dismutase,SOD)試劑盒、過氧化氫酶(catalase,CAT)試劑盒、谷胱甘肽過氧化物酶(glutathion peroxidase,GSH-Px)試劑盒、過氧化物酶(peroxidase,POD)試劑盒、酸性磷酸酶(acid phosphatase,ACP)試劑盒、堿性磷酸酶(alkaline phosphatase,AKP)試劑盒 南京建成生物工程研究所。
1.2 儀器與設備
S-20氧泵 中國浙江寧波塞爾電氣公司;HX-1508低溫冷卻循環泵 江蘇天翔儀器有限公司;25 cm×20 cm塑料自封袋 天津永業塑料制品有限公司;塑料周轉箱(490 mm×345 mm×285 mm)東莞市茶山中距塑膠卡板廠;5810R臺式高速冷凍離心機德國艾本德有限公司;Varioskan全自動酶標儀 賽默飛世爾科技(中國)有限公司。80i顯微鏡 日本尼康公司;RM2245半自動轉輪切片機 德國徠卡儀器有限公司;JEM-2100F場發射透射電子顯微鏡 日本電子株式會社;BCD-166TNA冰箱 青島海爾股份有限公司;DHG-9053A電熱鼓風干燥箱 上海一恒科學儀器有限公司;BSA224S電子分析天平 賽多利斯科學儀器(北京)有限公司。
1.3 方法
1.3.1 南美白對蝦脅迫處理與存活率測定
參照徐德峰等的方法,取6個塑料周轉箱(490 mm×345 mm×285 mm),分別注入20 L新鮮海水,并連續編號,將暫養后挑選的300 尾對蝦隨機分成6 組,每組50 尾,1號箱中50 尾對蝦記為正常對照組(NC組),自然室溫觀察和取樣測定相關指標。2~6號箱均先用低溫冷卻循環泵將室溫新鮮海水降至12 ℃,然后將50 尾對蝦轉移到該冷水箱進行急性低溫30 min誘導休眠,2號箱中的對蝦不取出(放置12 h),記為單獨冷脅迫組(AC組)。剩余3~6號箱中的對蝦經12 ℃冷休眠30 min后取出,隨機放入塑料自封袋,每袋10 尾,每組5 袋,共4 組,自封袋充分充入純氧氣后封口,將充氧后的4 組對蝦貯存在12 ℃的層析柜中,模擬對蝦無水運輸低溫環境,3~5號箱中對蝦分別經歷冷休眠和無水脅迫處理3、6、9 h時的聯合脅迫,相應記為AC+WD3 h、AC+WD6 h、AC+WD9 h組。6號箱對蝦經歷急冷和9 h無水聯合脅迫,之后于自然室溫海水中復蘇2 h,記為AC+WD9 h+R組。實驗過程中,嚴格控制對蝦個體差異,取各組大小、顏色、狀態基本一致的具有代表性的對蝦15 尾,測定集體防御系統相關指標。在統計存活率時將雙重脅迫處理組均置于自然室溫海水中復蘇2 h。
1.3.2 樣品采集與生化指標測定
樣品制備參照李曉麗等的實驗方法,將各組對蝦置于冰盤上迅速解剖,由頭胸部取出肝胰腺,用預冷至4 ℃的無菌生理鹽水沖洗掉表面血跡并用吸水紙吸干表面水分,液氮速凍后放于-80 ℃保存備用。指標測定時取出冷凍肝胰腺在4 ℃解凍30 min,再次用4 ℃的無菌生理鹽水沖洗組織表面并吸干水分,切取2 g左右組織樣品,按1∶9(/)的比例加入4 ℃預冷后的無菌生理鹽水,冰浴條件下充分研磨6~8 min,之后3 000×冷凍離心10 min,取上清液測定相關生化指標。ROS水平(以熒光強度表征)、MDA含量以及SOD、CAT、GSH-Px、PO、POD、ACP、AKP活力測定均嚴格按照試劑盒說明書進行,結果均以蛋白質量計。
1.3.3 肝胰腺組織樣品制備與觀察
取各組對蝦肝胰腺組織,超凈工作臺中的操作參照徐德峰等的實驗方法,用手術解剖刀配合剃須刀片,從肝胰腺正中縱切剖開,然后進一步在縱切面的正中橫切,在橫切面部位切取5 mm×5 mm×5 mm組織塊,在Bouin’s固定液中進行組織固定,固定結束后梯度乙醇脫水,組織透明后石蠟包埋,石蠟切片厚度6 μm,常規蘇木精-伊紅染色程序染色后進行光學顯微鏡觀察并拍照。
1.4 數據統計與分析
實驗所得結果均用平均值±標準差表示(=6),采用SPSS 19.0軟件中的最小顯著差異法進行差異顯著性分析,<0.05表示差異顯著。采用Origin Pro 8.5軟件繪圖。
2 結果與分析
2.1 雙重脅迫對南美白對蝦存活率的影響
由圖1可以看出,NC組和AC組在12 h的實驗周期內均全部存活,存活率均為100%,雙重脅迫下無水脅迫時間控制在6 h內均可全部復蘇,而無水脅迫9 h時部分對蝦不能復蘇而沉在水底,存活率為92.13%,表明對蝦存活率在無水脅迫9 h時開始發生變化,提示機體內部臟器組織可能因強烈生理應激而發生不可逆損傷。
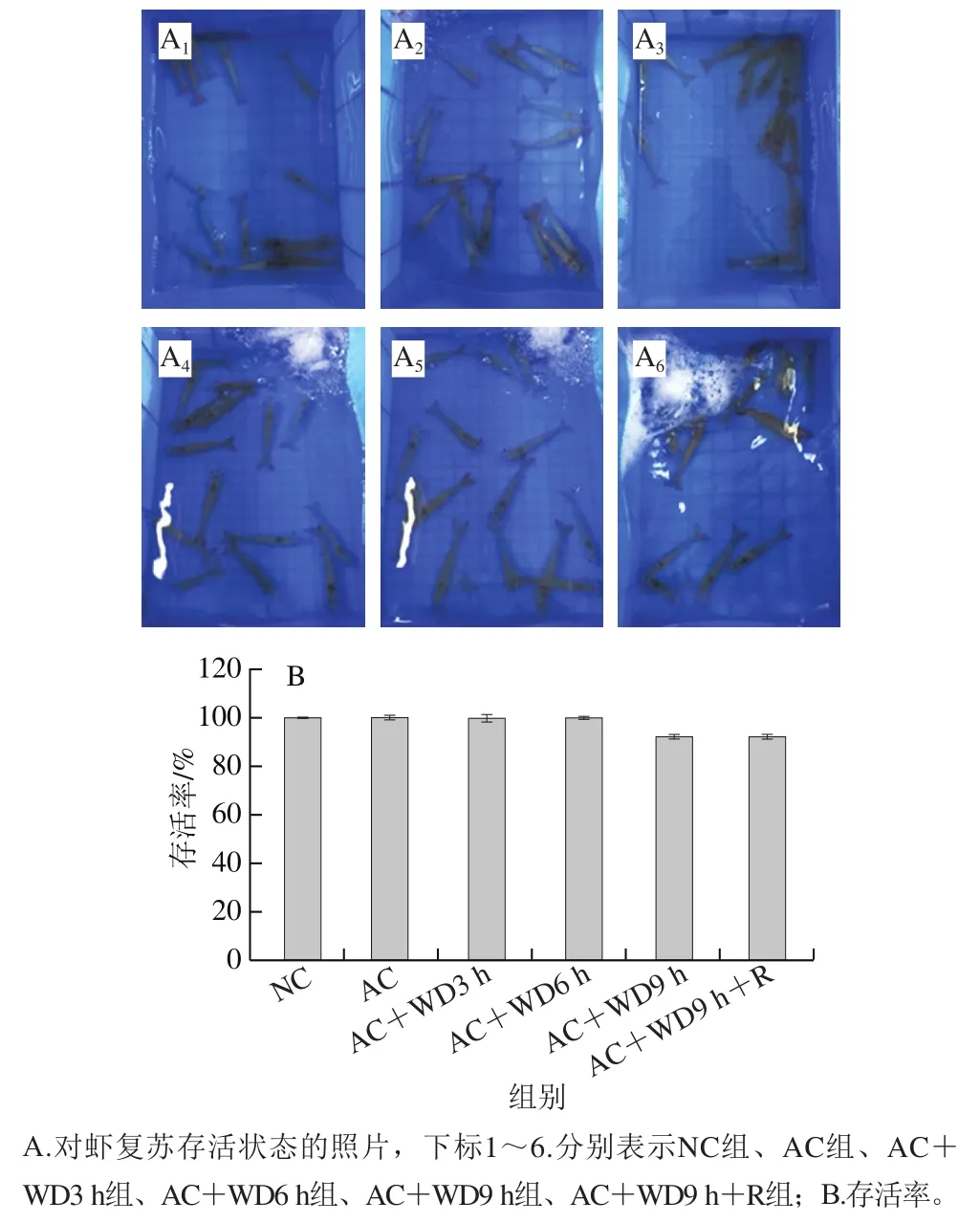
圖1 雙重脅迫對南美白對蝦存活情況的影響Fig. 1 Effect of combined stress on the survival rate of L. vannamei
2.2 雙重脅迫對南美白對蝦肝胰腺氧化應激損傷的影響
由圖2A可知,與NC組相比,AC組對蝦肝胰腺ROS生成量顯著增加(<0.05),原因可能是對蝦為應對急性低溫脅迫需要迅速產生能量,因而呼吸速率增加,使ROS的生成速率大于清除速率。但隨后無水脅迫下ROS生成量顯著降低(<0.05),雙重脅迫下無水脅迫9 h組在自然室溫海水中復蘇后ROS再次恢復至NC組水平。
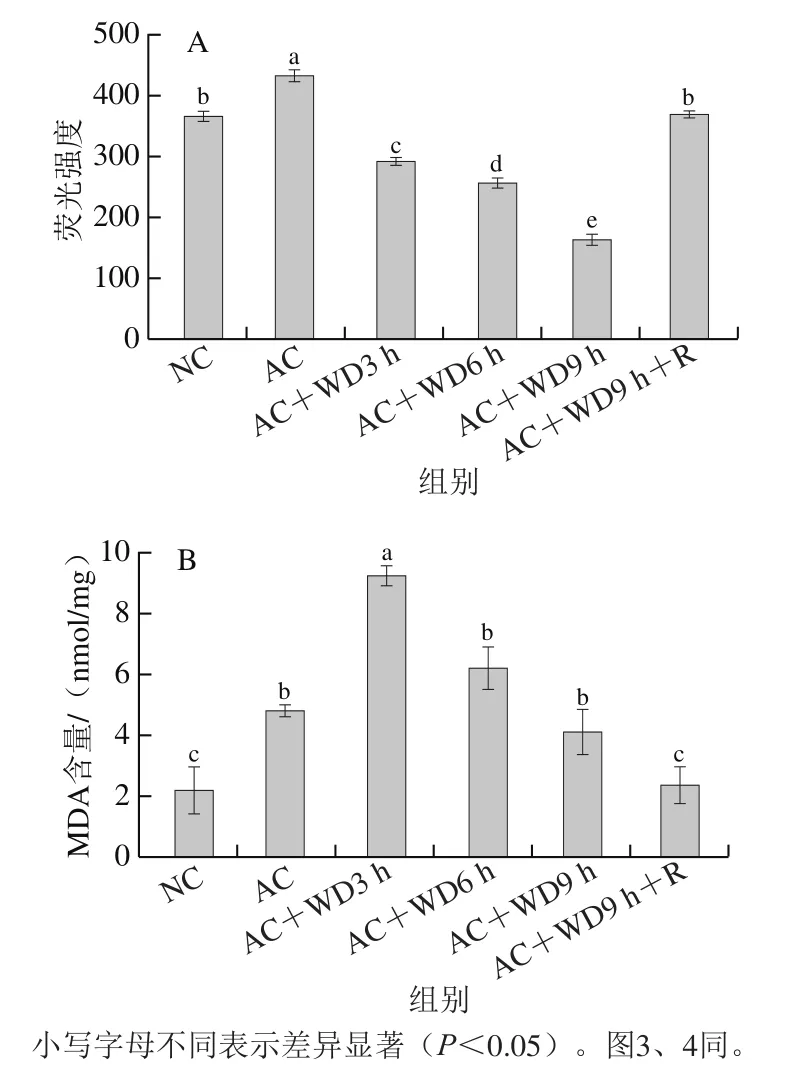
圖2 雙重脅迫對南美白對蝦肝胰腺ROS水平(A)和MDA含量(B)的影響Fig. 2 Effect of combined stress on ROS level (A) and MDA content (B)in hepatopancreas of L. vannamei
由圖2B可知,與NC組相比,AC組南美白對蝦肝胰腺MDA含量顯著增加(<0.05),且在進一步的無水脅迫過程中,MDA含量呈先升后降的趨勢,但均顯著高于NC組(<0.05),其中MDA含量在雙重脅迫3 h時達到9.23 nmol/mg,之后MDA含量逐漸下降,自然室溫海水中復蘇后(AC+WD9 h+R組)可基本恢復到初始水平。整體來看,在無水活運雙重脅迫過程中,MDA含量相比ROS水平高峰出現的時間晚,表明體內ROS積累后造成MDA含量隨之增加,無水脅迫后MDA含量下降可能緣于對蝦抗氧化系統被激活,加速清除過多MDA以維持體內氧化和抗氧化系統平衡,提示機體抗氧化防御系統基本可以呈時間依賴性消除脂質過氧化。
2.3 雙重脅迫對南美白對蝦肝胰腺主要抗氧化酶活力的影響
抗氧化系統是一個復雜的適應性系統,用于維持細胞功能和防止氧化應激,SOD、CAT和GSH-Px是甲殼類動物抗氧化系統中的關鍵酶。由圖3A可以看出,AC組SOD活力(2.22 U/mg)相比NC組(1.69 U/mg)顯著上升(<0.05),之后隨著無水脅迫時間的延長略微上升,并在雙重脅迫3~6 h之間基本保持穩定,但在9 h時再次顯著升高至2.82 U/mg(<0.05),AC+WD9 h+R組在自然室溫海水中復蘇后SOD活力有所下降,但未恢復到初始水平,表明脅迫環境下機體可通過增加南美白對蝦肝胰腺SOD活力以減輕氧化應激損傷。
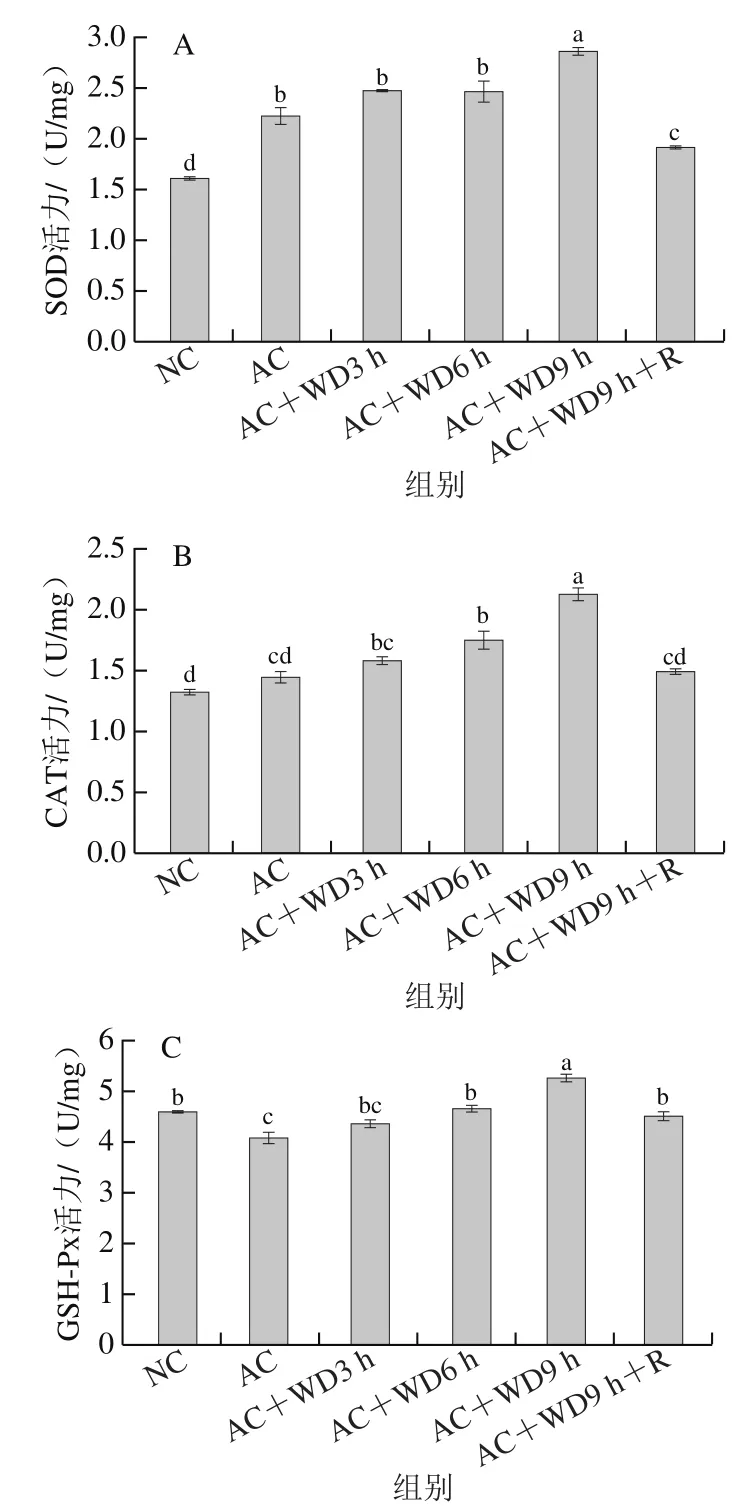
圖3 雙重脅迫對南美白對蝦肝胰腺SOD(A)、CAT(B)、GSH-Px(C)活力的影響Fig. 3 Effect of combined stress on the activities of SOD (A), CAT (B)and GSH-Px (C) in hepatopancreas of L. vannamei
CAT和GSH-Px能夠有效地清除過氧化氫,以維持動物體內的氧化應激與抗氧化系統平衡。由圖3B可以看出,與NC組相比,急冷后對蝦肝胰腺CAT活力有所增加,但不顯著(>0.05)。隨后在雙重脅迫下CAT活力隨無水脅迫時間的延長而增加,且均較NC組顯著提升,9 h時達到2.12 U/mg,AC+WD9 h+R組在自然室溫海水中復蘇后CAT活力與NC組無顯著差異,提示CAT對于對蝦抵抗無水暴露引起的不良影響有積極作用。由圖3C可以看出,急冷脅迫后對蝦肝胰腺GSH-Px活力較NC組顯著降低(<0.05),之后隨著無水脅迫時間的延長,GSHPx活力逐漸上升,并在9 h時達到5.26 U/mg,AC+WD9 h+R組在自然室溫海水中復蘇后可基本恢復到NC組水平,提示對蝦可通過提升GSH-Px活性以抵抗氧化損傷。
2.4 雙重脅迫對肝胰腺主要非特異性免疫酶活力的影響
PO是一種金屬蛋白酶,動物體內酚氧化酶原激活系統被異物激活后,PO可將酚催化成黑色素,并將一些病原體殺死,在對蝦非特異性免疫中起重要作用。由圖4A可以看出,對蝦經歷急冷脅迫后,肝胰腺PO活力基本沒有變化,但經雙重脅迫后均較NC組顯著增加(<0.05),其中3 h后PO活力最高,之后逐漸降低,直到復蘇后(AC+WD9 h+R組)仍未下降到初始水平,推測在雙重脅迫前期(3 h之前)可刺激機體表達PO,之后可能由于免疫系統受到不可逆損傷而使PO表達量下降。
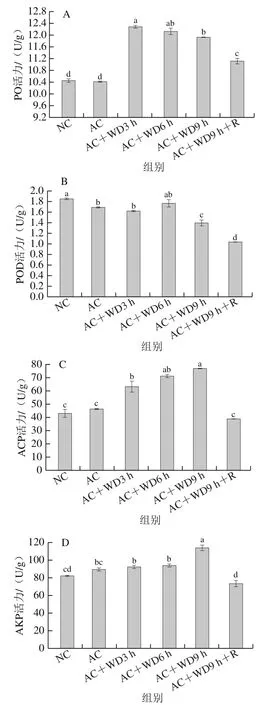
圖4 雙重脅迫對南美白對蝦肝胰腺PO(A)、POD(B)、ACP(C)、AKP(D)活力的影響Fig. 4 Effect of combined stress on the activities of PO (A), POD (B),ACP (C) and AKP (D) in hepatopancreas of L. vannamei
POD是與對蝦體液免疫相關的酶,可以將超氧陰離子自由基轉化為無毒的水和氧,以避免細胞損傷,POD水平可間接反映機體免疫水平。由圖4B可以看出,除了雙重脅迫6 h時POD活力略低于NC組之外(>0.05),其他組POD活力均顯著低于NC組(<0.05),且隨脅迫時間的延長整體呈降低趨勢;復蘇后(AC+WD9 h+R組)POD活力也沒有恢復,仍顯著低于NC組(<0.05)。ACP和AKP直接參與磷酸基團的催化、代謝和水解,并能與其他酶類形成水解酶系消除異物,在對蝦免疫防御系統起關鍵作用,可作為生物抵抗環境脅迫評價的標記物。由圖4C、D可以看出,ACP和AKP活力均隨無水脅迫時間的延長而逐漸增加,在9 h分別達到76.76 U/g和114.13 U/g,顯著高于NC組(<0.05)。急冷脅迫后,ACP和AKP活力相比NC組有所上升,但變化不顯著(>0.05),復蘇后(AC+WD9 h+R組)ACP和AKP活力均與初始水平無顯著差異。
2.5 重脅迫對肝胰腺組織結構的損傷效應
環境脅迫下的機體應激強度不僅可通過生理指標進行考察,還可通過組織病理改變進行評價,雙重脅迫對南美白對蝦肝胰腺組織結構的損傷效應見圖5。
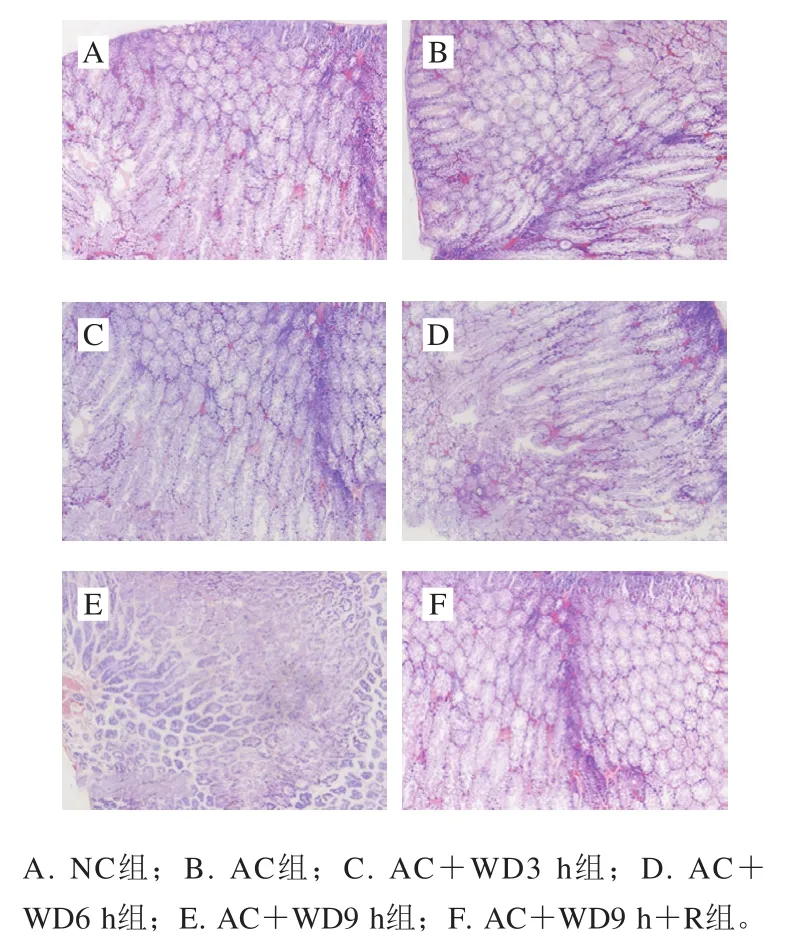
圖5 雙重脅迫對南美白對蝦肝胰腺組織結構的影響(40×)Fig. 5 Effect of combined stress on histopathology of hepatopancreas of L. vannamei (40 ×)
由圖5可以看出,NC組肝胰腺組織結構清晰、成分完整、肝小管排列規則、細胞形態正常,AC組經急冷脅迫后細胞形態整體無明顯變化,短期雙重脅迫也未見明顯結構變化(AC+WD3 h組),但雙重脅迫6 h和9 h時可見肝小管有輕微的破裂和內壁細胞脫落,復蘇后(AC+WD9 h+R組)組織損傷有所減輕,表明長時間的雙重脅迫對肝胰腺細胞結構造成了明顯的損傷,而復蘇后可在一定程度上修復細胞損傷。
3 討 論
ROS的來源主要是線粒體,細胞呼吸代謝產生能量的過程中,約0.1%~0.2%的O可轉化為ROS,引起細胞代謝的氧化應激。氧化應激被定義為ROS產生和消除之間的不平衡,可引起典型的脂質過氧化反應,損傷細胞膜系統,其標志性表現就是MDA水平的顯著增加。溫度脅迫會導致細胞呼吸強度大幅增加,產生大量ROS,使細胞氧化應激增強,從而對機體細胞造成一系列生理和病理損傷,為緩解和消除氧化應激,機體必然會調動自身抗氧化防御系統來消減ROS和MDA。研究表明,在急性冷應激或熱應激后,南美白對蝦抗氧化酶基因表達均上調,由此推測環境突然改變可誘導機體抗氧化酶系激活,加速分解體內累積的ROS,本研究中雙重脅迫9 h復蘇后ROS水平恢復到初始水平,其原因可能在于常溫水體中機體細胞代謝逐漸恢復至正常水平,ROS因機體代謝活動的應激性增強而積累,因此ROS水平再次上升。研究發現,空氣暴露時間的長短不僅影響南美白對蝦的存活率,同時會導致肌肉和肝胰腺中MDA含量有所升高,且空氣暴露3 h可使日本囊對蝦肝胰腺中的MDA含量顯著增加,放回水中恢復至正常值,此研究與本實驗結果相符。
SOD是一種胞質酶,催化兩分子ROS生成一分子過氧化氫和一分子氧氣,在自衛系統中發揮重要作用。南美白對蝦經歷低鹽和亞硝酸鹽聯合脅迫24 h和48 h后,肝胰腺SOD活力顯著上升,此外,南美白對蝦遭受持續應激時SOD活力上升,可見不良環境脅迫會誘導南美白對蝦體內SOD活力增加。南美白對蝦在氨氮或硫化物脅迫的過程中,體內SOD和CAT活力顯著上調,與本研究結果相符。將南美白對蝦從鹽度30‰水體轉移到鹽度15‰的水體時GSH-Px活力上升,提示對蝦可通過提升GSH-Px活力以抵抗氧化損傷。另外,劉曉云等的研究中中國對蝦在持續環境應激下PO活力的變化與本研究結果相似,表明南美白對蝦免疫能力總是隨其自身狀態和外界環境的改變而發生改變。徐子涵研究發現,無水低溫脅迫下南美白對蝦血淋巴和肝胰腺POD活力在保活6 h內較高,隨后POD活力顯著降低,且基因表達量也顯著下降。以上研究結果提示,長時間低溫和無水脅迫抑制了對蝦免疫功能。
吳萌探討了低溫對養殖凡納濱對蝦呼吸性能及抗氧化能力的影響,表明在水溫由20 ℃降至10 ℃的過程中,肝胰腺MDA含量先降低后升高,總抗氧化能力呈下降趨勢,其中抗氧化酶CAT活力呈上升趨勢,并在溫度降至10 ℃時最高;此外,鰓組織結構損傷加重,角質層及上皮細胞出現壞死脫落,血細胞出現破裂。賈旭穎模擬環境溫度突變系統研究了溫度脅迫對凡納濱對蝦存活和生理代謝的影響,發現當溫度由22 ℃驟降到12 ℃時,對蝦存活率隨時間的延長而下降,且體內血細胞數量顯著下降,肝胰腺MDA含量在3~6 h增加,隨后逐漸恢復到溫度突變前的水平,PO、活性氮和SOD水平均呈先升高后下降的變化趨勢。在本研究中,急冷后ROS水平較NC組顯著增加,而急冷與無水聯合脅迫下ROS水平下降;MDA水平在急冷和雙重脅迫前3 h均較NC組顯著增加,而后顯著降低,可能在于酶促抗氧化系統被激活,誘導產生的抗氧化酶SOD和CAT快速清除體內過多的ROS,使脅迫中后期機體MDA水平相應降低。同時,伴隨著抗氧化系統的激活,機體免疫防御系統被激活,體內典型非特異性免疫相關酶除POD外,PO、ACP和AKP活力均有不同程度的提升。長時間的氧化應激必然造成不同程度的細胞損傷,AC+WD9 h組中肝小管的輕微破裂和內壁細胞脫落解釋了為何此時對蝦存活率會突然下降。因此可以推測,雙重脅迫觸發了細胞氧化應激損傷,激活了抗氧化和非特異性免疫系統,在6~8 h的時間內機體可以適應上述環境改變,可一旦超過9 h,細胞因累積性氧化損傷而造成組織損傷,進而導致存活率降低,研究結果證實了氧化和免疫系統參與了無水保活流通過程中急冷及無水聯合脅迫誘導的對蝦死亡,提示后續研究可進一步考察不同冰溫參數對存活率影響的氧化和免疫響應生化基礎,進而靶向優化保活流通參數。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
