井岡山地區蓮魚復合種養模式效益評價
2022-08-04 15:15:40米湘媛張朝碩張曉娟王海華郭小澤李思明趙大顯洪一江段明
水生生物學報 2022年7期
米湘媛 張朝碩 張曉娟 王海華 郭小澤 李思明趙大顯 洪一江 段明
(1.中國科學院水生生物研究所淡水生態與生物技術國家重點實驗室,武漢 430072;2.中國科學院大學,北京 100049;3.江西省水產科學研究所,南昌 330039;4.江西省農業科學院畜牧獸醫研究所,南昌 330200;5.南昌大學,南昌 330047)
漁農復合種養(Integrated Agriculture-Aquaculture,IAA)是指利用生物群落共生原理、系統內組分之間相互協調和促進及物質循環利用原理,通過產品多樣化來維持系統生產力和分散經濟風險的生產模式[1]。不僅能提高單位面積利潤[2],還能減少農業用水[3]、減少化肥和農藥的使用、減少作物病害[4]、降低溫室氣體的排放[5]、減輕養殖池塘中的富營養化程度[6]及增加農村就業人口等[7],被認為是以小農經濟為主的發展中國家的一種可持續農業模式[8]。稻田養魚是我國一種傳統的農漁復合種養模式,至今已經有兩千多年歷史[9],在此基礎上發展出來的稻-鱉[10]、稻-蝦[11,12]、稻-蛙[13]、稻-魚[14]和稻-鰍[15]等復合種養在我國得到了廣泛的生產實踐,在少數蓮藕資源豐富的地區,還因地制宜地開展了蓮-蝦[16]和蓮-鱉復合種養[17]。漁農復合種養還為稻田相對分散、海拔較高、水溫偏低、作物畝產量相對平原地區較低的山區提供了相比單一種植水稻更為經濟高效、生態友好的可持續發展生產模式,成為我國四川廣元[7]、貴州遵義[18]、浙江青田[4,19]和江西井岡山[17]等山區農業產業結構調整、穩糧增效、帶動當地農戶脫貧致富的有效手段。
浮游植物群落結構不僅能反映水體的營養狀態,還是衡量水體生態系統健康程度的重要指示生物[20]。浮游植物對水體狀況十分敏感,水體理化因子如營養鹽、溫度、pH和溶氧等都會對水體中的浮游植物產生影響,不同環境的主要影響因子各不相同且存在較大差異[21,22];而氮、磷長期以來被認為是影響浮游植物群落結構的重要環境因子[23]。有研究表明,復合種養系統中藻類不僅能通過光合作用增加水體中的氧氣,還能加快尿素的水解,對氮元素轉移與轉化十分重要,然而過多的藻類又會因對氮元素的競爭作用影響作物的生長[24]。此外,復合種養系統中浮游植物群落多樣性的增加,有助于形成更復雜的食物網結構,從而提升生態系統的穩定性和生產力[25]。研究不同種養系統中浮游植物群落結構及其與環境因子的關系可為制定合理的水質、藻相管理措施提供科學依據。
目前,關于漁農復合種養模式中水體的浮游植物群落特征研究,多以稻漁復合種養為主,缺乏對山區蓮漁復合種養模式的研究;對漁農復合種養模式的效益評價多以直接收益法和產投比法[8,26]為主,或通過其化肥和農藥減施量描述其生態效益[4],缺乏對其經濟和生態效益統一量綱的、定量的綜合評價。本文選擇江西省井岡山地區具有代表性的蓮魚復合種養模式作為研究對象,以當地魚單養和蓮單種模式作為對照模式,研究該復合模式中浮游植物的群落結構特征,采用冗余分析研究該模式中驅動浮游植物群落動態變化的環境因子,基于生態足跡法[27,28]對其生態效益進行統一量綱的分析。為蓮魚復合種養模式的水質、藻相管理提供理論基礎,為漁農復合種養模式的效益評價提供參考。
1 材料與方法
1.1 研究地點
研究地點位于中國江西省吉安市井岡山市(東經113°49′—114.23′,北緯26°27′—26°49′),該地區年平均溫度19℃,年降水量1597.5 mm,年日照時數1698.7h,年平均濕度可達78%。其中,蓮魚復合種養模式及魚單養模式均來自同一生產基地(大隴,圖 1),蓮單種模式來自該地區另一生產基地(田頭,圖 1),兩研究地點相距41 km,采集的池塘面積均在(0.133±0.05) hm2,兩個基地種植的蓮品種均為太空蓮36號。太空蓮36號全生育期(移栽至采摘結束)200—210d,井岡山地區種植的太空蓮在春季開始立葉,秋季下旬結束采摘,本研究在該時期進行。蓮魚復合種養模式環溝寬度在1—1.5 m左右,溝深1.5 m。蓮魚復合種養及魚單養模式中放養的均為中科五號異育銀鯽,蓮魚復合種養模式放養量為195 kg/hm2,魚單養系統放養量為225 kg/hm2。

圖1 研究地點位置Fig.1 The locations of two study sites
1.2 生產調查及樣本采集
生產調查通過實地勘查、訪問農戶、查閱生產記錄本及調查問卷獲取生產數據。采集的數據包括水產品的放養量、收獲量、人工成本、田租、投飼量、用水量、肥料及農(漁)藥施用量,并通過其單位面積的收入減去單位面積的成本得到單位面積利潤。
水樣、浮游植物的采集、測定在4月(春季)、7月(夏季)和10月(秋季)對蓮魚種養、魚單養和蓮單種模式各三個池塘進行水樣和浮游植物的采集。水樣采集依據 HJ 494—2009《水質采樣技術指導》和 SL 187—1996《水質采樣技術規程》所要求的方法,采集的水樣保存在1 L水樣瓶中。除使用便攜式水質測定儀YSI現場測定溶氧、水溫和pH外;其他參數均通過現場固定后運回實驗室進行測定。浮游植物的采集參照《淡水浮游生物研究方法》[29],浮游植物定性樣品用25#浮游生物網在水深0.5 m處緩慢劃“8”字進行采集,拖拉時間3min。定量樣品使用5 L的有機玻璃采水器在水深50 cm處進行采集,取1 L水樣加入魯哥氏液固定后帶回實驗室進行分析。將水樣沉淀并濃縮至50 mL后于顯微鏡下計數。參照相關文獻[30]進行藻類種類鑒定,參照《淡水浮游生物研究方法》對浮游生物樣品計數及計算。
1.3 計算公式
浮游生物多樣性指數根據香農-威納多樣性指數(H',Shannon-Wiener index)、Pielou 均勻度指數(J,Pielou)和Mcnaughton 優勢度指數(Y,Mcnaughton index)對不同系統水體中的浮游生物進行分析評價[31],各指數的計算公式如下:
(1)H'=-Σpi×lnpi
(2) Pielou 均勻度指數J=H'/log2S
(3) Mcnaughton 優勢度指數Y=pi×fi
在此過程中,技術要點如下:(1)對電力系統的調峰能力進行科學合理的分析;(2)對系統的調頻能力進行科學合理的分析;(3)對電力系統傳輸能力進行分析;(4)將消納能力作為目標,對電力系統進行科學合理的規劃。
式中,ni為i種的個體數,N為所有種類總個體數,H'為香農-威納多樣性指數,S為物種數,pi=ni/N,fi為該物種在各池塘中出現的頻率。本研究中將Y>0.02的浮游植物定為優勢種,Y>0.1為強優勢種。
生態足跡生態足跡法通過計算一定區域內消費及排放廢棄物所需要的生態生產性面積來表示發展造成的生態負荷,計算公式如下:

式中,EF為生態足跡總量,i為消費項目類型,j為生物生產性土地類型(分為耕地、林地、草地、水域、建筑用地和能源用地),rj為均衡因子,Ai為第i種消費項目折算的生物生產面積,Ci為第i種消費項目的消費量,Yi為生物生產性土地生產第i種消費項目該地區年平均產量。在本文中,飼料、苗種、水、農(漁)藥、肥料氮、肥料磷和Ci值是通過生產調查得到的數據,水污染氮和水污染磷的Ci值是通過水質調查得到的水體中年平均總氮和總磷含量減去GB 3838-2002《地表水環境質量標準》中對該類水體規定的上限值得到的。該地區水資源年平均產量來源于2016—2019年《吉安市水資源公報》,飼料(糧食)、苗種(水產品)及生產性土地面積年平均產量來源于井岡山市2015—2019年《年經濟和社會發展統計公報》。土地對農(漁)藥、化肥中N、P的消納能力及水體對N、P的消納能力見文獻[32]。
1.4 數據處理
數據處理及分析均在R 3.6.0中完成。采用單因素方差分析(ANOVA)中的多重比較(Tukey HSD)分析不同系統水體理化因子及浮游植物密度差異。采用Pearson檢驗分析不同水體理化因子之間的相互作用及水體理化因子對浮游植物密度、Shannon-Wiener指數和Pielou 均勻度指數的影響。
使用R 3.6.0 中的vegan包進行浮游植物與環境因子的排序分析,用于排序分析的物種數據篩選標準:(1)在各樣點出現的頻率>12.5%,(2)物種的Mcnaughton 優勢度指數>0.02。對篩選出的物種密度數據進行Hellinger正態轉化,對環境因子數據進行正態性檢驗,對不符合正態分布的數據進行轉化。隨后使用Decorana函數對物種數據進行除趨勢對應分析,進一步得到各排序軸的物種變化梯度長度。
2 結果
2.1 浮游植物種類組成、密度及優勢種
共采集到裸藻門(Euglenophyta)、隱藻門(Cryptophyta)、黃藻門(Xanthophyceae)、藍藻門(Cyanophyta)、綠藻門(Chlorophyta)、硅藻門(Bacillariophyta)和金藻門(Chrysophytes)浮游植物7門103種,其中蓮魚種養模式90種,魚單養模式85種,蓮單種模式76種,3種模式的浮游植物種類數均為夏季>春季>秋季。在研究期間,采集到的綠藻門種類數最多,為54種,黃藻門種類數最少,僅有1種,各門種類數量排序為:綠藻門>藍藻門>硅藻門>隱藻門>裸藻門>金藻門>黃藻門。

圖2 蓮魚種養、魚單養和蓮單種模式水體中不同季節浮游植物細胞密度變化Fig.2 Seasonal densities of phytoplankton in the integrated lotusfish culture system,lotus monoculture system and fish monoculture system
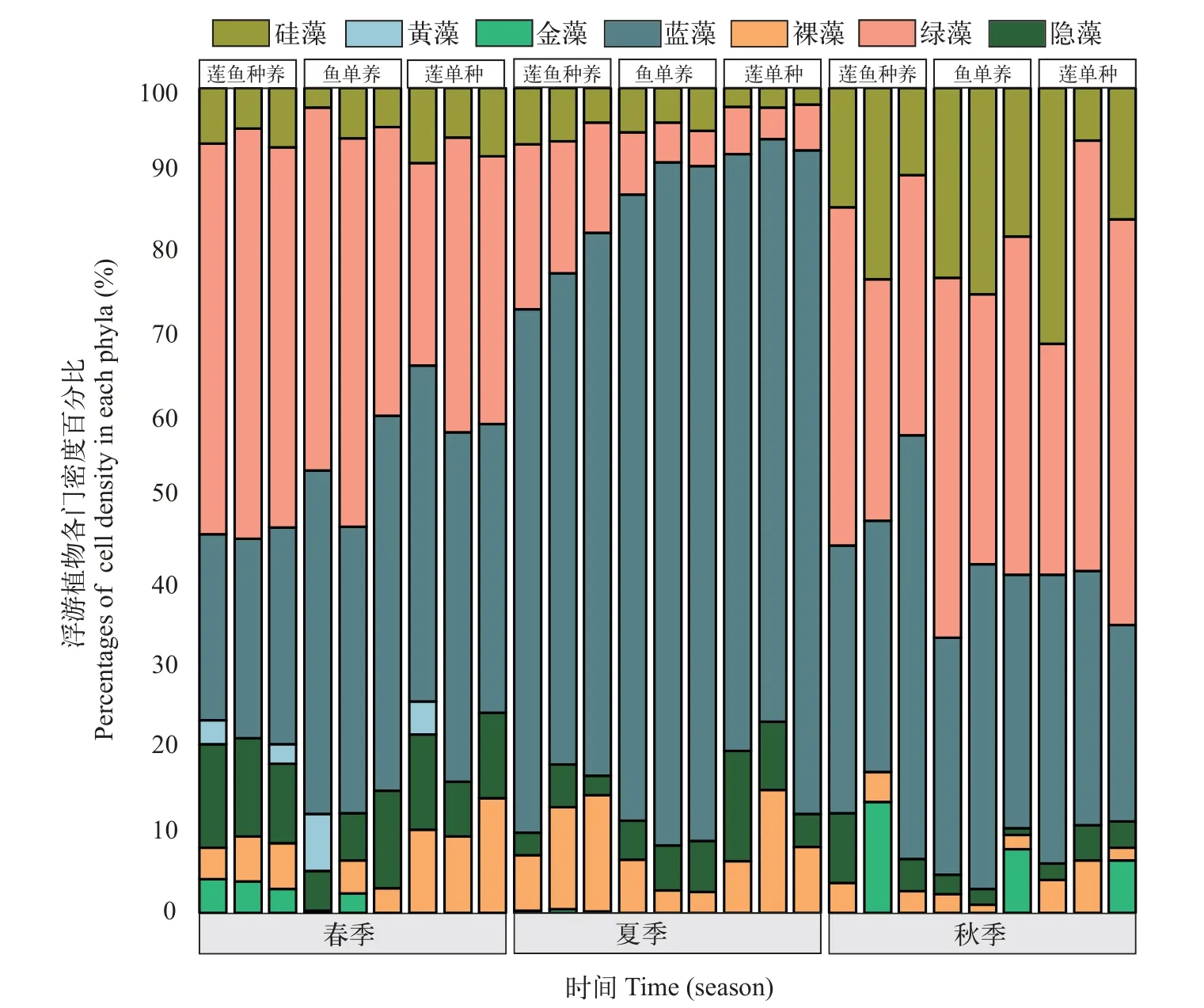
圖3 不同季節蓮魚種養、魚單養和蓮單種模式水體中浮游植物細胞密度百分比Fig.3 Seasonal density in percentages for each phyla in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
在調查期間,3種種養模式水體中的優勢種組成及優勢度具有明顯差異(圖4和表 1)。蓮魚復合種養模式水體中春季優勢種為卵形隱藻(Cryptomons ovata;0.098)、美麗顫藻(Oscillatoria formosa;0.046)、針狀藍纖維藻(Dactylococcopsis acicularis;0.047)、四尾柵藻(Scenedesmus quadricauda;0.153)和雙對柵藻(Scenedesmus bijuba;0.132);夏季優勢種為矩圓囊裸藻(Trachelomonas oblonga;0.080)、小席藻(Phormidium tenus;0.097)、顫藻(0.140)、銅綠微囊藻(Microcystis aeruginosa;0.241)和四足十字藻(Crucigenia tetrapedia;0.052);秋季優勢種為假魚腥藻(Pseudoanabaenasp.;0.050)、空球藻(Eudorinasp.;0.022)、橢圓小球藻(Chlorella ellipsoidea;0.062)和異極藻(Gomphonemasp.;0.066)。在魚單養模式中,夏季僅有藍藻門的藻類形成優勢種,且水華微囊藻、銅綠微囊藻和假魚腥藻形成強優勢種。在3種種養模式水體中均出現的優勢種有:卵形隱藻、四尾柵藻、小席藻和銅綠微囊藻。

表1 浮游植物優勢種代碼Tab.1 Codes of dominant phytoplankton species

圖4 不同季節蓮魚種養、蓮單種和魚單養模式中浮游植物優勢度指數Fig.4 Seasonal variation of Mcnaughton dominance index in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.2 浮游植物多樣性指數
研究期間,各模式的香農威納指數和均勻度指數均呈現出先下降后上升的趨勢(表2)。其中,蓮魚復合種養模式的香農威納指數變化為3.021—3.261;Pielou均勻度指數變化為0.502—0.645。魚單養模式的香農威納指數變化為2.649—2.882;Pielou 均勻度指數變化為0.454—0.613;蓮單種模式的香農威納指數變化為2.527—3.061;Pielou 均勻度指數變化為0.452—0.602。各模式香農威納指數H′均值排序為蓮魚復合種養>蓮單種>魚單養,Pielou均勻度指數均值排序為蓮魚復合種養>蓮單種>魚單養,但3種模式水體中的浮游植物香農威納指數和均勻度指數沒有顯著差異(P>0.05)。

表2 不同季節蓮魚種養、蓮單種、魚單養模式浮游植物香農威納指數及均勻度指數Tab.2 Seasonal variation of Shannon-Weiner index and Pielou index in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.3 水體中的環境因子
蓮魚復合種養模式及其單種、單養模式水質呈現出一致的季節變化,水體中總氮(TN)、總磷(TP)、氨氮(NH4-N)、硝酸鹽氮(NO3-N)、亞硝酸鹽氮(NO2-N)和水溫(WT)在復合種養期間均呈現出先升后降趨勢,而pH和DO則呈現出先降后升趨勢(圖5)。單因素方差分析結果顯示,春季蓮魚復合種養模式和蓮單種模式的TN顯著高于魚單養模式,蓮魚復合種養模式水體中的TP含量顯著低于魚單養模式(P<0.05),蓮魚復合種養模式水體中的NO3-N含量顯著低于魚單養模式(P<0.001),但顯著高于蓮單種模式(P<0.001),水體中NO2-N含量顯著高于蓮單種模式(P<0.001),水體中溶氧顯著高于魚單養模式(P<0.01),水體中pH顯著高于蓮單種模式(P<0.05);此外,春季各模式間的WT和NH4-N沒有顯著差異。夏季蓮魚復合種養模式水體中TN顯著高于魚單養模式(P<0.001)。與春季相反,在夏季蓮魚復合種養模式水體中TP含量顯著高于魚單養模式(P<0.05)。秋季蓮魚復合種養模式的TN含量顯著高于魚單養模式(P<0.05),TP含量在各模式中沒有顯著差異(P<0.05),NO3-N含量顯著低于魚單養和蓮單種模式(P<0.001),NH4-N含量顯著低于魚單養模式(P<0.001),溶氧顯著高于魚單養和蓮單種模式(P<0.001;圖 5)。
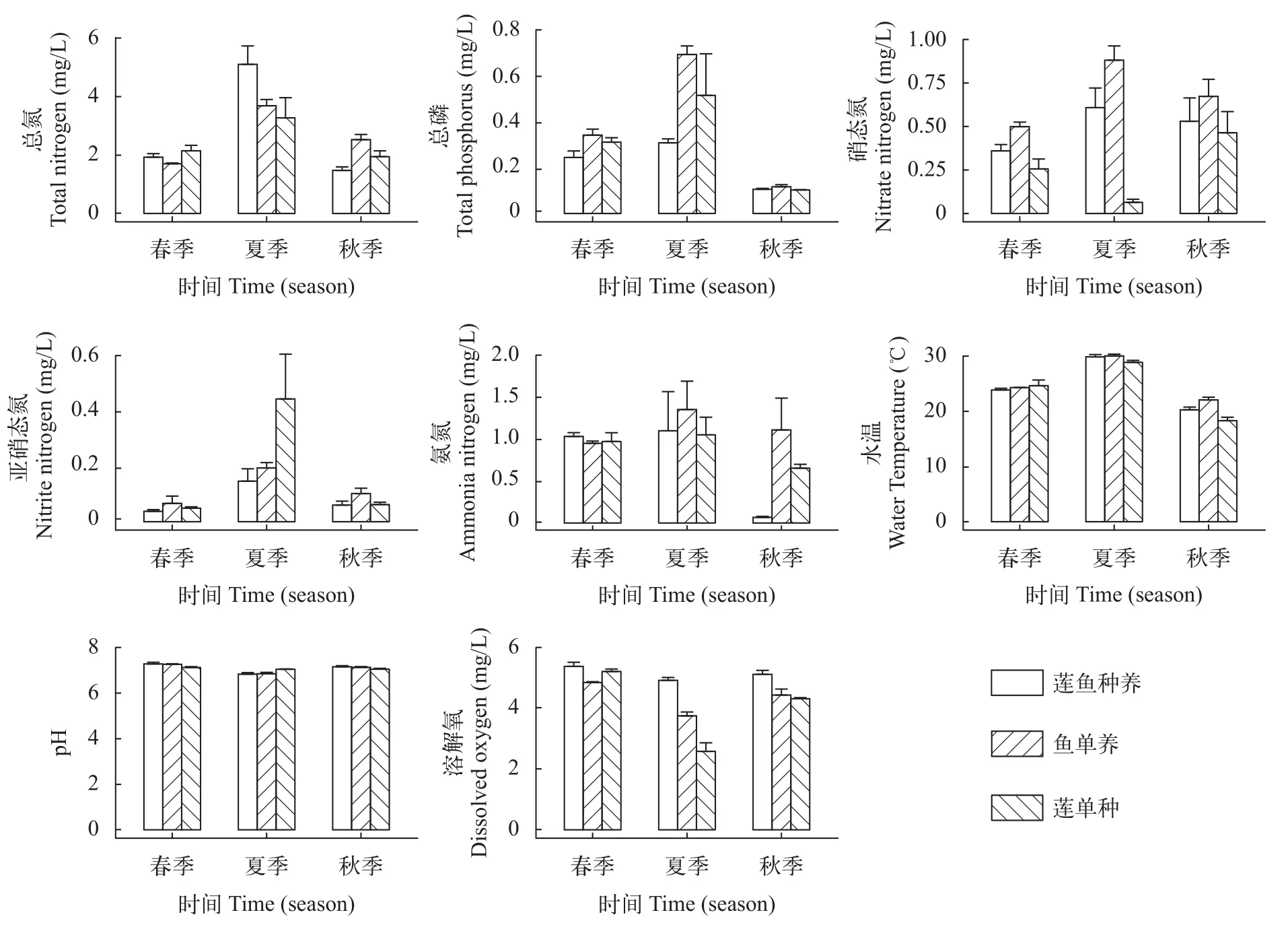
圖5 蓮魚種養、蓮單種和魚單養模式水體中不同季節水體理化因子Fig.5 Seasonal variation of environmental factors in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.4 環境因子和浮游植物群落的關系
本研究中蓮魚復合種養模式水體中環境因子和浮游植物群落的關系排序軸中梯度長度的最大值為2.142,使用RDA模型結果更為準確;魚單養和蓮單種模式中排序軸中梯度長度的最大值分別為3.775和3.368,既可使用CCA模型又可使用RDA模型進一步分析。為確保分析方法的統一性,本研究均采用RDA模型進行分析。進行初步RDA分析后,使用ordistep函數進行前向選擇,保留顯著影響浮游植物群落結構特征變化的環境因子(P>0.05),并進一步構建主要環境驅動因子和浮游植物群落結構特征相互關系的簡約模型。隨后對模型的R2進行矯正,并對約束軸進行蒙特卡羅置換檢驗,確保RDA模型、第一約束軸和第二約束軸均顯著(P<0.05)。簡約模型顯示,各季節樣方水體中浮游植物群落具有明顯差異(圖6)。其中,蓮魚復合種養模式浮游植物群落的主要驅動因子為WT和DO,對浮游植物群落特征的解釋率分別為28.48%和24.36%;魚單養模式浮游植物群落的主要驅動因子為TP和DO,對浮游植物群落特征的解釋率分別為39.47%和33.52%;蓮單種模式游植物群落的主要驅動因子為WT、NO3-N和DO;對浮游植物群落特征的解釋率分別為45.56%、17.37%和5.06%。

圖6 蓮魚種養、蓮單種、魚單養模式水體中浮游植物群落和環境因子的冗余分析Fig.6 RDA analysis of phytoplankton communities and environmental factors of the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.5 經濟效益和生態足跡
蓮魚種養每公頃可產生經濟效益2.52萬元,每公頃利潤為魚單養>蓮魚種養>蓮單種(表3),而每公頃蓮魚模式的總生態足跡為11.27 hm2,每公頃蓮單種的總生態足跡為18.91 hm2,每公頃魚單養模式的總生態足跡為19.54 hm2,魚單養和蓮單種帶來的生態負荷分別為蓮魚復合種養模式的1.73倍和1.68倍。此外,蓮魚復合種養模式單位利潤的生態足跡量為4.48 hm2,蓮單種模式單位利潤的生態足跡量為13.48 hm2,魚單養模式單位利潤的生態足跡量為5.54 hm2。即蓮魚模式每產生1萬元的經濟效益需要該地區4.48 hm2的生產土地來提供其所需投入品或消納其產生的廢物,魚單養模式則需要5.54 hm2生產土地,蓮單種模式則需要13.48 hm2生產土地。

表3 蓮魚種養、魚單養、蓮單種模式的生態足跡Tab.3 Ecological footprint of the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
3 討論
3.1 水體理化因子和浮游植物群落結構特征
本研究發現蓮魚復合種養模式及其單種、單養模式水體中TN、TP、NH4-N、NO3-N、NO2-N和WT在研究期間均呈現出先升后降趨勢,而pH和DO則呈現出先降后升趨勢。在本研究中魚單養模式和蓮單作模式水體中溶解氧在調查期間,尤其是夏季相比蓮魚種養模式較低,表明水體內有機負荷相對蓮魚種養模式較高[33];而蓮單種模式水體中溶氧在夏季顯著低于其他模式則更可能與蓮葉的過度生長及蓮葉和浮游植物對陽光的競爭有關。氮、磷等營養物質是浮游植物生長繁殖的物質基礎,也是在水質監測評價中重點關注的指標[34]。在本研究中,夏季蓮魚種養模式水體中TN含量達到高峰,且顯著高于單養/種模式,可能是由于共作過程中的施肥和投餌共同導致的。而在蓮魚種養模式水體中TP含量始終低于魚單養模式,這可能與蓮生長期對磷元素的旺盛需求有關。
浮游植物群落存在明顯的季節演替現象,研究期間各模式水體中浮游植物的演替規律與Sommer等[35]提出的演替模式基本一致。在各模式水體中,隱藻常在春夏季形成優勢種,這可能與隱藻能通過鞭毛調節自身位置并適應低光條件有關[36]。而藍藻門在各個季節均存在優勢種,與藍藻具有的可調節細胞浮力的氣泡有關,這種能力使得藍藻可以適應不同季節并廣泛分布[37]。秋冬季優勢度較高的異極藻和小環形藻均屬于硅藻門,而這些藻類的最適溫度一般低于25℃。在研究期間,春季魚單養和蓮魚種養模式水體中浮游植物密度均顯著高于蓮單作模式水體中浮游植物密度,一方面與鯽對浮游植物的攝食壓力弱有關[38],另一方面可能與鯽作為底層魚類,可通過生物擾動作用促進營養物質從底泥中釋放,從而引起水體中浮游植物生物量增加有關[39]。此外,復合種養模式各個季節的香農威納指數均比單養/種模式高,說明種養模式中浮游植物群落多樣性高;而復合種養模式中浮游植物群落多樣性的增加,有利于形成更復雜的食物網結構,提升模式的穩定性和生產力[25]。而在魚單養模式中,夏季僅有藍藻門的藻類形成優勢種,對水體中浮游植物密度的貢獻最大,優勢度較高的種類將占據更多生態位[40],從而導致群落結構穩定性下降,增加發生水華的風險。
3.2 水體理化因子對浮游植物群落結構特征的影響
浮游植物群落結構受外界環境中溫度、營養鹽等環境因子的驅動,始終處于動態變化過程之中[41]。溫度和營養鹽是影響浮游植物生長的兩大重要環境因素。水溫作為顯著影響蓮魚種養和蓮單種模式中浮游植物群落特征的共同環境因子,一方面影響浮游植物的新陳代謝速率,另一方面也通過影響各類營養物質在水體中的溶解度影響浮游植物生長和繁殖[42]。通常浮游植物密度會隨著溫度上升而上升,且較高的溫度有利于綠藻門和藍藻門的生長和繁殖[43]。由于藍藻對溫度的耐受性高于其他藻類,在高溫下通常以快速生長的r對策獲得競爭優勢[44],這與本研究中藍藻門中的銅綠微囊藻、水華微囊藻、不定微囊藻和顫藻在高溫季節的高細胞密度結果一致。眾多研究表明水體中氮、磷等營養鹽對藻類的生長均具有限制作用,氮磷比(TN/TP)通常用來判斷氮或磷是否為限制性因素,當氮磷比>16時,則認為磷是限制性因素;反之,則認為氮是限制藻類生長的主要因素[45,46]。在本研究中魚單養和蓮單種模式中秋季氮磷比均大于16,蓮魚模式中秋季氮磷比接近16(13.36—15.43),秋季浮游植物密度較低,水體均處于極貧營養狀態。冗余分析表明,氮、磷營養鹽并不是影響蓮魚種養模式水體中浮游植物群落特征的主要影響因素;而水體中總磷含量則是影響魚單養模式水體中浮游植物群落特征的主要影響因素,且與微囊藻等藍藻的高密度密切正相關。而在蓮單種模式中,浮游植物還受到硝酸鹽的調控。冗余分析結果顯示,銅綠微囊藻、不定微囊藻與水體中NO3-N含量呈負相關,這可能與微囊藻細胞能將NO3-N轉化為其他形式的氮排出細胞外有關[47]。在魚單養模式中,藍藻類中的假魚腥藻、銅綠微囊藻、水華微囊藻、小席藻和TP高度正相關,且夏季水體中總磷含量顯著高于蓮魚種養模式。而隨著溫度上升底質磷的釋放可導致水體中可利用的磷酸鹽增多[48],導致上述浮游植物生物量增加。
3.3 水體富營養化和污染狀況
浮游植物群落結構特征與水體環境密切相關,且能對水體變化做出快速響應,主要表現在種類組成、細胞密度和優勢種的變化。因此,浮游植物群落結構和多樣性的變化常用于水體營養狀態、污染程度評價[49]。根據浮游植物密度[50],春季蓮魚種養模式水體屬于貧中營養,魚單養和蓮單種模式均屬于貧營養狀態;夏季蓮魚種養模式和蓮單作模式均處于中營養狀態,魚單養模式處于富營養狀態;秋季3種水體均處于極貧營養狀態。為避免單一多樣性指數評價的局限性,本研究選用Shannon-Wiwner多樣性指數和Pielou均勻度指數共同進行評價。根據Shannon-Wiwner多樣性指數評價標準[50],蓮魚種養模式3個季節水體均處于和均勻度指數得到的蓮魚種養模式各季節均處于輕污染-無污染狀態,魚單養各季節均處于β-中污狀態,蓮單種模式除秋季水體屬于輕污染-無污染狀態外,其他季節均處于β-中污狀態。根據Pielou均勻度指數,蓮魚種養模式3個季節水體均處于和均勻度指數得到的蓮魚種養模式各季節均處于輕污染-無污染狀態,魚單養和蓮單種模式除夏季屬于β-中污狀態,其他季節均屬于輕污染-無污染狀態。在本研究中,較高的浮游植物多樣性指數與水體的富營養化程度并不完全一致,與前人研究結果一致[51,52]。一方面可能與水體受到干擾導致其生物多樣性和水體的水質情況有悖有關,另一方面本研究中未對浮游植物進行功能群分類,根據生物多樣性指數得到的水質評價結果可能比實際偏高。雖然,浮游植物密度和兩種指數的評價結果顯示,各模式中水體富營養化程度不完全一致,但總體而言各模式中水體富營養化程度的季節變化均為夏季>春季>秋季,且蓮魚種養模式的水質優于單養/單種模式,可能與蓮魚種養模式中的蓮作為高等水生植物具有一定的抑藻效應[53],能抑制藻類優勢種的生長、擴散有關。
3.4 蓮魚種養模式的生態成本
生態足跡模型[27,28]從1999年引入我國后,被廣泛應用于國家、地區、社區、學校、行業乃至個人的生態負荷計算,為度量可持續發展提供了一個“公平秤”,并逐漸開始應用在漁業和種養業的可持續發展衡量中,進而為政策制定提供參考依據。如通過計算漁業集約化養殖模式的生態足跡為具體生產管理提供理論基礎,通過計算大菱鲆流水養殖和循環水養殖養殖場生態足跡為中國大菱鲆循環水養殖提供參考建議等[54,55]。在本研究中蓮魚種養每公頃可產生經濟效益2.52萬元,平均每公頃利潤低于魚單養模式的3.53萬元,高于蓮單種模式的1.40萬元,經濟效益適中;然而蓮魚復合種養模式每公頃所產生的生態足跡為11.27 hm2,其用來提供其所需投入品或消納其產生的廢物的生產土地遠低于單養/種模式,表明復合系種養統具有更好的可持續性,生態效益和經濟效益更高,符合循環經濟理念。在井岡山地區,太空蓮雖然被廣泛種植,然而蓮子和蓮藕的產量極易受天氣影響,給種蓮的農戶們帶來了更多的成本風險和收益不確定性。而蓮魚種養則提供了一種更為經濟高效、生態環保的可持續發展模式。
