六種沉水植物的克隆生長特征
2022-08-04 15:15:56姚軼倩袁昌波呂超超曹特張霄林倪樂意
水生生物學報 2022年7期
姚軼倩 袁昌波 呂超超 曹特 張霄林 倪樂意
(1.中國科學院水生生物研究所,武漢 430072;2.中國科學院大學,北京 100049)
沉水植物作為淡水生態系統中重要的初級生產者,在維持淡水生態系統“清水穩態”中發揮著極為重要的生態功能[1—3]。然而,隨著水體富營養化程度的加劇,沉水植被衰退和消失的現象在湖泊、河流等眾多水體內普遍出現。近二十余年,人們逐步認識到沉水植被在凈化水質和修復淡水生態系統中的重要作用,因此沉水植被的恢復越來越受到重視[4,5]。但是,相比于近年來全國范圍內廣泛開展的沉水植被恢復工程,我們對沉水植物種群建立、維持和擴張等過程的模式仍缺乏足夠深入的了解,在對沉水植物生長模式的理解與恢復沉水植物的實踐之間存在錯配,這種錯配將影響沉水植物恢復工程效果的實現和穩定。此外,在很多生態學領域的研究模型中,沉水植物的克隆生長模式仍存在空缺,很多模型僅以生物量作為沉水植物模塊的參數,并沒有考慮到沉水植物種群空間擴張特征及由此引發的種間相互作用,這將會限制模型在描述和預測淡水生態系統過程中的科學性和準確性。我們需要獲取不同沉水植物生長模式的關鍵參數,并依此構建不同物種的生長模型,基于此類模型在考慮種間相互作用關系后搭建出更為復雜的種群和群落動態模型,才能科學地解決上述理論研究與工程實踐中遇到的問題。
克隆生長(Clonal growth)是指在自然條件下植物產生具有實際或潛在獨立生存能力的分株(Ramet)的營養生長過程[6,7]。與陸生植物相比,克隆生長在水生植物中更為普遍也具有更高的重要性[8]。植物的克隆構件(Clonal module)是克隆生長中的重要功能單元,一般分為分株和間隔子(Spacer)[9],分株是指具有完整生物學功能的完整植株,包括地上直立莖、分枝和葉,及地下根系;間隔子是指連接相鄰分株之間的匍匐莖或根狀莖。本研究中的分株是指不包括間隔子的具有完整根、莖和葉的獨立植株。水生植物通過克隆生長不斷產生克隆分株進行分散發育和繁殖,并通過間隔子在不同分株之間進行生理整合[10]。基于這種克隆生長特性,水生植物具有很強的種群擴張能力、空間競爭能力及對異質生境的適應能力,能夠很快地占據生存空間,進行資源的吸收和利用,并在相連接的克隆分株之間進行資源共享,進而提高種群的適合度[11,12]。
作為一種維持和擴張種群的方式,克隆生長長期受到研究者的關注[13—15]。張偉濤[16]的研究表明,香蒲 (Typha orientalis) 在不同水位梯度下有不同的克隆構型,間隔子長度與分株數存在顯著的負相關關系;符輝等[17]的研究表明,苦草 (Vallisneria natans) 的克隆分株數和間隔子總長隨水深增加顯著降低;Xiao等[18]的研究表明,苦草的生物量和分株數在光異質生境中高于光同質生境;Villamagna和Murphy等[19]的研究表明鳳眼蓮 (Eichnornia crassipes)因其旺盛的克隆生長成為世界性入侵種并泛濫成災。但是這些研究更多是從表型可塑性變異對異質生境的響應方面探討水生植物的適應性能力,對水生植物克隆生長基本模式和基本參數的探索仍需加強,例如克隆分株的形成速率、分株數量和質量的分布,間隔子的長度,單一克隆株種群的面積擴張速率等,這些都是與克隆生長和種群擴張密切相關的基礎生物學參數,并且這些參數可能具有物種特異性,這些特異性也將影響到不同物種的種群構建。對克隆生長模式的研究是一項非常基礎的工作,基于這方面的研究可以獲得非常重要的生長參數,從而為構建不同沉水植物克隆生長模型以及種群動態模型提供非常有意義的信息。因此,我們對沉水植物中的一些代表物種開展研究,通過同質園實驗(Common garden experiment)的方法,跟蹤觀測單個無性系中所有克隆構件的動態變化趨勢,探討不同物種克隆生長關鍵生物學參數及獲取空間資源策略方面的特征,總結不同物種的克隆生長模式,為沉水植物種群擴張模型和群落模型的構建提供參考,并為沉水植被修復工程提供科學支撐。
1 材料與方法
1.1 實驗地點與實驗單元
洱海位于云南省大理市,是云南省九大高原湖泊之一,具有非常高的水生植物多樣性[20,21]。根據調查發現,洱海中目前分布有沉水植物22種(其中眼子菜屬有12種),為研究沉水植物克隆生長提供了豐富的實驗材料來源。實驗地點位于大理市上關鎮洱海紅山灣水生植物種苗繁育基地北區(25°56.8′N,100°8.4′E)。將繁育基地中的一個面積約480 m2的獨立種苗塘作為同質園。排干塘內水體后,將塘底淤泥進行翻耕,清除其中的水生植物斷莖、休眠芽和種子等可再次萌發的器官,在曬塘一個月后,使用空心磚構建邊長為1 m的正方形實驗單元(圖1)。
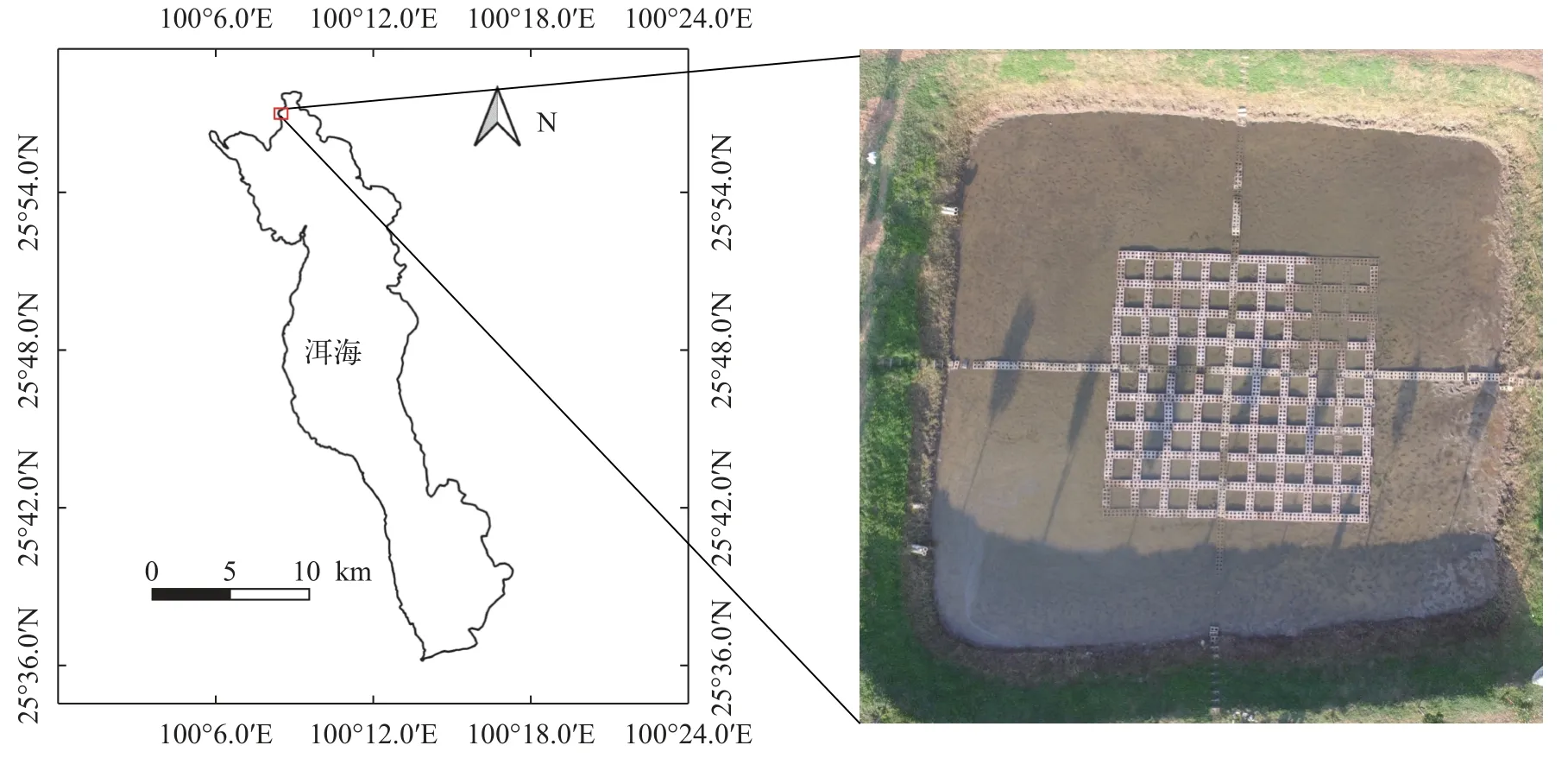
圖1 實驗地點及同質園Fig.1 Experimental site and common garden
1.2 實驗材料
眼子菜屬和苦草屬植物是我國淡水水體中常見的沉水植物類群。近年來,這些物種常被用于湖泊、河流等水體生態修復工程中。因此,對這類沉水植物克隆生長特性的研究也具有較強的實踐指導意義。本研究選擇了具有代表性的6個物種,均為多年生沉水植物,包括竹葉眼子菜(Potamogeton wrightii)、眼子菜(P.distinctus)、光葉眼子菜(P.lucens)、穿葉眼子菜(P.perfoliatus)、扭葉眼子菜(P.intortifolius)和苦草。實驗材料采集于洱海近岸淺水區(水深不超過100 cm),均為根、莖和葉齊全的完整植株。實驗前將采集到的沉水植物清洗干凈,清除植株表面的附著藻、魚卵和泥沙等雜質,并測量單株株高和生物量鮮重(以下簡稱生物量)。
1.3 實驗設計
在每個物種中選擇9株根、莖和葉完整,并且株高和生物量基本一致的植株(不包含匍匐莖)作為本實驗中的母株,母株的初始株高和生物量如表 1所示。每個實驗單元內種植一株,將所有母株在同一天內種植在每個實驗單元的中心點,將其作為原點建立各實驗單元的平面直角坐標系。每個物種設置9個重復,共計54個實驗單元。

表1 六種沉水植物初始株高和生物量(平均值±標準差)Tab.1 Initial height and biomass of 6submersed macrophytes (mean±SD)
實驗于2019年8月31日開始,在當天完成全部種植后立即注入洱海湖水至50 cm水深,培養1周后開始定期觀測。每次觀測時首先使用相互垂直的長度1 m的直尺對每個單元中新增分株的坐標進行確定,并在已確定坐標的分株上懸掛標簽進行標記(標簽上注明時間和坐標,此后對于已標記的分株僅測量株高增長情況),之后測量各分株的株高,計數單元內的所有分株數。此觀測工作每隔7d重復一次,直至實驗結束。在觀測活動中,實驗人員全程借助單元四周的空心磚進行移動,最大限度降低觀測活動對水體和底泥的擾動。為了防止實驗單元邊界對實驗材料克隆擴張造成空間限制從而影響實驗結果,62d后在非常接近實驗單元邊界處發現有分株形成時結束實驗。
為了評價實驗周期內的環境指標,在實驗池塘四周及中心位置設置5個點位監測水體環境指標,每隔7d現場測定5個點位的水溫,同時采集5個點位水樣帶回實驗室測定水化學指標(總氮TN、總磷TP、硝態氮、銨態氮和正磷酸鹽測定方法參照《水和廢水監測方法(第四版)》,實驗期間水體環境指標如表 2所示。試驗期間定期清除雜草以消除對實驗材料的影響。

表2 實驗區域水環境指標(平均值±標準差)Tab.2 Water environment index in experimental area (mean±SD)
1.4 克隆生長參數測定
在第62天,將每個單元內的所有實驗材料 (母株以及所有由間隔子連接的分株)全部挖出做好標記帶回實驗室洗凈,再用蒸餾水清洗3次。將與母株通過間隔子直接連接的分株記為 Ⅰ 級分株、與Ⅰ 級分株相連的分株記為 Ⅱ 級分株,依此類推直至所有分株編號完成。將所有間隔子與分株分離,間隔子編號與低級別分株保持一致。待測植株存放于4℃冰箱,測量工作由兩組同時進行,同步測量了分株株高、間隔子長度、分株生物量和間隔子生物量,36h內完成了收獲和測量工作。此后將所有樣品置于鼓風干燥箱中于80℃烘至恒重并稱量干重。
1.5 數據分析
使用單因素方差分析(One-way ANOVA)比較不同沉水植物在分株株高、分株形成速率的種間差異,使用S-N-K法對不同物種的組間差異進行事后多重比較,方差分析之前對所有數據進行正態分布和方差齊性檢驗。統計分析均使用SPSS 25完成,并使用Origin 2017軟件完成繪圖。
克隆擴張面積的計算采用了“任意多邊形面積計算公式”[22]。對于每個實驗單元,將同一方向或相近方向上距母株直線距離最遠的分株作為多邊形的頂點,并依此計算克隆系所占據的面積。
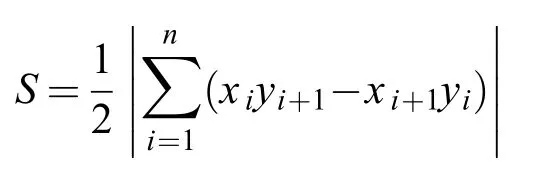
式中,式中S為多邊形面積,n為多邊形頂點數量,xn+1=x1,yn+1=y1。
2 結果
2.1 克隆構件形態特征
克隆構件形態特征中最大的一個差異在于間隔子(匍匐莖)上“節”的形成。如圖 2所示,苦草的間隔子完整光滑,與之不同的是5種眼子菜屬植物的間隔子均形成“節”,并在“節”上形成不定根,但是“節”不發育形成地上植株,因此“節”在克隆生長過程中僅提供固著作用及從底泥中吸收營養,沒有光合作用的功能。為了驗證間隔子“節”的形成在眼子菜屬中的普遍性,我們進一步廣泛地采集了不同生境中的眼子菜屬物種并對其匍匐莖進行觀察,發現眼子菜屬的大部分物種的間隔子上均形成“節”,我們認為匍匐莖上的“節”也可以作為眼子菜屬的一個分類特征。此外,從克隆構件的地上部分來看,苦草沒有形成地上直立莖,葉片全部基生于木質化的根莖;而眼子菜屬的物種均具有發達的直立莖和地上分枝,其分株具有明顯的高度優勢。本研究中涉及的6種沉水植物克隆構件的形態特征與其分類單元相一致。眼子菜屬植物具有相對發達的地上結構,形成較高的分株高度和生物量,其地下間隔子具有明顯的“節”;而苦草克隆構件的地上結構較為簡單,分株高度和生物量也相對較低,但是其地下間隔子不具有“節”,從而形成的分株密度相對較高。
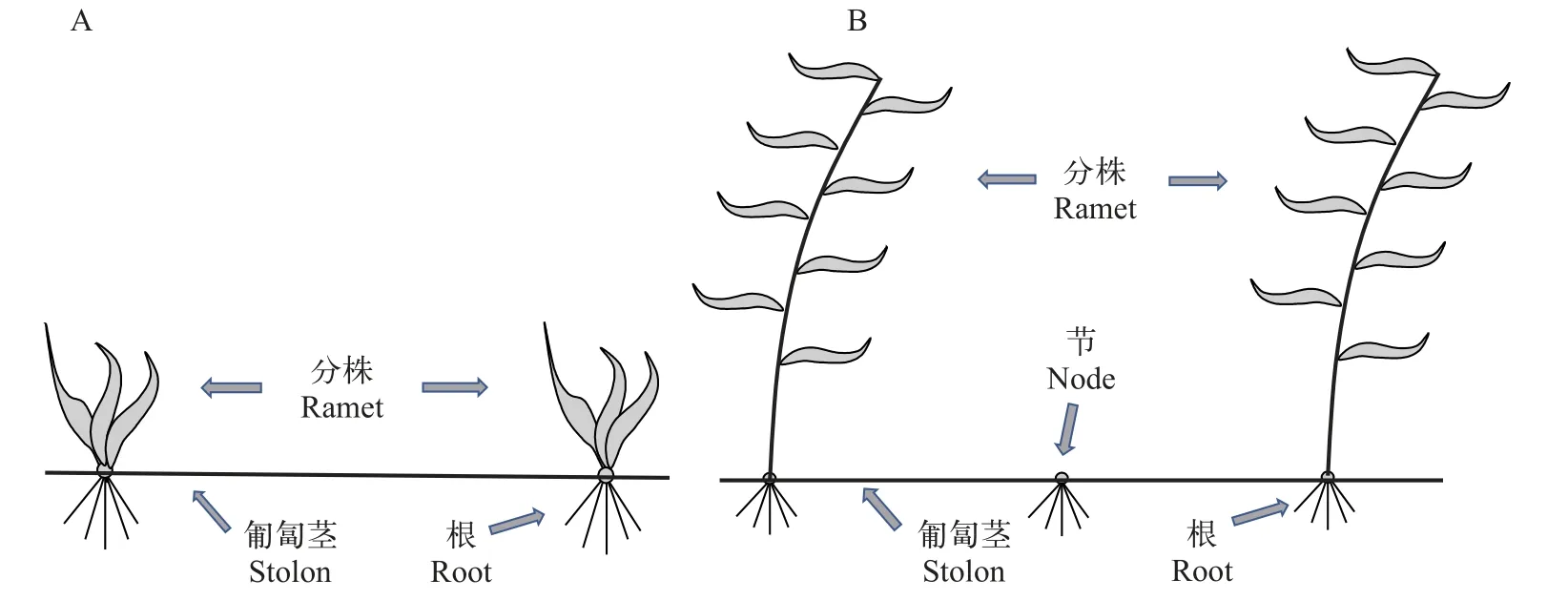
圖2 克隆構件特征(A.苦草屬;B.眼子菜屬)Fig.2 Clonal module (A.Vallisneria;B.Potamogeton)
2.2 克隆構件特征差異
我們從分株數、分株株高、分株生物量和間隔子長度(及間隔子總長)幾個方面對6種沉水植物的克隆特征進行了總結。在實驗結束時(第62天),根據不同克隆特征之間的相關性分析,我們發現分株數與分株株高之間存在顯著的負相關(P<0.001),而分株數與間隔子總長度表現出顯著的正相關(P<0.001)。
實驗結果表明,6種沉水植物的分株特征也表現出種間差異。如圖 3A所示,苦草的分株數量最高,且苦草、穿葉眼子菜和扭葉眼子菜的分株數顯著高于竹葉眼子菜、眼子菜和光葉眼子菜(P<0.05)。如圖 3B所示,克隆分株的平均生物量(鮮重)表現為穿葉眼子菜(2.19 g)>竹葉眼子菜>光葉眼子菜>扭葉眼子菜>眼子菜>苦草(0.59 g)。穿葉眼子菜的分株生物量顯著高于其他5個物種(P<0.05),竹葉眼子菜、眼子菜、光葉眼子菜、扭葉眼子菜和苦草之間沒有顯著差異。如圖 3C所示,與分株數結果相反的是,竹葉眼子菜(32.62 cm)和眼子菜(30.26 cm)的分株株高顯著高于苦草(12.42 cm)、穿葉眼子菜(21.92 cm)和扭葉眼子菜(20.02 cm)的分株高度(P<0.05)。如圖 3D所示,穿葉眼子菜、扭葉眼子菜和竹葉眼子菜的間隔子長度與光葉眼子菜、眼子菜和苦草之間差異顯著(P<0.05),且穿葉眼子菜的間隔子長度最長,與其他物種差異極顯著(P<0.001)。
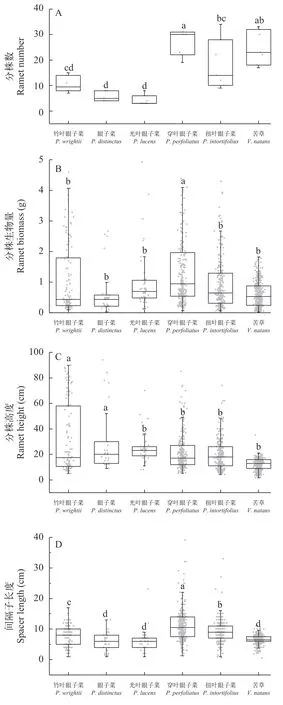
圖3 六種沉水植物克隆特征差異Fig.3 Differences in clonal characters of 6 species
2.3 種群擴張動態
分株數動態變化如圖 4所示,6種沉水植物在0—28d內基本都沒有分株產生,28d后,穿葉眼子菜、扭葉眼子菜和苦草的分株數表現出迅速增加;而竹葉眼子菜、眼子菜和光葉眼子菜的分株數增長較為緩慢。穿葉眼子菜和扭葉眼子菜分株數的最大增長速率均出現在第41天之后;苦草分株數在第55天之后表現出最大增長速率。
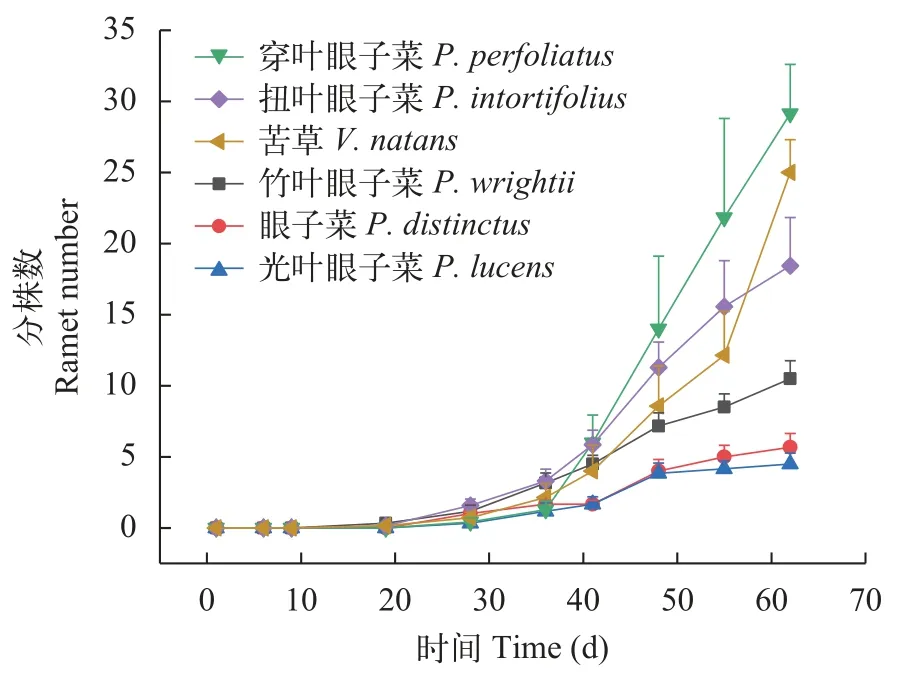
圖4 六種沉水植物的分株數變化Fig.4 The variations of ramet amounts of 6 species
面積擴張動態如圖 5所示,面積擴張速率也表現出類似分株數的種間差異,穿葉眼子菜、扭葉眼子菜和苦草的種群面積隨時間增加的速率超過其他3個物種(竹葉眼子菜、眼子菜和光葉眼子菜)。擴張面積隨時間的變化速率為穿葉眼子菜(113.22 cm2/d)>扭葉眼子菜(71.70 cm2/d)>苦草(35.48 cm2/d)>竹葉眼子菜(12.09 cm2/d)>眼子菜(3.07 cm2/d)>光葉眼子菜(0.53 cm2/d)。
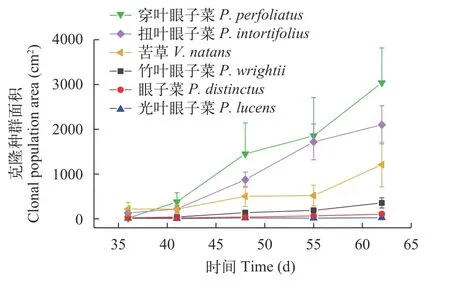
圖5 六種沉水植物克隆系面積擴張趨勢Fig.5 The increase of occupied area by ramets of 6 species
2.4 克隆生長參數
如表 3所示,間隔子上“節”的存在使眼子菜屬的5個物種(竹葉眼子菜、眼子菜、光葉眼子菜、穿葉眼子菜和扭葉眼子菜)兩分株間的匍匐莖上出現分級現象,而苦草匍匐莖無分級,這就導致苦草間隔子長度的變異程度最低,眼子菜屬的間隔子長度變異程度較高。在克隆生長的動態變化方面,苦草隨著時間的變化,分株數增加較快但株高增長緩慢,最大分株形成速率為1.84株/d。在眼子菜屬中,穿葉眼子菜和扭葉眼子菜的分株數增加最快,最大分株速率分別為1.14和0.78株/d;竹葉眼子菜(1.68 cm/d)和眼子菜(2.14 cm/d)的株高增長速率最大;光葉眼子菜幾乎未產生分株且株高增長緩慢。各物種的分株形成平均速率表現出顯著的種間差異(P<0.05),苦草和穿葉眼子菜的分株形成平均速率分別為1.09和1.00株/d,二者顯著高于其他物種。
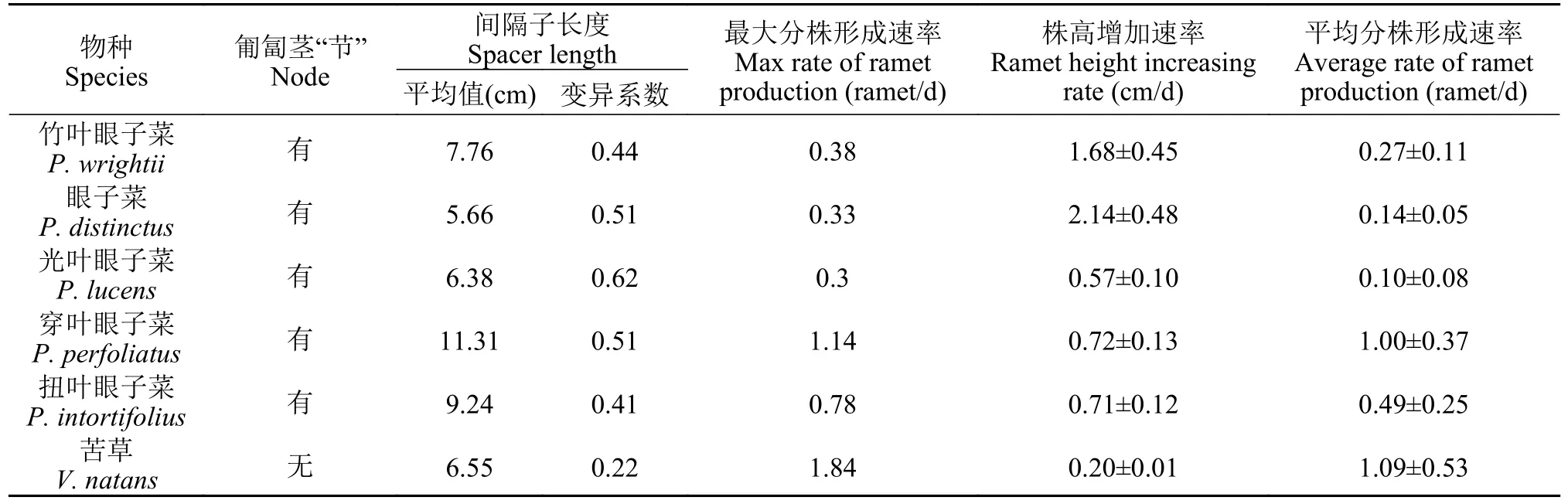
表3 六種沉水植物克隆生長的基本參數(平均值±標準誤)Tab.3 Basic parameters of clonal growth in six submersed macrophytes (mean±SE)
3 討論
沉水植物的克隆生長特性和克隆構件特征往往也是其形態分類特征。在本研究中我們發現眼子菜屬植物普遍具有的間隔子“節”的形成,根據本實驗結果以及我們對眼子菜屬物種的補充觀察,可以確定匍匐莖上“節”的形成是大部分眼子菜屬物種的固定性狀,可以作為一種分類特征。此外,這一性狀還具有兩方面的生態功能。一方面“節”的形成為間隔子長度的增加奠定了基礎。理論上,“節”的形成能夠支撐2倍匍匐莖長度的間隔子,而且“節”上形成的不定根能夠為較長的間隔子提供穩定的固著能力,從而穩定整個克隆系。沉水植物的克隆生長習性,使它在形態和生理等方面表現出一定的可塑性,并通過自身的可塑性變化實現對有效資源的獲取(如覓食行為等),分株間通過相互連接的匍匐莖或根狀莖進行各種資源的傳導[23—25]。Cain等[26]的研究表明一些克隆植物生長在有利生境中縮短匍匐莖(或根莖)節間長度或增加分枝來維持種群的生長。長的間隔子能使分株之間的空間跨度更遠,有利于分株對更遠處資源的獲取和利用,或者使分株更容易地“跨過”較為不利的底質區域,降低底質異質性對克隆系的影響。另一方面,匍匐莖上的“節”還有可能作為克隆生長過程中的“檢查點”(Checking point)。通過這一生長節點,植株將能夠對周邊生境進行識別,并對“節”后匍匐莖的形成策略做出選擇,可以大角度地改變原有的克隆擴張方向,從而更好地適應周邊的底質生境。因此,我們進一步推測,相比于苦草屬植物,具有形成“檢查點”能力的眼子菜屬植物將能夠更好地適應異質性底質或者形成游擊型的種群結構。而不具有“檢查點”能力的沉水植物(比如苦草屬植物)能夠通過更快地形成分株使克隆系占據較大的面積,可在局部形成密集型種群結構。
沉水植物克隆生長能力的不同,一方面取決于自身生物學特性的差異,另一方面,自然環境的異質性也是重要影響因素[27,28]。在異質生境中,克隆植物一般采用形態結構的可塑性(克隆可塑性)、分株的選擇性放置(覓食行為)、分株間物質傳遞與共享(克隆整合)等生態策略來最大限度地利用資源與空間[29—31]。但克隆植物也廣泛地存在于同質環境下,有研究指出,在淺水區域不受光限制的條件下,多種沉水植物可以通過增加匍匐莖長度、快速的克隆生長(克隆分株數增加)等行為來占據更多的空間[17,31]。本研究跟蹤觀測了各物種的單個無性系在同質環境中表現出的所有克隆構件的動態變化,旨在探究不同物種克隆生長特征種間差異。在環境一致的淺水區域,穿葉眼子菜和苦草產生克隆分株的速率最高,光葉眼子菜最低,幾乎不產生分株。在6種植物中,苦草的株高最低,竹葉眼子菜的株高最高,穿葉眼子菜的株高處于中等,但其分株生物量最高。此外,苦草的間隔子長度也顯著低于眼子菜屬物種。
在種群擴張面積方面,穿葉眼子菜、扭葉眼子菜和苦草比竹葉眼子菜、眼子菜和光葉眼子菜的擴張面積大;在種群擴張速率方面,穿葉眼子菜、扭葉眼子菜和苦草也表現出較高的擴張速率,按擴張速率由高到低排列為穿葉眼子菜>扭葉眼子菜>苦草>竹葉眼子菜>眼子菜>光葉眼子菜。不同物種的擴張速率的差異主要是由它們分株的形成速率和間隔子長度決定。有研究表明,間隔子的結構生長遵循特定物種的克隆規則[31],其在方向和長度上隨物種不同變化很大[13,32]。在本研究中,資源同質情況下6種沉水植物的間隔子長度有顯著差異,間隔子長度排列為穿葉眼子菜>扭葉眼子菜>竹葉眼子菜>苦草>光葉眼子菜>眼子菜。有研究表明,分株之間不僅通過間隔子交換能量和資源,同時克隆分株也靠間隔子來進行選擇性放置[33,34],所以間隔子的放置位置也直接決定了種群的空間擴張情況。可以推測,苦草的間隔子長度變異較低且轉向能力較弱,所以隨著間隔子長度的增加,種群擴張面積也隨之呈現線性增長;但眼子菜屬物種間隔子上“節”的出現使匍匐莖具備轉向能力,極可能出現匍匐莖“回頭”,長度增加但擴張面積不變的情況。
對比6種沉水植物的分株形成速率,苦草的分株形成速率最高,其均值為1.09株/d,最大值為1.84株/d,但是,其株高增加速率卻是最低的,僅有0.2 cm/d。這也與上述結論一致,說明苦草將更多的資源分配給形成新的分株來進行種群擴張,更快地占據平面空間資源。對于眼子菜屬物種,總體來說在克隆生長方面的資源分配策略表現為向分株株高和生物量增長投入更多比例,雖然分株形成速度和平面空間占據速度相對較慢,但占據垂向空間資源的速度較快。但是,眼子菜屬中的穿葉眼子菜和扭葉眼子菜卻表現出較強的分株形成能力,這兩個物種以較長的匍匐莖占據平面空間,維持種群的增長。
4 結論
本研究采用了同質園實驗的研究方法,在排除了自然條件下廣泛存在的資源異質性和干擾隨機性的影響前提下,研究了6種沉水植物在淺水環境的克隆生長過程中自身生物學特性的差異,結果表明,苦草株高最矮但分株數最多,面積擴張速率為35.48 cm2/d,傾向于產生更多的分株來形成密集型的種群。眼子菜屬的物種基于匍匐莖上“節”的存在,理論上在空間擴張方面更具優勢,但實際上竹葉眼子菜、眼子菜和光葉眼子菜的面積擴張速率卻低于苦草,因為這三個物種傾向于將資源分配給分株來保證自身的生長。而穿葉眼子菜和扭葉眼子菜的擴張模式則更好地體現了“節”的作用,通過更長的間隔子占據更多空間。因此我們有理由推測,如果不考慮種間相互作用,在空間廣闊的生境中,苦草、穿葉眼子菜、扭葉眼子菜更容易搶占空間資源,從而形成奠基者效應。基于本研究種間特異性參數,可以為構建不同沉水植物克隆生長模型以及種群動態模型提供參考數據,還能對沉水植被修復工程提供理論依據。這些沉水植物的克隆特性以及在異質生境中的不同響應策略在以后的研究工作中是值得進一步探討的。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
