液相色譜-串聯質譜法定量分析萊茵衣藻1,3-二油酸-2-棕櫚酸甘油三酯
2022-08-04 15:15:52魯媛晨李艷華余麗華韓丹翔馬海燕
水生生物學報 2022年7期
魯媛晨 李艷華 余麗華 韓丹翔 馬海燕
(1.中國科學院水生生物研究所,武漢 430072;2.中國科學院大學,北京 100049;3.北京大學工學院藻類生物技術創新實驗室,北京 100871)
1,3-二油酸-2-棕櫚酸甘油三酯(1,3-Dioleoyl-2-palmitoylglycerol,OPO)是母乳脂肪中含量最豐富的甘油三酯(Triacylglycerol,TAG)[1]。大量研究證明,TAG的立體化學結構即脂肪酸在甘油骨架上的分布會影響其吸收代謝[2,3]。人乳中這種獨特的sn-2位為棕櫚酸(C16∶0),sn-1/3位為油酸(C18∶1)的甘油三酯在嬰兒的成長期間起著重要的作用,可以促進脂肪酸和鈣質的有效吸收,保證嬰兒的能量攝入,并且可以改善嬰兒便秘,增加嬰兒骨骼強度[4—7]。2008年,中華人民共和國衛生部公告批準OPO可以作為營養強化劑應用于嬰兒配方奶粉中[8],預計未來我國嬰幼兒奶粉市場對OPO純品的年需求量可達3.6—13×107kg。
然而,目前國內市面上的商業化OPO大多依賴進口,并且是由一步酸解反應或兩步醇解酯化反應酶法合成的[9]。酶催化的過程提高了工藝的成本和復雜性,價格與質量之間的矛盾限制了OPO的應用程度[10]。因此亟待開發出可以規模化生產的天然OPO產品,如尋找新的富含OPO的油脂來源以滿足市場需求,同時解決依賴進口的問題。
萊茵衣藻(Chlamydomonas reinhardtii)是研究脂質代謝的模式生物,在環境脅迫的條件下可以積累大量的TAG作為儲存脂質[11]。分析發現其總脂肪酸組成絕大部分為C16和C18脂肪酸[12]。此前,有研究者發現,在缺氮脅迫條件下萊茵衣藻細胞中90%以上的TAGsn-2位是C16脂肪酸,且60%以上為C16∶0,而sn-1/3位上C16∶0和C18∶1的占比也接近30%[13]。在萊茵衣藻中存在兩條TAG從頭合成的肯尼迪途徑,分別定位于內質網和質體,其中溶血性磷脂酸酰基轉移酶(Lysophosphatidic acid Acyltransferase,LPAAT)是負責TAG中sn-2位酰基化的酶。當在萊茵衣藻中過表達萊茵衣藻質體定位的CrLPAAT1時,sn-2棕櫚酸酯的含量明顯提高[14];而內質網定位的CrLPAAT2也更加偏好C16∶0-CoA作為供體[15]。以上研究暗示萊茵衣藻中可能存在天然合成OPO的通路。
由于細胞內甘油三酯的組成非常復雜,針對特定結構TAG的檢測一直是研究的難點,目前國內還未出臺有關OPO的標準檢測方法。現有的針對OPO的檢測方法主要通過氣相色譜、高效液相色譜和氣相色譜-質譜聯用等。其中氣相色譜法和氣相色譜-質譜聯用法無法將OPO和它的同分異構體OOP完全分開[16,17]。銀離子色譜柱液相色譜法雖然可以使TAG同分異構體受到不同的束縛力分離開,但穩定性差,柱子使用壽命短[18]。而液相色譜-質譜聯用具有更高的靈敏度和精確度,且不需對樣品進行前處理,在特定TAG分析上更具優勢。超高效液相色譜(Ultra-High Performance Liquid Chromatography,UPLC)可以極大地提高色譜分離能力并縮短分析時間;質譜(Mass Spectrometer,MS)僅利用特征碎片離子就可以進行分子結構分析,其中三重四級桿分析器(Triple Quadrupole System,TQS)是進行單一m/z掃描最靈敏的儀器[19];電噴霧電離(ESI)離子源在脂質分析中早有應用,技術成熟。UPLC與MS聯用儀可以對藻油樣品中的OPO進行高效分析。
本研究首先建立了基于超高效液相色譜串聯三重四級桿質譜(UPLC-MS/MS)的OPO分析方法,并對缺氮脅迫條件下一株野生型萊茵衣藻cc-5325及其半乳糖基水解酶基因敲降突變體M08的OPO進行了定量分析,為進一步研究萊茵衣藻中OPO的生物合成提供了技術支持,也為產業化OPO生產提供了思路和方向。
1 材料與方法
1.1 萊茵衣藻藻種與培養方法
萊茵衣藻C.reinhardtiicc-5325(cw15mt+)和萊茵衣藻半乳糖基水解酶(Galactosyl Hydrolase,CrGH)基因插入突變株LMJ.RY0402.088610均購自美國明尼蘇達大學衣藻資源中心(ChlamydomonasResource Center,https://www.chlamycollection.org/)。為方便描述,上述藻株分別簡稱為cc-5325和M08。Crgh基因插入突變株M08的外源基因插入位點位于該基因內含子,且在缺氮條件下,CrGH蛋白表達水平顯著降低[20]。
從TAP固體平板上挑取單藻落接種于20 mL新鮮TAP液體培養基中,在光照強度40 μmol/(m2·s)、溫度25℃和轉速140 r/min下培養至對數生長期。以起始濃度2.0×105cells/mL轉接至200 mL TAP液體培養基培養,3d后更換為缺氮培養條件。以1000×g離心力離心藻液5min收集藻細胞,用缺氮TAP培養基清洗藻泥1次,重復離心后重懸藻泥至對應體積的缺氮TAP培養基中,在同樣條件下繼續培養3d。從更換缺氮培養基開始,每天取樣進行細胞計數、干重測量、定量PCR檢測及OPO含量分析。本實驗設置3個生物學平行。
1.2 生長曲線和生物質測定
每天固定時刻取藻液1 mL,添加魯哥氏碘液(終濃度1%)固定藻細胞。使用血球計數板在普通光學顯微鏡下計數,計算藻細胞密度。每份樣品計數4—6次并計算平均值。
藻細胞的干重測定參考Yin等[21]的105℃恒重法。在測定細胞干重前,將玻璃微纖維濾膜(Whatman GF/C膜)編號,于105℃過夜烘干,置于真空干燥容器中冷卻,稱量記錄膜重M1。每天固定時刻取藻液V=10 mL,使用GF/C膜抽濾去除培養基并用1 mol/L碳酸氫銨溶液清洗藻細胞1次。抽濾后的GF/C 膜移至105℃過夜干燥,次日置于真空干燥容器中冷卻,稱量帶有烘干藻細胞的濾膜并記錄總重M2,計算藻細胞生物量DW(g/mL)=(M2-M1)/V。
1.3 實驗試劑和標準品
三氯甲烷(色譜級)、乙酸銨(質譜級)和甲酸(質譜級)購自美國Sigma-Aldrich公司;甲醇(質譜級)、乙腈(質譜級)和異丙醇(質譜級)購自德國Merck公司。色譜級試劑用于總脂提取,質譜級試劑用于液質聯用儀分析。
外標標準品OPO和內標標準品十七烷酸甘油三酯(C17:0/C17:0/C17:0)標準品均購自美國Sigma-Aldrich公司。
1.4 萊茵衣藻Crgh基因轉錄水平檢測
RNA提取方法參照TransZolUp Plus RNA Kit試劑盒(ER501-01,TransGen Biotech)說明書。cDNA合成參照TransScript?All-in-One First-Strand cDNA Synthesis SuperMix for qPCR試劑盒(AT341-01,TransGen Biotech)說明書。
采用相對定量法檢測Crgh基因的轉錄水平,內參基因選擇α-Tubulin。熒光定量使用的Crgh引物和內參基因引物為:Crgh-F 5′-ACTGGTGGGG CATCAACTACTAC-3′/Crgh-R 5′-GTCTCAGTGA TGTACATGGGAATG-3′;α-Tubulin-F 5′-CTCGCT TCGCTTTGACGGTG-3′/α-Tubulin-R 5′-CGTGG TACGCCTTCTCGGC-3′。
qPCR擴增體系包括5 μL 2 × ChamQ Univeral SYBR qPCR Green Master Mix(Q711,Vazyme),10 μmol/L引物各0.2 μL,100 ng稀釋后的cDNA模板,加入ddH2O至反應體系為10 μL。qPCR反應條件為:95℃,600s;95℃,15s;60℃,30s;72℃,30s;40 cycles;95℃,60s;40℃,60s;65℃,1s;97℃,1s;37℃,30s。
1.5 總脂提取
總脂提取參考Yoon等[22]和Guo等[23]的方法并在此基礎上改進,在收獲的藻泥中加入4 mL甲醇∶氯仿∶甲酸(20∶10∶1,v/v/v),室溫下劇烈振蕩1h;加入2 mL的0.2 mol/L H3PO4與1 mol/L KCl混合溶液,室溫下劇烈振蕩5min;1000×g離心力離心10min后,用巴斯德玻璃吸管吸取下層有機層轉移至新的玻璃樣品瓶中;另外吸取1 mL氯仿加至提取管中,重復前兩步操作萃取出殘留的油脂;氮氣吹除有機溶劑,藻油凍存于-20℃。
1.6 標準溶液配制和標準曲線建立
將OPO標準品和C17∶0甘油三酯標準品分別用氯仿溶解,配制成終濃度為2 mg/mL的OPO標準溶液和20 μg/mL的C17∶0甘油三酯標準溶液,保存于-20℃備用。
定量吸取2 mg/mL的OPO標準溶液,用氯仿:甲醇(1∶1,v/v)溶液稀釋成質量濃度為500 μg/mL的OPO溶液,然后由高至低逐級稀釋得到質量濃度梯度為250、125、62.5、31.25、15.625和7.8125 μg/mL的OPO系列標準工作液。每個濃度梯度的OPO標準工作液取195 μL并各加入5 μL 20 μg/mL的C17∶0甘油三酯,使內標質量濃度為0.5 μg/mL。繪制OPO峰面積/內標物峰面積——OPO濃度/內標物濃度的曲線,計算內標標準曲線方程及相關系數。
1.7 藻油樣品前處理
將提取的藻油復溶于氯仿∶甲醇(1∶1,v/v)溶液中,每10 mg 藻生物質對應1 mL 氯仿:甲醇溶液;經孔徑0.22 μm的有機相針式過濾器濾過濾;取195 μL藻油濾液,加入5 μL 20 μg/mL的C17∶0內標溶液,充分振蕩混勻后進行UPLC-MS/MS分析。
1.8 UPLC-MS/MS法定量分析OPO
儀器采用ACQUITY超高效液相色譜儀(美國Waters公司)串聯三重四級桿質譜(美國Waters公司)。向1 μg/mL的OPO標準溶液加入10 mmol/L乙酸銨,以10 μL/min流速直接進樣于質譜的ESI源,在正離子模式下,對OPO的母離子[M+NH4]+(m/z 876.80)進行子離子掃描。高純氮氣作為霧化和干燥氣體,流速為600 L/h,去溶劑溫度為300℃。毛細管電壓和錐孔電壓分別為3000和40 V,碰撞能量20 V。
色譜條件參考Li等[24]的方法并在此基礎上改進。色譜分離采用ACQUITY UPLC BEH C18色譜柱(2.1 mm×100 mm,1.7 μm,美國Waters公司);柱溫35℃;進樣量為1 μL;流速為0.2 mL/min;流動相:A相為甲醇∶乙腈∶水(19∶19∶2,v/v/v),B相為異丙醇,兩者皆含有0.1%的甲酸和10 mmol/L的乙酸銨。梯度洗脫程序見表 1。
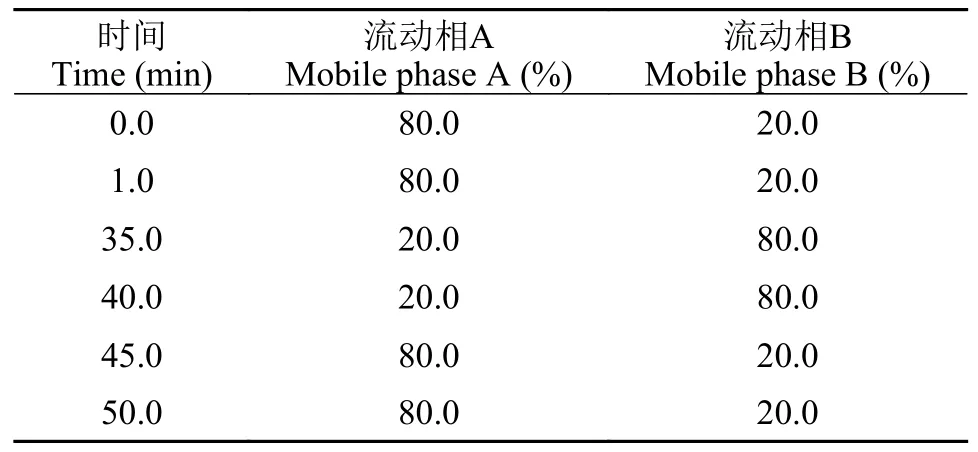
表1 UPLC梯度洗脫條件Tab.1 Gradient elution conditions of UPLC
質譜檢測的離子源為電噴霧離子源(Electrospray Ionization,ESI),在正離子模式下,采用多反應監測(Multiple Reaction Monitoring,MRM)模式,以高純氮氣作為霧化和干燥氣體,流速為800 L/h,去溶劑溫度為450℃。毛細管電壓和錐孔電壓分別為3000和40 V,碰撞能量20 V。
1.9 數據統計
采用MassLynx V 4.1軟件(Waters)進行數據采集和分析,采用Microsoft Excel 2010軟件進行數據處理和圖表制作,實驗結果以算術平均值±平均絕對偏差表示,采用Student’st-test對組間數據進行比較,P<0.05時,認為差異顯著。
2 結果
2.1 建立基于UPLC-MS/MS的OPO分析方法
在正離子模式下,對OPO標品溶液直接進樣進行全掃描,得到其準分子離子為[M+NH4]+,即母離子質荷比m/z為876。對[M+NH4]+(m/z 876)進行子離子掃描,如圖 1所示,碎片離子m/z 577.24 [M+NH4-299]+和m/z 603.28 [M+NH4-273]+分別為中性丟失299(C18H34O2+NH3)和中性丟失273(C16H32O2+NH3)的碎片,根據質荷比推算所丟失的碎片分別是sn-1或sn-3位上的油酸酰基和sn-2位上的棕櫚酸酰基相關碎片。碎片離子m/z 313.18 [R2CO+74]+和m/z 339.08 [RxCO+74]+(X=1,3)分別為丟失了sn-1,3位酰基和sn-1,2或sn-2,3位酰基的碎片。碎片離子m/z 264.99 [R2CO]+(X=1,3)和m/z 239.02 [R2CO]+分別為油酸酰基和棕櫚酸酰基相關碎片。
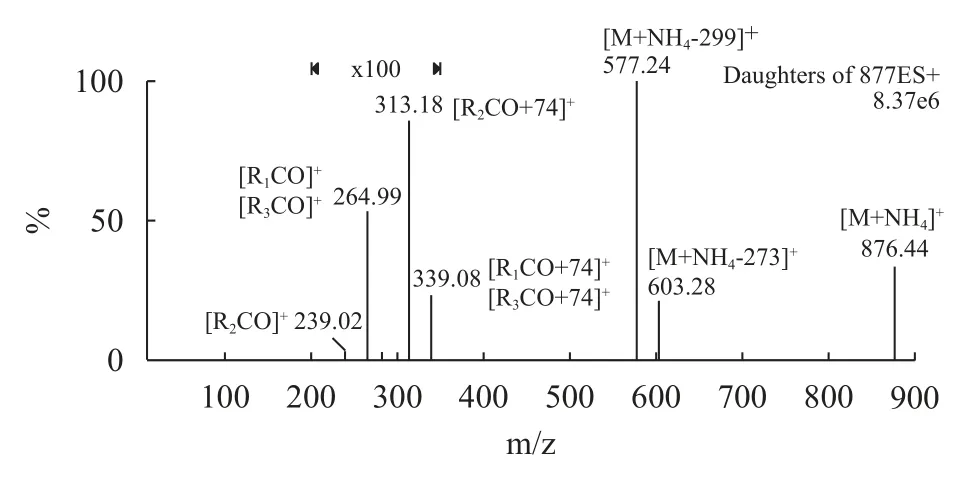
圖1 OPO子離子掃描質譜圖Fig.1 The product-ion scanning spectrum of the [M+NH4]+ ion of OPO
根據OPO的子離子掃描圖譜,本研究選擇了其中相對豐度最高的子離子m/z 577.24及其母離子m/z 876.44作為OPO的MRM定量離子對。考慮到色譜的分離效率和樣品中離子化效率,流動相中添加0.1%的甲酸和10 mmol/L的乙酸銨以獲得理想的分離度、保留時間和峰型。在優化的質譜和色譜條件下,以MRM模式檢測OPO標品的總離子流色譜圖如圖 2所示,保留時間為21.25min,保留時間窗口為0.6min。
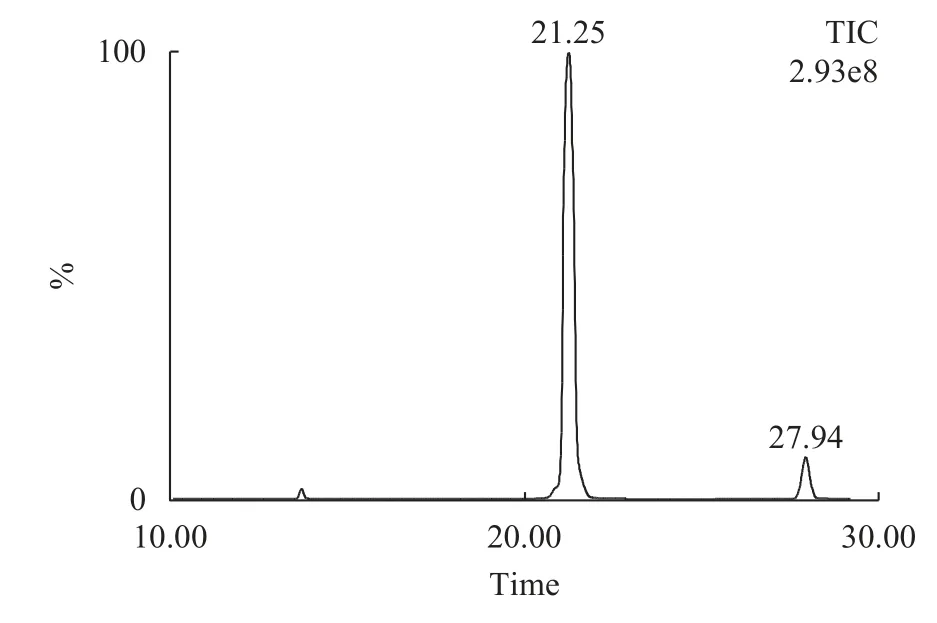
圖2 OPO總離子流圖Fig.2 Total ion chromatogram of OPO
2.2 OPO標準曲線方程
根據已知濃度的OPO標品溶液建立標準曲線。對1.5中的系列梯度標準溶液進行超高效液相色譜-串聯質譜測定,以OPO的濃度/內標物濃度為橫坐標(X,μg/mL),標準品OPO與內標C17:0的峰面積之比為縱坐標(Y),進行線性回歸分析,得到OPO的標準曲線方程為:Y=0.0083X+0.2248(相關系數R2=0.991),如圖 3所示,結果顯示OPO在7—250 μg/mL濃度范圍內線性關系良好,相關系數R2大于0.99,表明該方法有較高的靈敏度和準確度,因此可采用此方法對樣品中的OPO進行定量分析。
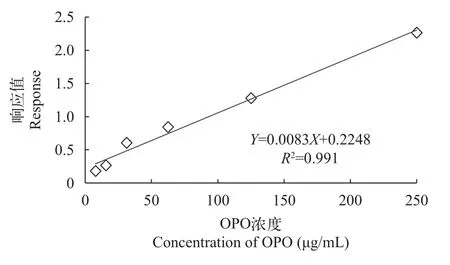
圖3 OPO的標準曲線方程Fig.3 Standard curves of OPO
2.3 萊茵衣藻cc-5325中OPO的檢測
通過上述研究建立的UPLC-MS/MS法對缺氮脅迫條件下的萊茵衣藻cc-5325細胞總脂粗提物中的OPO進行檢測,根據提取離子流圖中OPO峰強度可以判斷cc-5325在缺氮條件下的OPO積累情況(圖4)。結果顯示,在正常培養條件下(N0),細胞內僅積累極少量的OPO(圖4A),而在缺氮脅迫下(N1-N3)可以檢測到OPO的特征離子峰,且峰強度遠高于正常培養條件下OPO的峰強度,表明萊茵衣藻中的OPO在缺氮時大量積累。
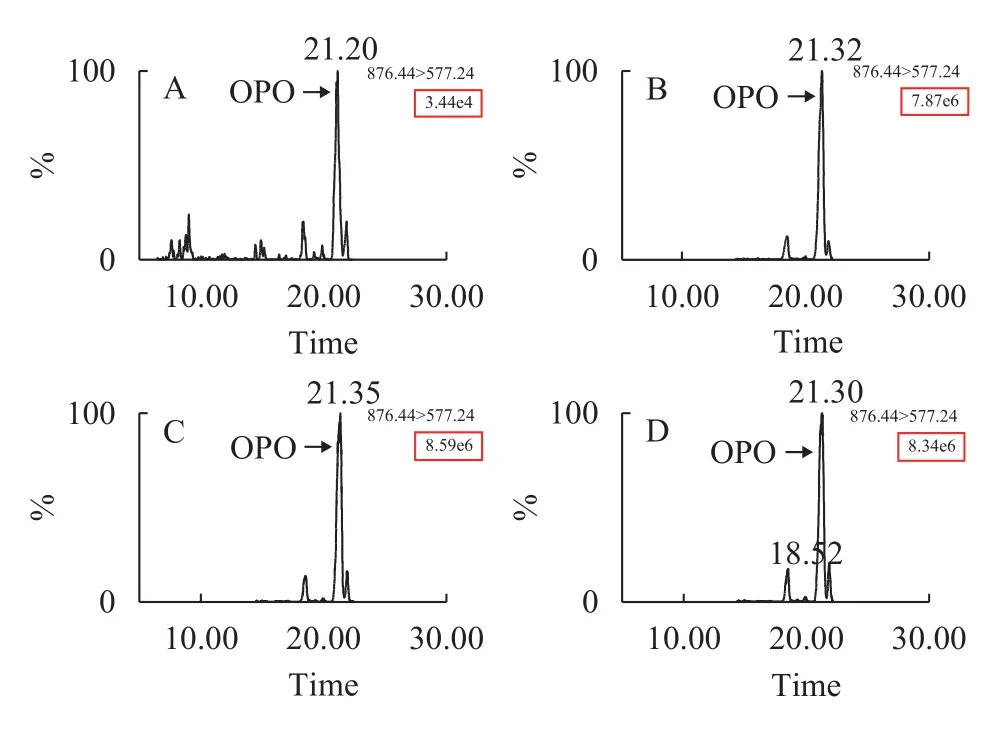
圖4 MRM檢測模式下特征離子掃描鑒定OPOFig.4 Identification of OPO by characteristic ion scanning detected by MRM
2.4 萊茵衣藻野生型和Crgh突變株的性狀表征
圖5為遺傳背景不同的萊茵衣藻野生型cc-5325和Crgh插入突變株M08的生長、生物質積累及Crgh基因表達性狀表征。在正常培養條件下,M08的Crgh基因轉錄水平相較于野生型下調了85.6%,說明插入突變對M08的Crgh基因表達造成了影響(圖5A),以上結果表明,突變株M08的Crgh基因在轉錄水平下調。
在缺氮脅迫條件下,萊茵衣藻細胞內會積累大量TAG。圖 5B和5C為野生型cc-5325和Crgh插入突變株M08在缺氮培養條件下的生長情況,結果顯示,兩者的生物量在缺氮培養期間持續積累,在缺氮第3天時達到最高,分別為0.60和0.38 mg/mL;兩者的細胞密度在缺氮期間不再大幅變化,在缺氮第3天時分別保持在0.85和1.41×105cells/mL左右。整體來看,插入突變株M08的生物量在整個培養期間都低于cc-5325,而細胞密度卻高于cc-5325,即cc-5325具有更高的單細胞干重。
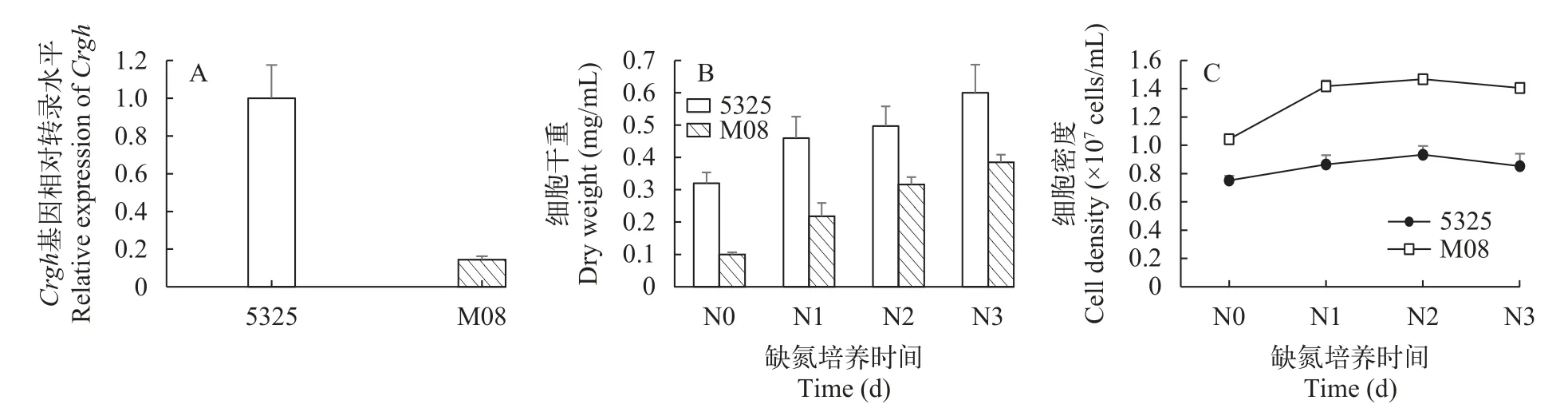
圖5 萊茵衣藻野生型cc-5325和Crgh突變株M08的性狀表征Fig.5 Characterization of wild-type cc-5325 and Crgh mutant strain M08
2.5 萊茵衣藻野生型和Crgh突變株的OPO含量表征
通過前述研究建立的UPLC-MS/MS方法對cc-5325和M08缺氮條件下的OPO含量進行定量分析。結果如圖 6所示,在缺氮脅迫第1至第3天,野生型cc-5325的OPO含量分別為0.82%、1.05%和0.96%,突變株M08的OPO含量分別為3.84%、4.22%和3.59%,兩者的OPO含量變化呈現出先升高后降低的趨勢,都在缺氮第2天達到了最高。M08的OPO含量始終高于野生型cc-5325,且在缺氮的第2和第3天達到了顯著(P<0.05),相較于野生型cc-5325分別提高了3.70、3.04和2.74倍。在缺氮脅迫第1至第3天,cc-5325的OPO產量分別達到了3.76、5.25和5.90 μg/mL,M08的OPO產量分別達到了8.01、13.26和13.73 μg/mL,兩者的OPO產量都在缺氮第3天時達到了最高。插入突變株M08的OPO產量也在整個缺氮脅迫期間顯著高于野生型cc-5325(P<0.05),相較于cc-5325分別提高了1.13、1.53和1.33倍。
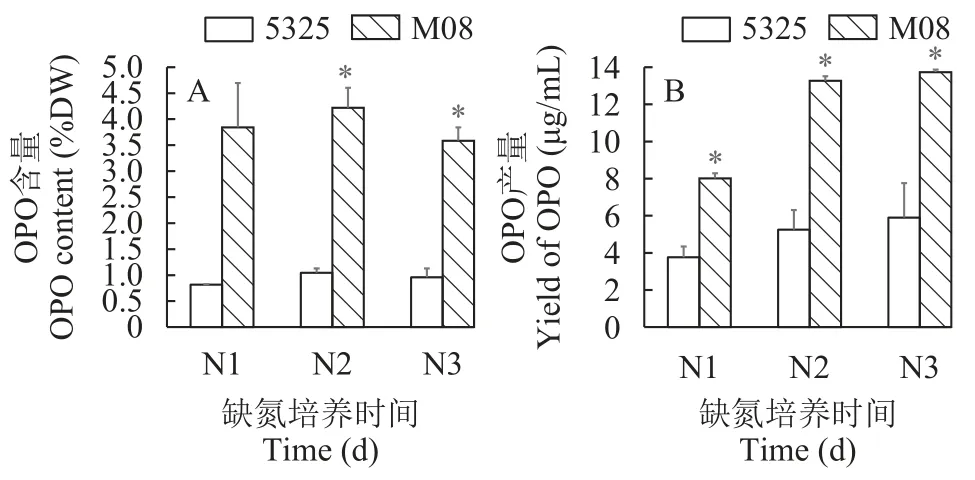
圖6 缺氮培養時萊茵衣藻野生型cc-5325和Crgh突變株M08的OPO積累(*P<0.05)Fig.6 The accumulation of OPO in wild-type cc-5325 and Crgh mutant strain M08 under nitrogen deprivation (*P<0.05)
3 討論
3.1 UPLC-MS/MS法定量定性OPO
本文通過優化高效液相色譜以及質譜條件,建立了適用于分析藻油中OPO含量的UPLC-MS/MS定量定性方法。結果表明,該方法可以有效地鑒定和定量分析細胞中的OPO,同時該方法也將適用于分析其他特定酰基組成的TAG。TAG是一種中性脂質,一般的電離技術很難讓TAG帶電荷,本研究采用的ESI離子源可以使TAG分子在離子對試劑的輔助下進行很好的電離[25],目前較常用的離子對試劑是金屬鋰化合物或鈉鹽,形成母離子加合物[M +Li]+或者[M+Na]+,但是金屬離子會污染質譜系統從而導致檢測器靈敏度降低。因此在本研究中,選擇了較理想的對質譜友好且易揮發的乙酸銨作為離子對試劑,形成母離子加合物[M+NH4]+,同時添加0.1%的甲酸改善分離度、保留時間和峰型。在子離子模式中,母離子[M+NH4]+經過碰撞可分解產生[M+NH4-RCOONH4]+(DAG+)和[RCO]+特征碎片離子,[M+NH4]+、DAG+和[RCO]+的信號對于TAG分子的定性和定量十分重要[26]。本文中所建立的方法正是基于此原理,在電子碰撞作用下,TAG會產生三種脂肪酸酰基碎片和三種DAG+特征碎片,分別為[R1CO]+、[R2CO]+、[R3CO]+和sn-1,2-DAG+、sn-2,3-DAG+、sn-1,3-DAG+,由于OPO的sn-1/3位上具有相同的脂肪酸組成,所以OPO會產生兩種脂肪酸酰基碎片和兩種DAG+特征碎片。甘油骨架上斷裂一個sn-2位脂肪酸所需要的能量要高于斷裂一個sn-1/3位的脂肪酸,因此sn-1,2-DAG+和sn-2,3-DAG+的比例要大于sn-1,3-DAG+,而sn-2脂肪酸所對應的[R2CO]+碎片離子的豐度值更低,通過這種方式可以判斷TAG上脂肪酸鏈的組成和位置,從而完成TAG的定性[27]。UPLC-MS/MS方法不需要對樣品進行衍生處理,具有重復性好,檢測方便快捷,精度高等優點。
本研究建立的方法對OPO標品進行了定性并建立了OPO的定量標準曲線(圖1— 3),結果顯示在5—250 μg/mL濃度范圍內OPO線性關系良好,相關系數可以達到0.99以上,表明本方法能夠準確地對樣品中的OPO含量進行定量。利用該方法對萊茵衣藻野生型cc-5325中提取的藻油進行了分析,發現總離子流圖的特征離子峰保留時間十分穩定(圖4),表明本研究方法的重現性較好、穩定性和可靠性高。缺氮條件下的萊茵衣藻細胞中可以檢測到天然OPO,外標法定量分析發現其含量可以達到干重的1.05%,這證實了萊茵衣藻作為OPO合成細胞工廠的應用潛力和商業化價值。該方法可以應用于研究不同遺傳改造的萊茵衣藻OPO合成情況的動態變化,從而為進一步開發OPO新油脂來源提供方向。
3.2 不同遺傳背景萊茵衣藻細胞內的OPO變化
萊茵衣藻在脅迫的環境條件下會積累大量的TAG,這個過程伴隨著光合膜脂(類囊體膜和葉綠體膜上的脂質)的降解[11]。不同遺傳背景的萊茵衣藻在生長速率、碳流分配等方面存在著差異,TAG合成通路及其關聯代謝途徑的基因的突變極有可能影響OPO的含量。本研究選擇了萊茵衣藻野生型cc-5325和Crgh缺陷突變株M08定量分析其細胞內的OPO合成情況,發現在由正常培養轉換為缺氮培養的過程中,兩株藻株的OPO含量都在初始階段急劇增加,隨后基本穩定,OPO產量在此過程中持續積累,而M08的OPO積累在缺氮條件下遠高于野生型cc-5325。研究發現,在脅迫條件下,萊茵衣藻Crgh基因表達和蛋白水平均有上調,半乳糖基水解酶介導了光合膜脂向TAG的轉化,由于Crgh的插入突變,M08的TAG積累會低于其野生型cc-5325[20]。而本研究發現M08的OPO含量和產量均較其野生型cc-5325有顯著提高,表明萊茵衣藻中OPO與總TAG的積累不是簡單地正相關,其背后還有一些復雜的未知機制。Crgh在高等植物如擬南芥中的同源基因AtSFR2可以利用糖脂合成二脂酰基甘油(Diacylglycerol,DAG)[28],DAG是肯尼迪途徑中合成TAG的前體產物。故推測Crgh在萊茵衣藻中也行使相同的功能,M08突變株中該基因的缺陷可能會導致缺氮條件下質體肯尼迪途徑TAG合成的降低。OPO含量的升高可能是因為其合成主要由位于內質網的真核肯尼迪途徑貢獻。但該假說還需要進一步實驗驗證。
通過遺傳改造萊茵衣藻油脂代謝途徑的某些相關基因,如Crgh,的確會對萊茵衣藻細胞內sn-2棕櫚酸酯如OPO的合成造成影響。此前,有研究者利用代謝工程手段對擬南芥(Arabidopsis thaliana)種子的油脂代謝途徑進行遺傳操作,在種子中過表達偏好C16∶0-CoA的歐洲油菜(Brassica napus)Bnlpaat1基因,隨后敲降了擬南芥真核途徑的原生Atlpaat2基因,并敲除了會使磷脂酰膽堿(phosphatidylcholine,PC)和甘油二酯(Diacylglycerol,DAG)互變的磷脂酰膽堿:二酰基甘油膽堿磷酸轉移酶(PC:DAG cholinephosphotransferase,PDCT)Atpdct基因,最終使得TAG中sn-2棕櫚酸酯含量提高了20多倍[29],這說明利用功能已知的靶基因去理性改造油脂代謝途徑可以實現特定立體化學結構的TAG(sn-2棕櫚酸酯)的大量積累,使其具有成為細胞工廠的潛力。萊茵衣藻質體來源的CrLPAAT1及內質網來源的CrLPAAT2對棕櫚酸的底物偏好性使得其作為OPO細胞工廠的優勢更加突出[14,15]。
本研究為定量分析微藻中特定結構的TAG提供了一種高效、準確和便捷的技術手段,并證實了萊茵衣藻作為OPO細胞工廠的可行性。作為一種安全級微生物,萊茵衣藻基底盤細胞的發展空間巨大,在脅迫條件下如何精準地調控微藻細胞內的碳流,將固碳網絡向目的產物的方向延伸,加強萊茵衣藻中OPO的積累是未來重點研究的方向。脂質合成途徑的關鍵酶基因作為潛在靶點,對其的進一步研究將幫助加速構建萊茵衣藻綠色生物制造平臺。
