嚙齒動物對旺業甸森林不同林木種子擴散格局的影響
2022-08-10 05:45:42查木哈烏云嘎吳琴馬成功白國棟邰峰張楠
生態環境學報 2022年6期
關鍵詞:差異
查木哈,烏云嘎,吳琴,馬成功,白國棟,邰峰,張楠
1.赤峰學院,內蒙古 赤峰 024000;2.阿拉善左旗草原工作站,內蒙古 巴彥浩特 750300;3.赤峰市防汛抗旱調度中心,內蒙古 赤峰 024000;4.內蒙古赤峰市喀喇沁旗旺業甸實驗林場,內蒙古 赤峰 024423;5.赤峰市森林草原保護發展中心,內蒙古 赤峰 024000
林木種子是嚙齒動物重要的食物來源。嚙齒動物同時作為種子的取食者和擴散者對林木種子進行取食和貯藏,貯藏的種子以備食物短缺期食用,但是貯藏的種子不能全部被找回,部分被遺忘在貯藏點,這些被遺忘的種子在合適的條件下,萌發成幼苗(Vander,1990)。所以,植物進化出不同種子特征,吸引動物取食的同時,又幫助自身完成擴散,在長期的進化過程中,動植物形成了一種互惠關系(Vander,1995;Forget et al.,1998;李宏俊等,2001)。嚙齒動物對不同種類的種子具有不同的行為反應及貯藏策略(Moles et al.,2003;Vander,2003;Jansen et al.,2004),特別是嚙齒動物對某些植物種子的選擇性貯藏,影響了這些植物種子的特征進化,如種子大小、種殼厚度及果實成熟期等,進而影響了這些植物種群的進化方向(Muoz et al.,2012)。種子大小的變化均在種子傳播、種子萌發和幼苗更新中起著重要的作用(L?nnberg et al.,2012;Sobral et al.,2014)。嚙齒動物對種子大小的行為反應及貯藏策略已有諸多研究,大部分研究支持大種子具有更高的擴散適合度(Xiao et al.,2005;Xiao et al.,2006;常罡等,2008;張博等,2016),而中等大小的種子在種子萌發及更新階段存活率更高(Cao et al.,2016)。種子的大小可能在種子命運中起著核心作用(Vander,2003;Xiao et al.,2004)。
林木種子成熟后嚙齒動物會對其進行捕食和貯藏。紅松(Pinuskoraiensis)、蒙古櫟(Quercus mongolica)、和平榛(Corylusheterophylla)均為旺業甸實驗林場廣泛分布的林木(李文臣等,2016),其種子為嚙齒動物的主要食物來源。種子大小與種皮特征直接影響嚙齒動物的擴散策略,本研究通過野外釋放3種同域分布的林木種子,調查嚙齒動物對不同特征種子的取食和擴散,探討嚙齒動物對旺業甸森林3種同域分布的林木種子擴散的影響及林木種子早期擴散過程中的影響,為森林天然更新提供科學依據。
1 材料與方法
1.1 試驗地概況
旺業甸實驗林場位于內蒙古赤峰市喀喇沁旗,屬燕山山脈北麓七老圖山支脈,118°09′—118°30′E,41°21′—41°39′N,是茅荊達壩次生林區重要組成部分。屬溫帶季風氣候,冬季漫長寒冷,夏季溫熱短暫,氣溫變化劇烈,早晚溫差大,日照時數在2700 h以上。有林地面積為23118 hm2,森林植被主要有油松(Pinustabulaeformis)、落葉松(Larixgmelinii)以及山楊(Populusdavidiana)、白樺(Betulaplatyphylla)、平榛、蒙古櫟、紅松等。林下草本植物主要有蒺藜草(Cenchrusechinatus)、罌粟葵(Callirhoeinvolucrata)、小苦荬(Ixeridiumchinense)、車前草(Plantago asiatica)、蔞蒿(Artemisiaselengensis)、瓜葉烏頭(Aconitumhemsleyanum)、蔓孩兒參(Pseudostellaria davidii)、匐枝毛茛(Ranunculusrepens)等。
1.2 種子特征
從種子釋放試驗備用種子中,每種隨機挑選100粒,分別測量種子的長度、寬度、種皮厚度及質量。
1.3 種子標記與種子釋放
2020年9月中旬種子成熟時,直接從母樹上采摘收集種子后晾干,篩選出完好的種子備用。采用塑料標簽標記法進行種子標記,該標記法目前在種子擴散實驗中已被廣泛使用,對鼠類的影響較小(常罡,2012)。每個標簽上記錄對應種子釋放站、種子類別及種子序號等編號信息,以便后續對種子進行追蹤和調查。小型嚙齒動物取食種子或進行搬運埋藏后,標簽通常會留在地面,通過搜尋標簽來確定每一粒種子的命運。2020年9月底在典型針闊混交林中設置3條平行的樣線,進行種子釋放。樣線間隔30 m,每條樣線上,間隔20 m設置一個種子釋放站,面積為1 m2,設置10個種子釋放站,3條樣線共設置釋放點30個。每個釋放點放置3種種子各10粒,成列均勻放置,共釋放種子900粒(3條樣線×10個釋放點×30粒)。
1.4 種子命運追蹤
種子釋放后的第1、3、5、7、10、15天進行調查,以種子釋放點半徑為35 m的范圍調查,并詳細記錄其種子去向:原地剩余、原地取食、移動后取食、移動后丟棄、移動后貯藏和丟失;調查記錄搬運后取食點,種子貯藏點插上彩色木棍,并標記種子編號跟蹤種子動態,測量其種子搬運距離。
1.5 嚙齒動物群落結構調查
種子釋放試驗結束后,對試驗區嚙齒動物群落結構進行了調查,設置5條樣線,每條樣線布置20籠,樣線隔20 m,籠距10 m,以生花生做誘餌,連續捕獲 3 d;對所有捕獲的嚙齒動物進行常規測量和記錄后原地釋放。隨機選兩個種子釋放站,放置紅外相機(東方紅鷹E3H)記錄嚙齒動物取食和擴散。捕獲率=捕獲數/有效籠數。
1.6 統計分析
采用SPSS 24.0軟件對相關數據進行統計分析。不同林木種子的存留曲線與不同命運的百分比數據經過反正弦轉換后進行統計分析。種子存留曲線由剩余種子百分比表示,采用Cox回歸法分析不同林木種子存留曲線的差異性。Cox回歸法分析又稱為比例風險回歸模型,該模型以生存結局和生存時間為因變量,可同時分析眾多因素對生存期的影響。通過一般線性模型分析不同類型種子在原地取食、移動后取食、移動后貯藏命運之間的差異。移動后丟棄和丟失種子數量少,不做單獨統計分析。采用單因素方差分析比較3種種子在貯藏距離和種子特征的差異,使用LSD法進行多重比較。在Origin 2018中完成作圖。
2 結果與分析
2.1 種子特征
3種林木種子特征值見表 1,其中平榛種子體積和質量最大,紅松種子的體積和質量最小;平榛種子的種殼厚度最大,紅松種子的種殼厚度最小。3種種子的質量(F=438.704,P<0.01)、直徑(F=628.728,P<0.01)、長度(F=327.274,P<0.01)、種皮厚度(F=102.263,P<0.01)均存在極顯著差異,采用LSD法進行多重比較表明,3種林木種子兩兩間均具有極顯著差異。
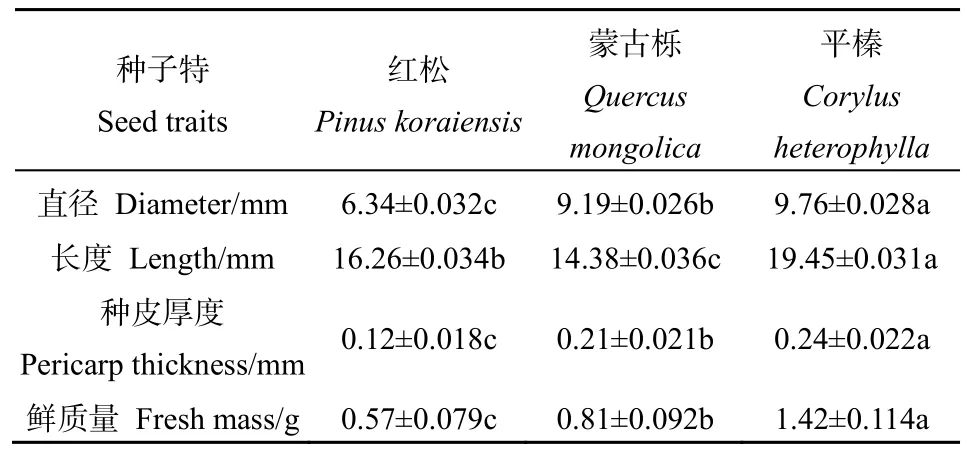
表1 3種林木種子特征Table 1 Seed characteristics of three forest trees
2.2 種子留存動態
3種林木種子原地留存動態見圖 1,Cox回歸分析結果表明,嚙齒動物對3種種子的留存曲線存在極顯著差異(Wald=12.880,df=2,P<0.01)。蒙古櫟種子和紅松種子存留時間較短,蒙古櫟種子在種子釋放10 d后全部被取食和擴散,紅松種子在種子釋放后15 d后均被取食和擴散,平榛種子試驗結束時原地剩余3.5%。
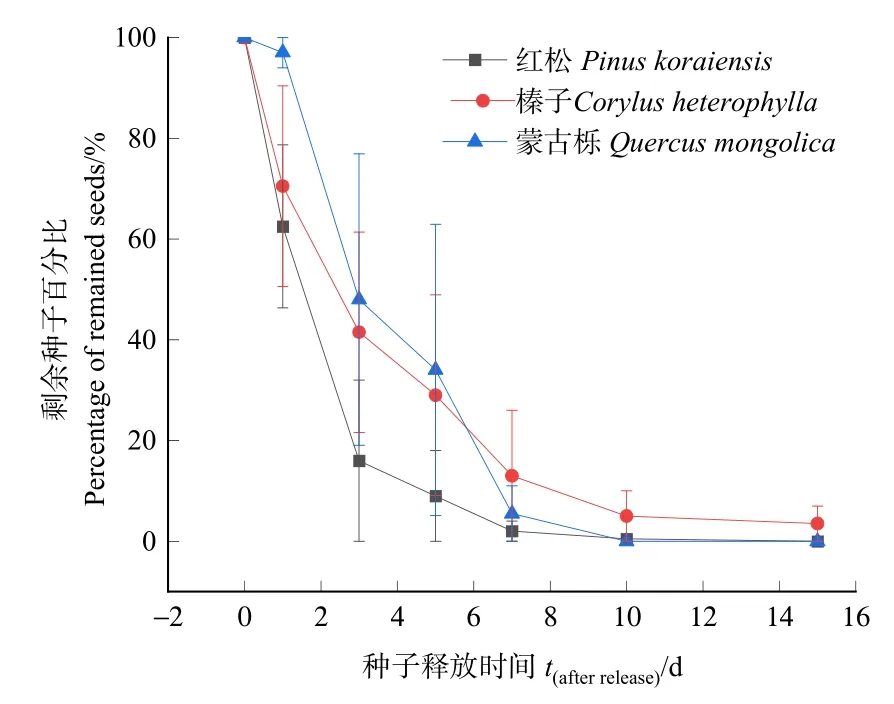
圖1 3種林木種子原地留存動態Figure 1 Dynamics of three kind of seeds that were remained in situ
2.3 種子命運
3種林木種子原地取食動態見圖2,3種林木種子原地取食動態一般線性模型分析結果表明,3種種子原地取食率存在顯著差異(F=4.039,P<0.05),紅松種子原地取食率最高,為65.5%,其次為蒙古櫟種子,原地取食率為39.0%,榛子原地取食率最低,為31.5%。
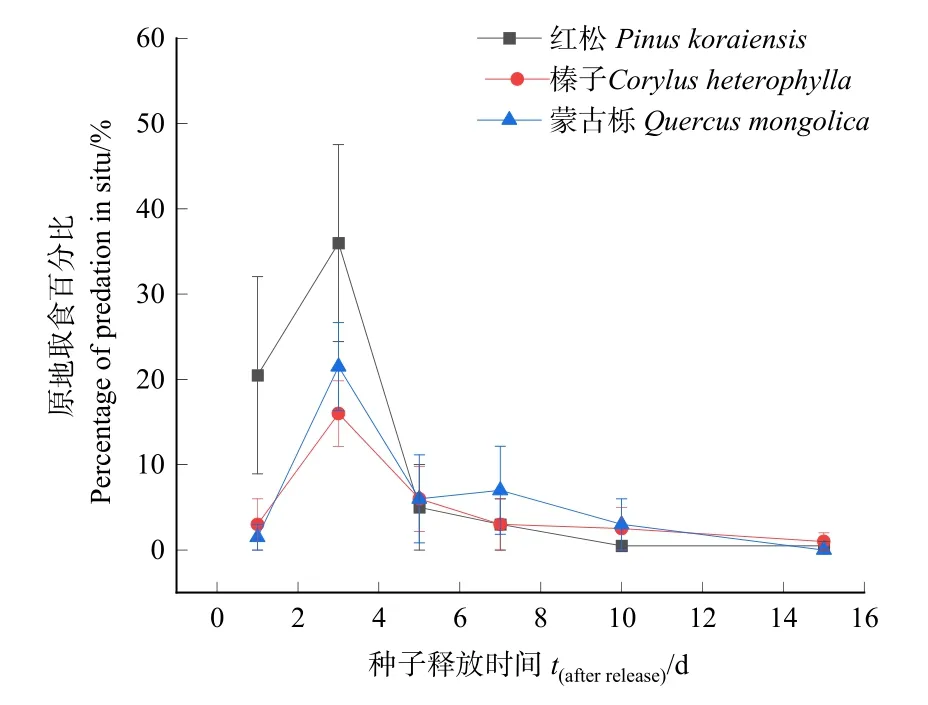
圖2 3種林木種子原地取食動態Figure 2 Dynamics of three kind of seeds that were eaten in situ
3種林木種子移動后取食動態見圖 3,紅松種子移動后取食率為14%,其次為榛子,移動后取食率為9%,蒙古櫟移動后取食率最低,為6%。一般線性模型分析結果表明,3種種子移動后取食率無顯著差異(F=3.293,P=0.052)。
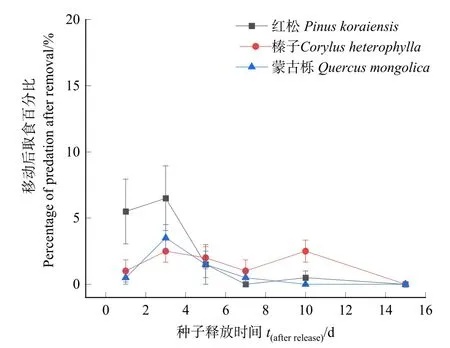
圖3 3種林木種子移動后取食動態Figure 3 Dynamics of three kind of seeds that were eaten after removal
3種林木種子移動后貯藏動態見圖 4,平榛種子移動后貯藏率為56%,高于其余2種種子,一般線性模型分析結果表明,3種種子移動后貯藏率具有顯著差異(F=4.954,P<0.05)。
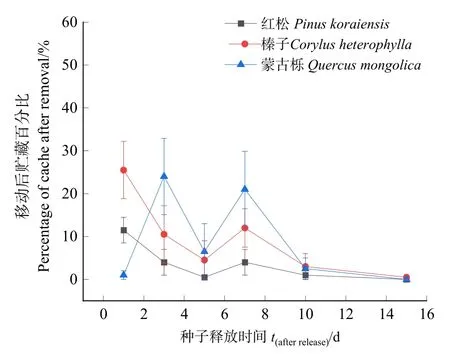
圖4 3種林木種子移動后貯藏動態Figure 4 Dynamics of three kind of seeds that were cached after removal
2.4 種子擴散距離
嚙齒動物移動后貯藏3種植物的種子的平均距離見圖 5,榛子移動后貯藏的平均距離最大,為12.78 m,蒙古櫟種子移動后貯藏的平均距離最小,為2.92 m,紅松種子移動后貯藏平均距離為5.82 m。LSD多重比較結果表明,榛子移動后貯藏的平均距離極顯著高于其他兩種種子移動后貯藏的平均距離(P<0.01),紅松與蒙古櫟種子移動后貯藏距離差異不顯著(P>0.05)。
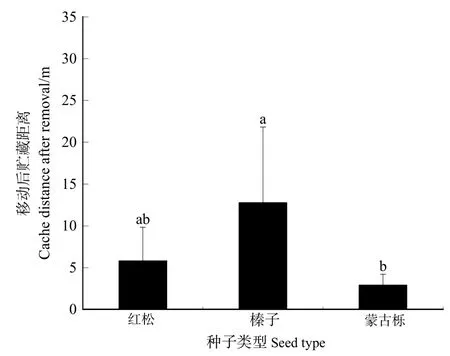
圖5 3種林木種子移動后貯藏距離Figure 5 Distance of three kind of seeds that were cached after removal
嚙齒動物移動后取食3種林木種子的平均距離見圖 6,紅松種子移動后取食的平均距離最大,為4.07 m,蒙古櫟種子移動后取食的平均距離最小,為2.83 m,榛子移動后取食平均距離為3.24 m。LSD多重比較結果表明,3種種子兩兩間均無顯著差異(F=0.815,P>0.05)。
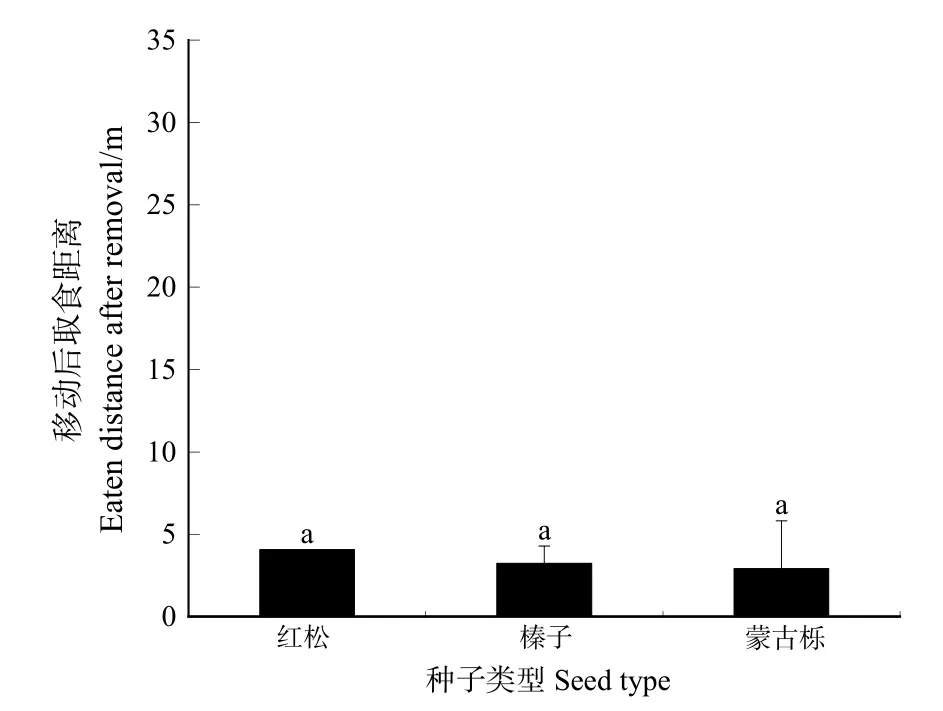
圖6 3種林木種子移動后取食距離Figure 6 Distance of three kind of seeds that were eaten after removal
2.5 嚙齒動物群落結構
試驗區共捕獲嚙齒動物3種,共16只(見表2),分別為大林姬鼠(Apodemuspeninsulae)、花鼠(Tamiassibiricus)和松鼠(Sciurusvulgaris),總捕獲率為5.33%,大林姬鼠捕獲率為3.67%,花鼠捕獲率為1.33%,松鼠捕獲率為0.33%。

表2 嚙齒動物群落結構及捕獲率Table2 Species of rodents and trap success rate in the study area
3 討論
嚙齒動物擴散植物種子的過程中,種子特征影響貯藏行為及擴散策略,從而影響種子命運(Xiao et al.,2006)。本研究調查嚙齒動物對紅松、蒙古櫟、平榛種子的取食和擴散,試驗結束時平榛種子仍有少量留存在原地未被取食和擴散,紅松種子取食和擴散時間早于其余兩種種子,原地取食率最高,榛子移動后貯藏率最高。嚙齒動物動物取食偏好種皮相對薄的種子,而貯藏種皮相對堅硬的種子(路紀琪等,2004),周立彪等(2013年)認為種皮特征比種子大小對嚙齒動物的覓食行為影響更大,與本研究結果相一致。3種林木種子中平榛種子種殼最厚,體積最大,質量最大,紅松種子的種殼最薄,體積最小,質量最小,蒙古櫟種子介于兩者之間。嚙齒動物處理較厚種殼種子的時間更久,增大被捕食風險(Dally et al.,2006),并且蒙古櫟種子易受昆蟲的侵害,不容易長期保存(王浩,2016),試驗中蒙古櫟種子被取食和擴散速度最快,表明嚙齒動物傾向于原地取食種皮相對較薄的小種子,將更多的具有堅硬種皮的大種子進行貯藏,嚙齒動物平衡能量消耗與獲益,中等的種子被取食和擴散的速度更快。這一試驗結果支持嚙齒動物傾向于原地取食小種子,移動后貯藏大種子(Moles et al.,2003;Xiao et al.,2006;Wang et al.,2012),中等大小的種子取食擴散速度最快。
嚙齒動物移動后貯藏3種植物種子的平均距離有所差異,平榛種子移動后貯藏的平均距離最大,達12.78 m,平榛種子貯藏距離最遠達 35m,主要集中在 10 m左右,導致平榛種子移動后貯藏距離誤差較大。牛可坤(2011)研究表明,在4種生境條件下平均擴散距離2.63—4.74 m,平榛平均擴散距離受不同生境的影響。平榛種子在不同的試驗中擴散距離差異較大,擴散距離受同域分布的其他種子、生境、種子豐富度、地勢等因素的影響。平榛種子移動后貯藏距離顯著高于其他兩種種子移動后貯藏的平均距離,紅松與蒙古櫟種子移動后貯藏距離差異不顯著。3種種子移動后取食的平均距離在兩兩間均無顯著差異。種子種類及大小對移動后取食的距離沒有顯著影響,而對移動后貯藏的距離有所影響。嚙齒動物為了平衡貯藏過程中的能量消耗與獲取,會盡可能將大種子搬運到更遠的位置貯藏,這一試驗結果支持最優貯藏模型(Dally et al.,2006)。
本研究中籠子僅捕到一只松鼠,但是紅外相機拍攝到種子站訪問頻率松鼠最高,松鼠比較靈敏,不易被籠子捕獲,活動距離相對較大,覓食范圍更廣。嚙齒動物對同一種種子具有不同程度的喜好(Chang et al.,2011)。嚙齒動物對種子的喜好程度影響種子命運,大林姬鼠從種子站將紅松種子搬運至洞穴集中貯藏后吃掉,這也是紅松種子取食擴散早于其他兩種種子的原因之一,但是集中貯藏種子不能成為潛在的種子庫,對林木的天然更新無積極的作用。
4 結論
本文研究結果驗證了:嚙齒動物傾向于原地取食小種子,將更多的大種子進行貯藏,貯藏的距離更遠。嚙齒動物平衡能量消耗與獲益,中等大小的種子被取食和擴散的速度更快。試驗結果支持中等大小的種子取食和擴散適合度更高。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
