PTEN 基因在周圍神經損傷修復中的作用機制研究進展
2022-08-16 08:37:24褚云峰于紅燕楊琪彭艷斌陳仲于
中國比較醫學雜志 2022年7期
關鍵詞:信號
褚云峰于紅燕楊 琪彭艷斌陳 仲于 斐
(1.北京大學深圳醫院,廣東 深圳 518036;2.濱州市濱城區市立醫院,山東 濱州 256600;3.骨科生物材料國家地方聯合工程研究中心,廣東 深圳 518036)
第10 號染色體缺失的磷酸酶及張力蛋白同源基因(phosphatase and tensin homologue deleted on chromosome 10,PTEN)是一種具有蛋白和脂質磷酸酶雙重活性的抑癌基因,與腫瘤發生關系密切,編碼隸屬于酪氨酸磷酸酶家族成員的蛋白質,通過調控細胞的增殖分裂影響腫瘤的進展[1]。 近年研究證實,PTEN基因在生物體周圍神經損傷修復中作用明顯[2],使其在骨科領域備受關注。
周圍神經損傷是一種由于解剖性原因和損傷性原因造成的周圍神經完整性變化的疾病,其致傷原因眾多,常見于體力勞動者,可造成嚴重的功能障礙,甚至殘疾,給家庭和社會帶來沉重的負擔,是骨科領域的棘手問題,也是我國健康領域亟待解決的重大公共衛生問題。 周圍神經損傷后,近遠端神經纖維將發生Wallerian 變性,隨時間的推移分解成小碎片,并逐步被吞噬細胞清除,神經修復后要經過變性、再生、跨越神經縫合口及終末器官生長成熟的過程,才能逐步恢復其支配功能,因此造成其臨床治療效果不佳。 國內外學者證實,PTEN基因與周圍神經損傷修復關系密切,可通過NF-κB[3]、PI3K/AKT/mTOR[4]、Jak/stat[5]、MAPK[6]等信號通路中因子的改變影響周圍神經損傷修復的進程。本文就PTEN基因在周圍神經損傷修復中的作用機制做一綜述,為相關研究提供便利。
1 PTEN 基因分子結構
PTEN是一個由9 個外顯子和8 個內含子組成的定位于染色體10q23.3 的200 kb 的基因[7],又被稱為MMAC1(mutated in multiple advanced cancer 1)或TEP1(TGF-regulated and epithelial cell-enriched phosphatase),其可編碼酪氨酸磷酸酶家族成員相關的由403 個氨基酸組成的蛋白質,該蛋白質的分子量為58×103~60×103,由N 末端、C2 結構域及C 末端構成。PTEN基因最早發現是作為磷酸酯酶作用行使腫瘤抑制基因的功能而進行研究的,與細胞生長相關的信號通路關系密切,可通過調控細胞周期影響到細胞的增殖及分裂[8]。 當腫瘤細胞中PTEN基因的表達量改變時,腫瘤細胞的生長、增殖及侵襲能力均受到顯著的影響。PTEN基因可通過甲基化、基因突變等方式在膀胱癌[9]、乳腺癌[10]、子宮內膜癌[11]等腫瘤疾病中起到重要的作用。
2 PTEN 基因與周圍神經損傷修復
研究顯示,PTEN基因也在骨科相關疾病中起到重要作用,如周圍神經損傷[12]、骨關節炎[13]、骨腫瘤[14]等。 由于其在周圍神經損傷后的突觸再生[15]、炎性及免疫反應[16]、神經元凋亡[17]、神經營養相關因子調控[18]等方面起到重要的作用而備受關注,成為該領域中的關注焦點。
2.1 PTEN 基因對周圍神經損傷后修復過程中突觸再生的影響
PTEN基因在周圍神經損傷修復中可以通過其翻譯后蛋白表達量的降低而促進軸突及髓鞘的再生[19]。 Singh 等[15]研究證實,在糖尿病周圍神經病變小鼠模型中,PTEN 蛋白上調可以抑制軸突再生,而體外培養這些小鼠的感覺神經元,可因PTEN基因敲除導致神經軸突生長和分支顯著增加。 Sun等[20]則證實,PTEN和細胞因子信號轉導抑制因子3(SOCS3)的同時缺失顯著地促進了軸突的穩健和持續再生,而其中激活mTOR 和STAT3 通路是維持成人中樞神經系統長距離軸突再生的關鍵,是功能恢復的關鍵步驟,這在神經損傷修復中起到重要作用。 Kar 等[21]實驗發現,損傷誘導的miR-21 和miR-199a-3p 表達的改變可以通過調節PTEN/mTOR 途徑改變整體和軸突內蛋白質合成,從而改變軸突生長能力。 Ohtake 等[22]綜述他人研究發現,通過PTEN條件敲除小鼠模型、短發夾RNA 敲除PTEN或通過藥理學方法阻斷PTEN基因作用,能夠刺激具有中樞神經系統損傷的幼年或成年嚙齒動物的不同程度軸突再生。 Liu 等[23]發現,脊髓損傷后,二十二碳六烯酸(DHA)顯著上調皮質脊髓神經元中的miR-21 并下調PTEN,在損傷后大鼠模型原代皮層神經元中,可以觀察到DHA 引起PTEN 下調及磷酸化AKT 的上調,與此伴隨神經軸突的生長。 Urban 等[24]則通過使用PTEN拮抗肽(PAP4)恢復了頸脊髓損傷后膈肌的功能,PAP4 能夠促進損傷呼吸神經元中頭端腹側呼吸組的軸突通過病變處長距離再生,并與膈肌運動神經元重新連接,從而影響膈肌呼吸功能恢復。 這些研究也從側面證實PTEN基因可以通過調控周圍神經損傷修復后的突觸再生影響神經功能恢復。
2.2 PTEN 基因對周圍神經損傷后修復過程中炎性及免疫反應的影響
PTEN基因在多種疾病中都能夠調控炎癥反應影響免疫進程。 在肝炎癥中,PTEN基因介導的蛋白激酶B/β-連環蛋白/叉頭框轉錄因子O 亞族1(Akt/β-catenin/Foxo1)信號通路在Toll 樣受體4(TLR4)相關先天性免疫調節中起到重要作用[25]。在免疫相關疾病類風濕性關節炎中,PTEN甲基化能夠促進成纖維細胞樣滑膜細胞中炎癥激活,造成關節軟骨的破壞[26]。 在中醫藥領域,補腎活血湯可以通過PI3K/AKT 途徑影響白介素-17A/白介素-17 受體(IL-17A/IL-17R)表達,導致PTEN mRNA上調,促進不明原因反復流產患者的蛻膜基質細胞增殖,這解釋了炎癥和凋亡關系及母胎界面Th17/Treg 免疫細胞之間動態平衡失衡機制[27]。
在周圍神經損傷修復過程中,炎性細胞引起的免疫反應也尤為重要。 Smith 等[28]發現,早期炎癥事件可能會抑制周圍神經系統和中樞神經系統損傷后的功能恢復,免疫相關因子腫瘤壞死因子介導周圍神經系統中的神經膠質細胞(schwann,又名雪旺細胞)中損傷誘導的NF-κB DNA 結合的快速激活,并且這些調控與損傷后軸突發芽的抑制有關。Schuld 等[29]認為,免疫蛋白酶體可被疾病、氧化應激及炎癥因子上調,在免疫蛋白酶體亞單位缺乏的視神經損傷小鼠模型中,位于炎癥經典通路NF-κB下游及PTEN基因上游的AKT 通路受到影響,參與視神經損傷修復進程。 Dodd 等[16]則認為,PTEN基因缺失能夠加快小鼠模型中惡性周圍神經鞘瘤這種影響機體免疫炎癥的形成,嵌合多肽阿霉素納米膠囊藥物制劑能夠使該模型中阿霉素濃度高于單純使用阿霉素給藥,這對于納米包載化療藥物治療類似腫瘤有重要的意義。 Kan 等[30]發現糖尿病神經病變中微血管病、血栓形成和炎癥浸潤明顯,伴隨缺氧誘導因子-1α(HIF-1α)和PTEN 的上調,而CD40 是引起兩者上調的關鍵分子之一。 由此可知,炎性反應和免疫反應在周圍神經損傷修復中有重要意義。
2.3 PTEN 基因對周圍神經損傷后修復過程中神經元凋亡的影響
神經元凋亡是周圍神經相關疾病中經常出現的一個進程,對其機制闡述有臨床意義。 周圍神經損傷通常會伴有缺血缺氧性損傷,尤其是當合并血管損傷時,可引起神經元凋亡,Cong 等[17]發現,皮膚前體來源的雪旺細胞的細胞外小泡,對暴露于氧糖剝奪條件下感覺神經元的存活和恢復意義重大,這一過程可能與miR-21-5p/PTEN/PI3K/Akt 軸有關。 Zhang 等[31]通過觀察Withaferin A 這種從催眠睡茄(Withaniasomnifera)中提取的甾體內酯對中風后神經元的作用,發現該物質可以抑制PTEN基因的表達,并通過激活PI3K/AKT 信號通路等作用抑制神經元的凋亡。 Song 等[32]通過實驗證實,PTEN磷酸酶代表了大鼠神經系統神經元存活和凋亡信號通路之間的一種新的轉化,PTEN 的抑制可以活化腦源性神經營養因子磷酸化AKT 的能力,并使基底前腦神經元免受前神經營養因子介導的凋亡。李叢琴等[33]認為,運動訓練可以通過抑制PTEN 通路的激活而減少海馬齒狀回區域凋亡蛋白的表達,減輕神經元凋亡并且減少大鼠抑郁程度。 林平等[34]則證實,神經生長因子過表達聯合PTEN基因敲減可以提升大鼠骨髓間充質干細胞的生存能力及分化為神經元樣細胞的能力,有利于神經元的再生從而提升周圍神經再生能力。 周坤等[35]則發現,甲氨蝶呤腹腔注射有利于急性脊髓損傷大鼠脊髓組織中PTEN 及衰老凋亡相關因子Bax 和Caspase-3的表達量降低,從而保護了神經組織,這樣也證實了PTEN基因下調對神經元的保護作用。
2.4 PTEN 基因對周圍神經損傷后修復過程中神經營養相關因子調控的影響
機體內有多種神經相關的營養因子,如神經生長因子( NGF)[36]、 腦源性神經營養因子(BDNF)[37]、睫狀神經生長因子(CTNF)[38]、神經營養因子3(NT-3)[39]、膠質細胞源性神經營養因子(GDNF)[40]等,這些神經營養相關的因子在周圍神經損傷修復中也起到一定的作用。 本課題組前期研究證實,大鼠坐骨神經鉗夾傷后的早期階段,CTNF 大致呈現出逐步上升的趨勢[38];當使用小分子多肽NP1 肌間隙單次注射治療坐骨神經鉗夾傷時,與損傷后注射生理鹽水組相比,β-NGF 的表達量明顯上調,周圍神經損傷也得到一定程度的修復[41]。 Sultan 等[42]將牙髓干細胞(DPSC)和三叉神經節神經元細胞(TGNC)共培養,發現DPSC 可以通過調控NGF、BDNF、NT-3 和GDNF 表達影響TGNC的存活和再生,在三叉神經無細胞修復中起重要作用。 Liu 等[43]通過生物信息學分析相關研究證實,PTEN 和BDNF 均可作為大鼠脊髓損傷后microRNA的靶點進行疾病的治療。 而且PTEN 抑制劑可以模擬人參皂苷Rg1 的作用上調海馬區BDNF 和ProBDNF 的表達,進而改善小鼠中神經相關的記憶問題[44]。 Cho 等[45]發現,脊髓損傷可導致膀胱功能障礙,此時相關組織中BDNF 和NGF 過度表達,使用PTEN 抑制劑后,膀胱功能改善,同時減弱了相關因子的過度表達。 陳歆維[18]發現,喉返神經損傷修復過程中PTEN基因起到負向調控作用,當敲減PTEN基因后,BDNF 和GDNF mRNA 的表達量上調,激活PI3K/Akt/GSK-3β 通路并實現喉返神經修復。 也由此可知,神經相關的營養因子能夠參與周圍神經損傷修復的進程。
3 PTEN 基因通過相關信號通路的作用對周圍神經損傷修復的影響
周圍神經損傷后的修復機制仍不清楚,只有對其機制了解清楚才能更好的治療該病。 在周圍神經損傷修復過程中,PTEN基因可以通過多條信號通路的作用影響損傷后的修復進程,如NF-κB[3]、PI3K/AKT/mTOR[4]、Jak/stat[5]、MAPK[6]等(表1)。
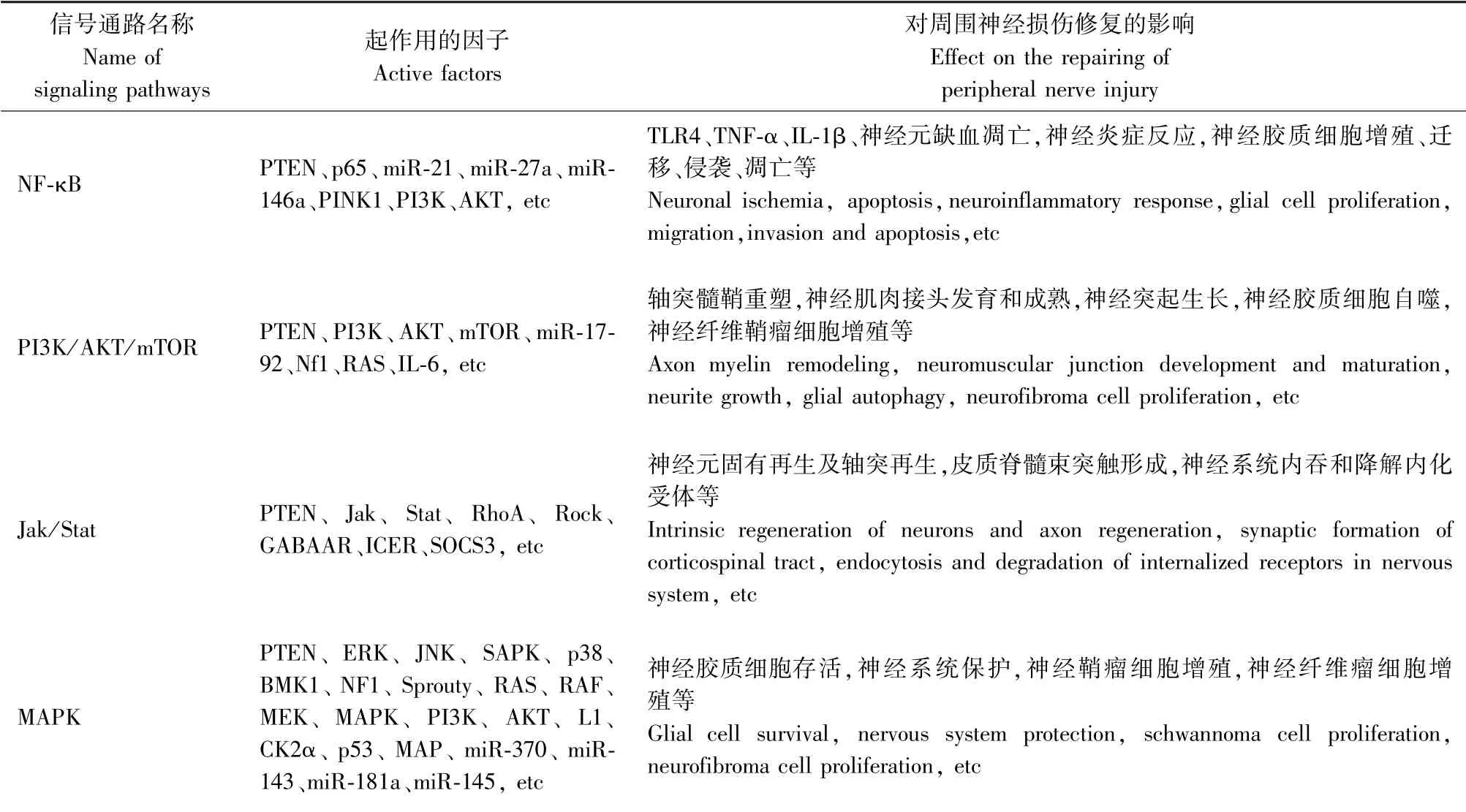
表1 PTEN 基因在周圍神經損傷修復過程中起作用的機制途徑Table 1 Mechanism and pathway of PTEN gene in the repair of peripheral nerve injury
3.1 PTEN 基因通過NF-κB 信號通路對周圍神經損傷修復的影響
NF-κB 信號通路是一條與炎癥關系密切的通路,參與機體的應激和損傷反應,其本身是蛋白因子轉錄家族,主要包括p50、p52、p65、RelB、c-Rel 5個亞單位。 由于周圍神經損傷修復過程中炎癥反應明顯,故該信號通路參與其中[46]。
Wang 等[3]發現,健康雪旺細胞的外顯體(SCExos)對糖尿病周圍神經病變具有治療作用,在這一過程中,SC-Exos 尾靜脈注射治療小鼠2 型糖尿病周圍神經病變時,逆轉了miR-21、27a、146a、PTEN及NF-κB 的表達,進而起到治療作用。 王尋等[47]證實,星形膠質細胞缺氧模型內PTEN誘導假定激酶1(PINK1)沉默后NF-κB 信號通路的活性明顯降低,使缺血神經元的凋亡升高,從而證實PINK1 介導缺氧損傷時星形膠質細胞對神經元具有保護作用。 趙敏[48]也認為,脂多糖處理過的海馬神經元細胞,神經元不是神經炎癥反應中的被動者,這一過程TLR4 介導的PTEN/PI3K/AKT/NF-κB 信號通路激活后,TNF-α 和IL-1β 等因子表達上調,最終參與了神經炎癥反應。 王澤夏[49]則發現,天然中藥小分子化合物姜黃素對神經膠質瘤的增殖、遷移、侵襲和凋亡具有影響,在細胞中,10 μmol/L 以上的姜黃素能夠抑制膠質母細胞瘤細胞株的增殖,在降低其遷移和侵襲能力的同時促進了細胞凋亡,在裸鼠皮下成瘤模型中也抑制了瘤的生長,這一過程中姜黃素抑制NF-κB 蛋白表達,但促進PTEN 蛋白的表達。 由此可知,NF-κB 信號通路介導的炎癥反應在PTEN基因的影響下參與周圍神經損傷修復的進程。
3.2 PTEN 基因通過PI3K/AKT/mTOR 信號通路對周圍神經損傷修復的影響
PI3K/AKT/mTOR 信號通路在細胞增殖、分化、凋亡、葡萄糖轉運中起到重要的作用,該通路與PTEN基因關系密切,參與多種神經相關疾病,如缺血性中風[50]、脊髓損傷[51]、帕金森病[52]等。
該通路在周圍神經損傷修復中作用明顯。 Xin等[4]發現,在大鼠中風模型尾靜脈注射miR-17-92慢病毒,可增加缺血邊界區神經軸突和髓鞘的密度,這種增強軸突髓鞘重塑的作用可能是通過PTEN下調誘導的PI3K/Akt/mTOR 通路激活實現的,從而證實MiR-17-92 簇能夠促進中風后功能的恢復。 Li 等[53]則證實,雪旺細胞中神經纖維蛋白1(Nf1)和PTEN的條件性失活導致神經肌肉接頭異常成熟,這一過程是由于RAS/PI3K/AKT/mTOR 信號通路過度活躍造成的,這一通路對神經肌肉接頭的發育及隨后的肌肉成熟意義重大。 Leibinger等[54]認為,高表達的IL-6 聯合視網膜神經節細胞特異性敲除PTEN基因可通過PI3K/AKT/mTOR 信號通路的激活促進神經突起生長,從而發現了治療視神經損傷的新方法。 彭湃等[55]實驗研究說明,右美托咪定聯合放療與單純放療相比,可以通過抑制PTEN/Akt/mTOR 增強神經膠質瘤細胞的自噬與放療敏感性,這一過程中AKT 和mTOR 的磷酸化水平降低,細胞凋亡率及PTEN 蛋白的水平升高。Bradtm?ller 等[56]則認為,PTEN 在惡性神經纖維鞘瘤組織中的表達量明顯低于神經纖維瘤和正常組織,使用mTOR 相關制劑與辛伐他汀合用,能夠起到協同抗細胞增殖的作用。
3.3 PTEN 基因通過Jak/Stat 信號通路對周圍神經損傷修復的影響
Jak/Stat 信號通路是一條由細胞因子刺激的信號轉導通路,參與免疫調節、細胞增殖分化及凋亡等生物學過程。 該通路同樣對神經相關疾病進程起到重要的作用,如慢性青光眼早期的視神經頭膠質細胞增殖[57]、坐骨神經瓦勒變性的損傷修復[58]等。
Lu 等[59]發現,PTEN/mTOR、Jak/Stat、DLK/JNK信號軸在哺乳動物成年成熟神經元的固有再生能力及軸突再生中起到重要作用。 Zareen 等[5]認為,皮質脊髓束受損時,慢性刺激將PTEN 轉變為非活性磷酸化形式,此時受損神經軸突向生長狀態過度,選擇性阻斷Jak/stat 信號通路并不消除刺激依賴性軸突發芽,而是阻止了慢性運動皮質刺激產生的皮質脊髓束突觸前位點增加,證實了Jak/Stat 信號通路在活動依賴性皮質脊髓束突觸形成中的重要作用。 Riffault 等[60]確認,RhoA-Rock-PTEN 途徑能夠降低神經系統中典型抑制性神經遞質GABA的A 型受體(GABAAR)的磷酸化水平,進而影響神經功能并觸發內吞和降解內化受體;JAK-STATICER 途徑則在這一過程中受到影響, 減少GABAAR 的合成。 Gallaher 等[61]則在小鼠模型中看到,PTEN基因缺失能夠增強坐骨神經損傷后軸突的再生,當合并SOCS3 雙基因缺失時,再生能力更強,而Jak/Stat 信號通路在坐骨神經損傷后早期階段內被激活,但損傷后7 d 時該通路恢復到基線水平。 因而我們可知道,PTEN基因能夠通過激活或抑制Jak/Stat 信號通路調控周圍神經損傷修復。
3.4 PTEN 基因通過MAPK 信號通路對周圍神經損傷修復的影響
MAPK 是一組進化保守的絲/蘇氨酸蛋白激酶,在哺乳動物中有四種類型的MAPK 級聯通路,研究最廣的是ERK1/ERK2 (p42/p44)激酶,此外JNK/SAPK、p38 和BMK1 均為其級聯通路。 該通路在細胞增殖、分化、轉化及凋亡等過程中至關重要[62]。
Zhang 等[6]通過酪氨酸激酶通路的活性程度,發現了成人彌漫性膠質瘤聚集的分子表征亞群,確定了與NF1、Sprouty和PTEN共表達的膠質瘤基因模塊,從而使WHO II-IV 級成人彌漫性膠質瘤能夠從5 個獨立數據集中聚集成兩個亞型,分別與RASRAF-MEK-MAPK 級聯和PI3K-AKT 通路相關,這項研究有助于解釋該類型膠質瘤患者的存活情況,并從微環境相關層面研究膠質瘤的進展。 Wang 等[63]則發現,右旋美托咪定這種α2 腎上腺素能受體激動劑能夠通過保護神經,起到治療衰老大鼠的認知功能障礙的作用,這一過程中右旋美托咪定能夠抑制異氟醚所致認知損害大鼠組織中PTEN 和p38 MAPK 蛋白的表達,并激活p-AKT 蛋白表達。 Farid等[64]則認為,惡性周圍神經鞘瘤這種罕見病中,PTEN功能喪失,MAPK 通路活性上調,從而為該類疾病患者的治療護理提供了理論基礎。 Wang 等[65]實驗證實,細胞粘附分子L1 通過激活CK2α 刺激神經細胞生成,這一過程導致PTEN 和p53 下調,同源src/MAP 激酶信號通路參與這一過程。 也有研究全面描述了叢狀神經纖維瘤中microRNA 的表達情況,發現叢狀神經纖維瘤中PTEN mRNA 表達降低,該病中參與RAS-MAPK 通路的四種miRNA(miR-370、miR-143、 miR-181a 和 miR-145) 出 現 異 常表達[66]。
4 結語與展望
當前的研究中,周圍神經損傷修復的機制并不明確,臨床醫生無法針對性的用藥促進其損傷后的修復進程,使該病的治療效果不佳。 研究證實,PTEN基因在周圍神經損傷修復中起到重要作用,它可以通過調控周圍神經損傷后的突觸再生、炎性及免疫反應、神經元凋亡、神經營養相關因子等過程以及NF-κB、PI3K/AKT/mTOR、Jak/stat、MAPK等信號通路的作用,影響周圍神經損傷后的修復。由于PTEN基因功能多樣且復雜,需要更多的研究論述其在周圍神經損傷修復中的作用機制,從而為臨床治療該病提供實驗依據。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
