紅樹林放線菌抗沃柑病原真菌的研究
2022-08-18 03:22:30李菲楊玲王巧貞許秀松黃庶識楊慧歡覃仙玲
中國抗生素雜志 2022年7期
李菲 楊玲 王巧貞 許秀松 黃庶識 楊慧歡 覃仙玲,*
(1 廣西科學院廣西近海海洋環境科學重點實驗室,南寧 530007;2 廣西科學院 廣西海洋天然產物與組合生物合成化學重點實驗室,南寧 530007;3 廣西師范大學 生命科學學院,桂林 541006;4 廣西師范大學 珍稀瀕危動植物生態與環境保護教育部重點實驗室,桂林 541006;5 南寧學院機電與質量技術工程學院,南寧 530200)
柑橘是我國重要的水果產業,品種多,分布廣,種植規模和產量均居全球第一[1]。沃柑系以色列研發的雜交柑橘品種,由于晚熟高糖和早結豐產等優點,在我國廣受青睞[2-4]。沃柑晚熟,需要越冬,但又不防凍,因此適合我國廣西等積溫高的地區種植。截至2018年底,廣西沃柑種植面積約200萬畝(13.33萬hm2),產量約100萬噸,位居全國第一位[5]。沃柑作為廣西柑桔產業的支柱產品,對廣西的農業經濟有著重大意義。然而,柑橘培育過程中發生的病蟲害不容小覷。據美國植物病理學會網站記錄,柑橘真菌性病害60余種,其危害程度因地區和品種而異[6]。為順利開展沃柑病害中病原真菌的防控工作,本課題組在廣西南寧上林沃柑培育基地開展了沃柑病害的調查,結果發現沃柑枝葉及果實在生長過程中,會出現不同程度的真菌感染,病征與柑橘炭疽病、柑橘潰瘍病及柑橘黑點病的癥狀相似,為弄清病原真菌種類及尋找針對性的防治方法,本研究對沃柑葉斑病害中病原真菌進行分離和初步鑒定,并開展了紅樹林放線菌抗沃柑病原真菌的研究。
放線菌,尤其是鏈霉菌是天然抗菌活性產物的主要來源,約2/3的抗生素是由放線菌產生[7]。放線菌作為土壤中的優勢菌群,經過數十年來大規模的挖掘和篩選后,普通生態環境中挖掘出新穎的放線菌資源越來越困難,故人們將目光轉向了特殊生態環境。紅樹林作為海陸交界的特殊生態系統,其微生物群落在適應生態環境與自我保護中,通過通信、覓食、拮抗等互作的微生物生存機制,來適應謂之“生命極限”的特殊環境,其獨特的生境必然會造成其代謝出更具特點的化合物及其相應的生物活性[8]。研究發現,紅樹林土壤放線菌不僅產生多種功能酶[9],而且還分泌具有抗菌、抗腫瘤、抗氧化等藥用活性的代謝產物[9-12]。本研究以分離到的植物病原真菌為指示菌,從紅樹林放線菌中篩選出了具有顯著抑菌活性的菌株,為沃柑葉斑病害的防控奠定了基礎。
1 材料與方法
1.1 試驗材料
1.1.1 供試樣本
(1)病害樣本于2021年10月采集自廣西農業科學院園藝研究所的沃柑培育基地,采集有明顯病癥部位的葉片樣品,裝于密封袋帶回實驗室進行研究。
(2)健康葉片樣本采自廣西農業科學院科研基地,選取形態相近且健康的沃柑帶葉枝條若干,放置于密封袋保存帶回實驗室備用。
1.1.2 主要試劑和儀器
(1)試劑 培養基原料、TAE緩沖液、2×EasyTaqMasterMix、引物(ITS1和ITS4)、DNA marker、GoldView核酸染料,Chelex 100樹脂(Bio-Rad),其他有機試劑(分析純)等購于北京康為世紀生物科技有限公司。
(2)儀器 SW-CJ-2F型超凈工作臺(杭州佳濾設備有限公司),HH·B11-BS-Ⅱ型恒溫培養箱(東莞市恒宇儀器有限公司),Tgradient型PCR擴增儀(德國Biometra),電泳儀(美國BioRad),Gel Logic 2200Pro凝膠成像儀(Carestream),VB-55型高壓滅菌鍋(德國Systec),MINI-6k型迷你離心機(常州市國旺儀器制造有限公司),MINIB-100型金屬浴(杭州米歐儀器有限公司),恒溫振蕩器(美國Crystal),N1000型旋轉蒸發儀(日本Eyela)和光學顯微鏡(日本Nikon)等。
1.1.3 培養基
真菌分離、純化及拮抗菌篩選培養基:PDA固體培養基(土豆200.0 g,葡萄糖20.0 g,去離子水1000 mL,瓊脂14.0 g,pH 6.8~7.2)。
發酵培養基:改良InternationalStreptomycesProject 2(ISP2)固體培養基[14]。
1.1.4 供試放線菌菌株
選取24株于2021年11月分離自廣西茅尾海紅樹林自然保護區內桐花樹根際土壤的放線菌,通過EzBioCloud數據庫(https://www.ezbiocloud.net/)在線比對[15],這些放線菌隸屬于12屬,其中5株鏈霉菌屬(Streptomyces),5株紅球菌屬(Rhodococcus),3株微桿菌屬 (Microbacterium),3株小單孢菌屬 (Micromonospora),2株類諾卡菌(Nocardioides),壤霉菌屬(Agromyces)、短狀桿菌屬(Brachybacterium)、戈登菌屬(Gordonia)、棒狀桿菌屬(Corynebacterium)、棲白蟻菌屬 (Isoptericola)、分枝桿菌屬(Mycobacterium)和Phycicoccus屬各1株;均保藏于廣西科學院北部灣海洋微生物種質資源庫中,具體信息見表1。
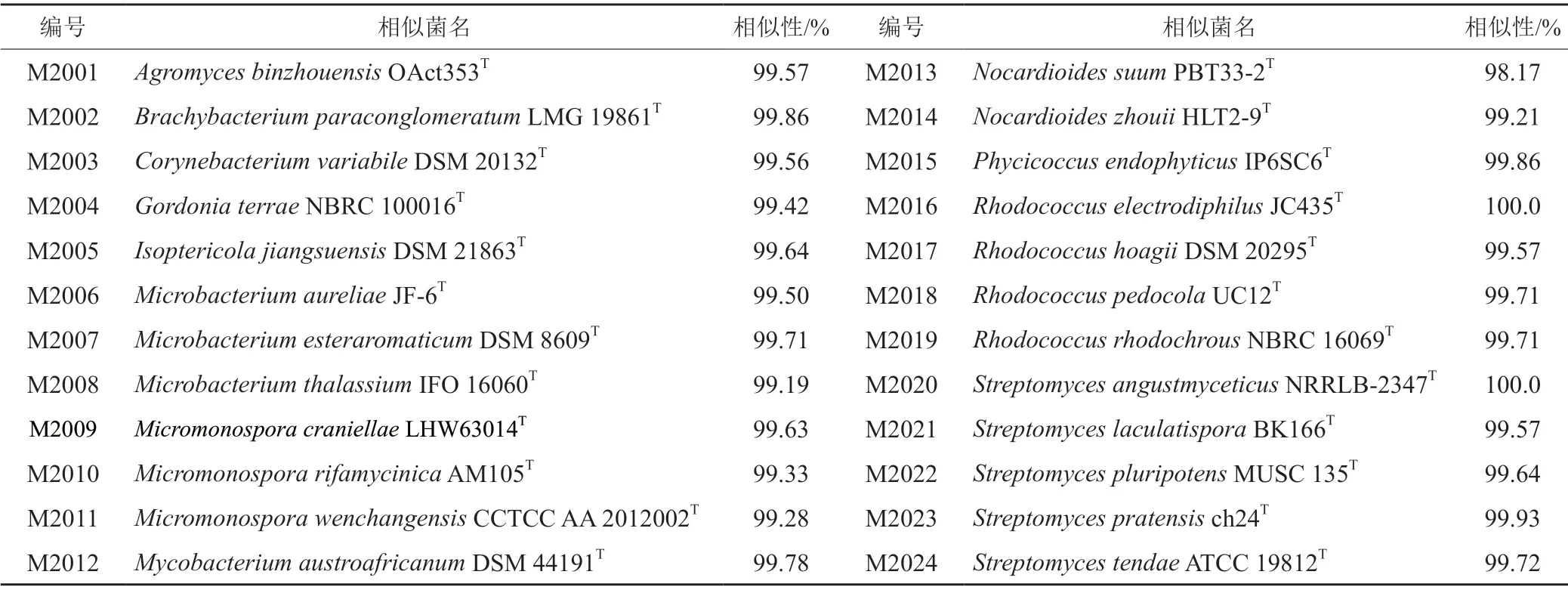
表1 拮抗實驗用紅樹林放線菌信息Tab.1 Actinomycete strains from mangrove for antagonistic assay
1.2 病原真菌分離與鑒定
1.2.1 菌株分離純化
用自來水沖洗患病葉片組織后,依次用75%乙醇和5%次氯酸鈉溶液浸泡組織各5 min,每次消毒后用滅菌水反復沖洗3次。處理后的病樣組織置于超凈工作臺晾干,用無菌剪刀將有病樣部位剪成小塊,分散鋪于PDA固體培養基上,28℃培養3~5 d。觀察平板上菌落生長情況,挑取肉眼可見菌落進行純化培養,并將純化好的菌株接種至PDA試管培養基中,待其生長至對數期后進行斜面保藏。
1.2.2 真菌生長速率測定及形態觀察
用打孔器(直徑6 mm)取新鮮的菌苔邊緣的菌絲塊轉接于PDA培養基(直徑90 mm)中央,置于28℃培養,每株菌3組平行。用十字交叉法測量菌落直徑,待菌絲長滿即停止測量,菌落平均生長速率單位為mm/d。挑取PDA表面的分生孢子,在光學顯微鏡下觀察其形態。
1.2.3 真菌菌株基因測序和系統發育分析
采用chelex-100樹脂[16]快速提取待測真菌的基因組DNA作為PCR模板,對待測菌進行PCR擴增。引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′),特異性擴增出各個菌株的ITS1-5.8S-ITS2區域,基因序列長度約為550 bp;擴增條件:94℃預變性8 min,94℃變性30 s,55℃退火30 s,72℃延伸2 min,共30個循環,循環結束后,72℃延伸10 min。擴增產物經1%瓊脂糖凝膠電泳檢測合格后,委托生工生物工程(上海)股份有限公司進行測序分析。序列經BioEdit Sequence Alignment Editor軟件整理后,測序結果在NCBI中進行BLAST同源性比對,分別獲得同源性相近的菌種序列,運用MEGA7.0軟件,采用Neighbor-Joining法構建系統樹,對各菌株的系統發育地位進行分析[17]。
1.2.4 致病性測定
參照趙洪濤等[18]的方法,按照柯赫氏法則,對分離到的疑似致病菌株進行回接驗證。具體操作如下,將新鮮葉子從枝條上剪下,用自來水進行表面沖洗后,再用75%乙醇溶液對其表面消毒3次,每次消毒后用無菌水沖洗3遍,置于超凈工作臺中晾干。用無菌牙簽在葉面上有序扎數個孔。用無菌生理鹽水制備真菌分生孢子液,懸浮液濃度約為1×107CFU/mL。滴加10 μL在扎孔處,對照組則滴加10 μL無菌生理鹽水。接種后的葉片置于玻璃平皿中,底部放入潤濕的無菌紗布,每組3張葉片,每組3個平行,置于28℃的人工氣候箱中,12 h光照/12 h黑暗交替培養,注意補水保濕;定期觀察葉片發病情況,對發病葉片進行病原菌的分離和鑒定。
1.3 拮抗菌的篩選及活性測試
1.3.1 菌株抑菌活性初篩
采用平板對峙法對24株供試菌株進行拮抗活性篩選,用無菌打孔器將分離到的病原真菌制成6 mm菌餅,接種于PDA固體平板中央,放置28℃培養,病原真菌C1和C4培養2 d,C2和C3培養1 d后,再將待測放線菌接種于PDA固體平板四周,每板接4株細菌;以僅接種病原真菌菌餅的PDA固體平板為空白對照組,放置28℃培養5~7 d,觀察并記錄病原真菌的生長情況。
1.3.2 菌株發酵產物的抗菌活性測試
選取初篩活性良好的菌株進行菌株發酵產物活性檢測,旨在確定活性產物極性。選擇改良ISP 2培養基作為搖瓶發酵培養基。采用與文獻[19]相同的菌株發酵條件及其次級代謝產物的提取方法。受試菌株留存3類樣品:①發酵液離心后的上清原液(下文簡稱為原液);②上清乙酸乙酯萃取后的甲醇濃縮物(下文簡稱酯相);③菌體丙酮浸提后的甲醇溶解液(下文簡稱菌體浸提物);3種樣品均用一次性過濾器(0.22 μm孔徑)處理后使用。采用牛津杯法[20]檢測菌株發酵產物的抑菌活性,以改良ISP2液體培養基/甲醇溶液為空白對照,每個牛津杯加入初始樣液150 μL,置于28℃培養箱培養7 d,觀察并記錄指示真菌的生長情況,并計算拮抗菌株發酵產物的抑菌率,公式如下:
抑菌圈直徑=對照組病原菌直徑-實驗組病原菌直徑
抑菌率=抑菌圈直徑/對照組病原菌直徑×100%
1.3.3 活性菌株發酵產物的病害防治測試
選取真菌C4侵染健康的沃柑葉片,測試活性菌株發酵產物的病害防治效果。試驗設計2組,按“1.2.4”處理新鮮的待試沃柑葉片。試驗1:用無菌牙簽在葉面上有序扎8~10個孔,將發酵原液/酯相均勻噴灑至葉片的正面和背面,晾干后,在扎孔處滴加10 μL真菌C4的分生孢子液(濃度約為1×107CFU/mL)。試驗2:用無菌牙簽在待試沃柑葉面上有序扎8~10個孔,將10 μL真菌C4的分生孢子液(濃度同上)滴加在扎孔處,培養至葉片出現病斑,再將發酵原液/酯相均勻噴灑接種至葉片的正面和背面。兩組實驗均以噴灑改良的ISP2液體培養基/甲醇溶液作為空白對照,接種后的葉片均置于玻璃平皿中,底部放入潤濕的無菌紗布,每組20張葉片,置于28℃的人工氣候箱中,12 h光照/12 h黑暗交替培養,注意補水保濕,定期觀察葉片發病情況,統計葉片上病斑數量,并計算沃柑葉片的病情指數和菌株發酵產物的防治效果。
參照文獻[18]方法,將病情分級標準制定如下:0級,無病斑;1級,每葉有1~3個病斑;3級,每葉有4~6個病斑;5級,每葉有7~10個病斑。
病情指數=[Σ(各級病葉數×相對級數值)/(試驗總葉數×5)]×100%
防治效果=(對照組病情指數-處理組病情指數)/對照組病情指數×100%
1.4 目標菌株基因組序列antiSMASH在線預測與分析
對目標菌株進行基因組測序分析。將菌株接種于改良ISP2液體培養基中,28℃、180 r/min搖瓶震蕩培養5 d,8000 r/min離心10 min收集菌體,用無菌水洗滌3次后裝入15 mL離心管,迅速冷凍后寄送至上海美吉生物醫藥科技有限公司完成基因組的提取、測序、組裝和注釋。基因組序列通過antiSMASH服務器(https://antismash.secondarymetabolites.org/)進行次級代謝生物合成基因簇的預測。
2 結果與分析
2.1 病原真菌分離與鑒定
2.1.1 菌落形態、生長速率及分生孢子等形態學觀察
從病樣組織中分離出7個菌株,根據其菌落及分生孢子形態特征(圖1)分為4種真菌:①C1菌落呈棕黑色,橙紅色氣生菌絲致密,常產生橙色露珠狀液滴使之呈紅色,生長緩慢(8.02 mm/d);分生孢子呈短桿狀或橢圓狀。②C2菌落呈紫紅色,后期產生紫紅色色素,氣生菌絲白色濃密,生長較快(16.21 mm/d);分生孢子呈鐮刀形。③C3菌落呈粉紅色,氣生菌絲白色濃密,生長較快(16.13 mm/d);分生孢子呈鐮刀形。④C4菌落呈白色或奶白色,氣生菌絲灰白色濃密,生長較慢(12.58 mm/d);分生孢子呈磚格狀或鐮刀形。
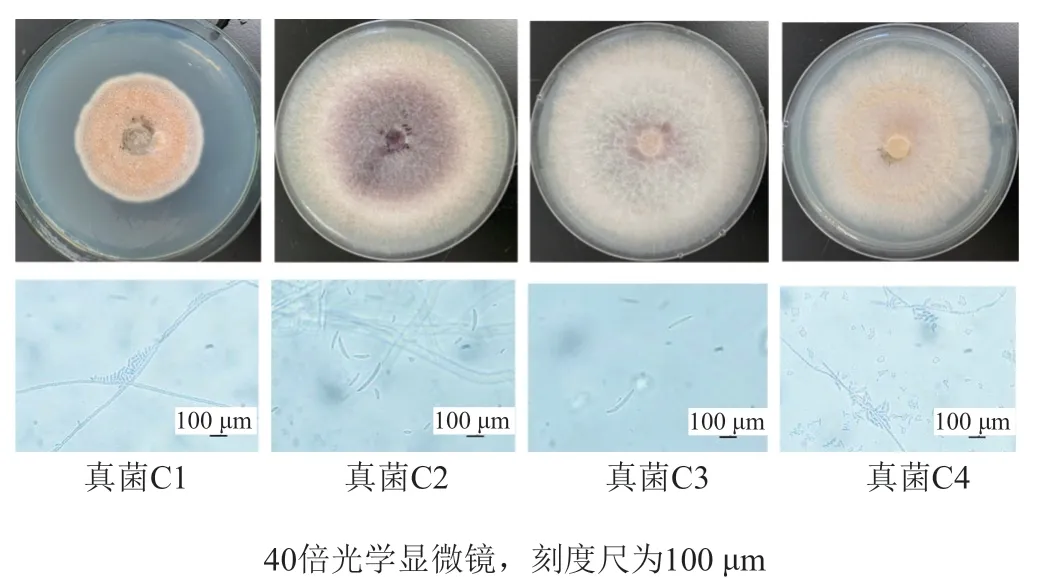
圖1 病樣分離菌株的菌落及分生孢子形態Fig.1 Colony and conidia morphology
2.1.2 病原真菌的分子生物學鑒定
經測序比對,真菌C1的ITS基因序列與Boeremia exigua(登記號MH484014.1)完全一致;真菌C2、C3和C4均屬于鐮刀菌屬(Fusarium)菌株,其ITS基因序列分別與層出鐮刀菌Fusariumproliferatum(登記號HQ380763.1),藤黑鐮孢菌Fusariumfujikuroi(登記號KJ000429.1)和腐皮鐮刀菌Fusarium solani(登記號KT184399.1)一致,并對其構建Neighbour-Joining系統發育樹(圖2);并根據真菌鑒定手冊及相關文獻報道[21-25],這些真菌的分子鑒定結果與觀察到的生物學特性相吻合。
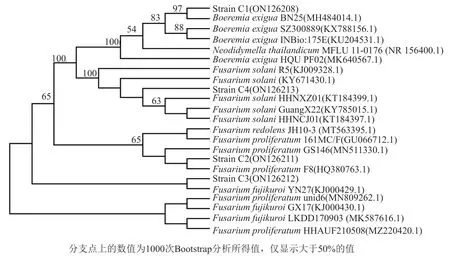
圖2 基于ITS基因序列構建菌株C1,C2,C3和C4的N-J系統發育樹Fig.2 N-J phylogenetic tree of the strain C1,C2,C3 and C4 based on the ITS gene sequences
2.1.3 致病性測定
按照柯赫氏法則,通過針刺接種法將疑似致病菌株C1~C4的分生孢子接種到健康的沃柑葉片上。結果表明,接種C1的葉片在第5天出現褐色病斑,棕色處呈凹陷空洞,病健交界處出現黃綠暈圈(圖3a);接種C4的葉片在第2天出現黑色病斑,外延微微隆起,黑色病斑向外擴張,病癥惡化速度極快(圖3d);且從各自發病的葉片上,重新分離到C1和C4菌株。接種層出鐮刀菌C2(圖3b)和藤黑鐮孢菌C3(圖3c)則均與對照組的表征相同,未見病灶。可見,真菌C1和C4均能感染沃柑葉片,且真菌C4的感染情況較真菌C1的感染情況嚴重。

圖3 C1-C4菌株致病性測定結果(真菌感染5~7d)Fig.3 Orah leaf infected by fungal strain C1-C4 after 5-7 days
2.2 拮抗菌活性測試
2.2.1 拮抗菌的篩選
采用平板對峙法對24株紅樹林放線菌進行拮抗活性初篩,篩選到1株鏈霉菌M2020對4株病原真菌表現出顯著活性(圖4A組)。再用牛津杯法對菌株M2020的發酵原液、發酵液酯相及菌體浸提物進行抑制病原真菌C1~C4的活性測試。結果顯示,M2020菌體浸提物對真菌C1~C4均無抑菌活性,M2020的發酵原液對真菌C1、C2和C3無抑制作用,對真菌C4有抑制活性,抑菌圈小于等于22.74 mm,抑菌率為45.33%。M2020的發酵液酯相對真菌C1、C2、C3和C4均有抑制效果(圖4B組),抑菌圈直徑分別為25.70、47.60、37.10和25.30 mm,抑菌率分別為63.61%、62.63%、53.22%和47.65%。
圖4 鏈霉菌M2020抗真菌C1~C4活性Fig.4 Antifungal activities of strain M2020 against fungal strains C1~C4
2.2.2 鏈霉菌M2020的防病害效果
以致病真菌C4作為指示菌株,通過針刺接種法將其分生孢子接種到健康的柑桔葉片上,用菌株M2020的發酵原液和酯相處理葉片,觀察葉片出現病害的情況,從而判斷M2020菌株的防病害效果。結果表明,菌株M2020的發酵原液和酯相對真菌C4侵染有顯著抑制效果。試驗1中,發酵原液處理組有2片葉子出現褐色病斑,部分葉片的扎孔處均有患病后結痂痕跡(圖5A-1),酯相處理組中有5片明顯病征的葉子,部分病斑出現中心斑塊淡化(圖5A-2);結合葉片患病指數和防治效果(表2),試驗1中發酵原液處理后的葉片患病率較低(患病指數為8.42%),即發酵原液防止真菌C4侵染效果較好(防治效果為89.87%)。試驗2中,發酵原液處理組有5葉片出現黑色病斑,有3處黑斑呈向外擴張惡化嚴重(圖5B-1);酯相處理組中多數葉斑呈現結痂狀態。從表2可知,試驗2中發酵酯相處理的葉片患病率較低(患病指數為5.00%),即發酵液酯相在沃柑葉片出現葉斑后的控制效果佳(防治效果為94.56%)。綜上,在沃柑葉斑病害(真菌C4感染)的防治上,可先用菌株M2020發酵原液處理葉片來預防病原真菌侵染;若葉片出現病癥,可用發酵液酯相控制該病害的擴大。

圖5 M2020發酵產物對沃柑葉片患病前/后處理的藥效試驗圖Fig.5 Field test of strain M2020 fermentation products on pretreatment or post treatment of Ora
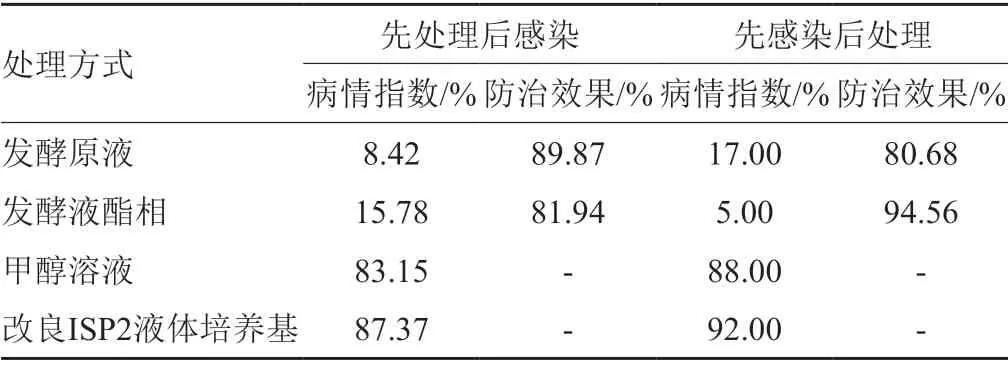
表2 M2020發酵產物對沃柑葉片患病前/后處理的藥效試驗結果Tab.2 Field test of strain M2020 fermentation products on pretreatment or post treatment of Orah
2.3 鏈霉菌M2020次級代謝產物合成潛力分析
基于基因組信息對鏈霉菌M2020合成次級代謝產物的潛力進行預測,菌株M2020基因全長8.1 Mbp,GC含量為72.2 mol%。經antiSMASH預測分析,其潛在的次級代謝產物生物合成基因簇(biosynthetic gene cluster,BGC)共有38個(表3),主要包含聚酮類(T1PKS、T2PKS和T3PKS)、非核糖體多肽類(NRPS)、萜烯類(terpene)、非α-多聚氨基酸類(NAPAA)、核糖體合成和翻譯后修飾肽(RiPP-like) 、羊毛硫肽類(lanthipeptide)、嗜鐵素(siderophore) 和四氫嘧啶(ectoine)等已知基因簇。
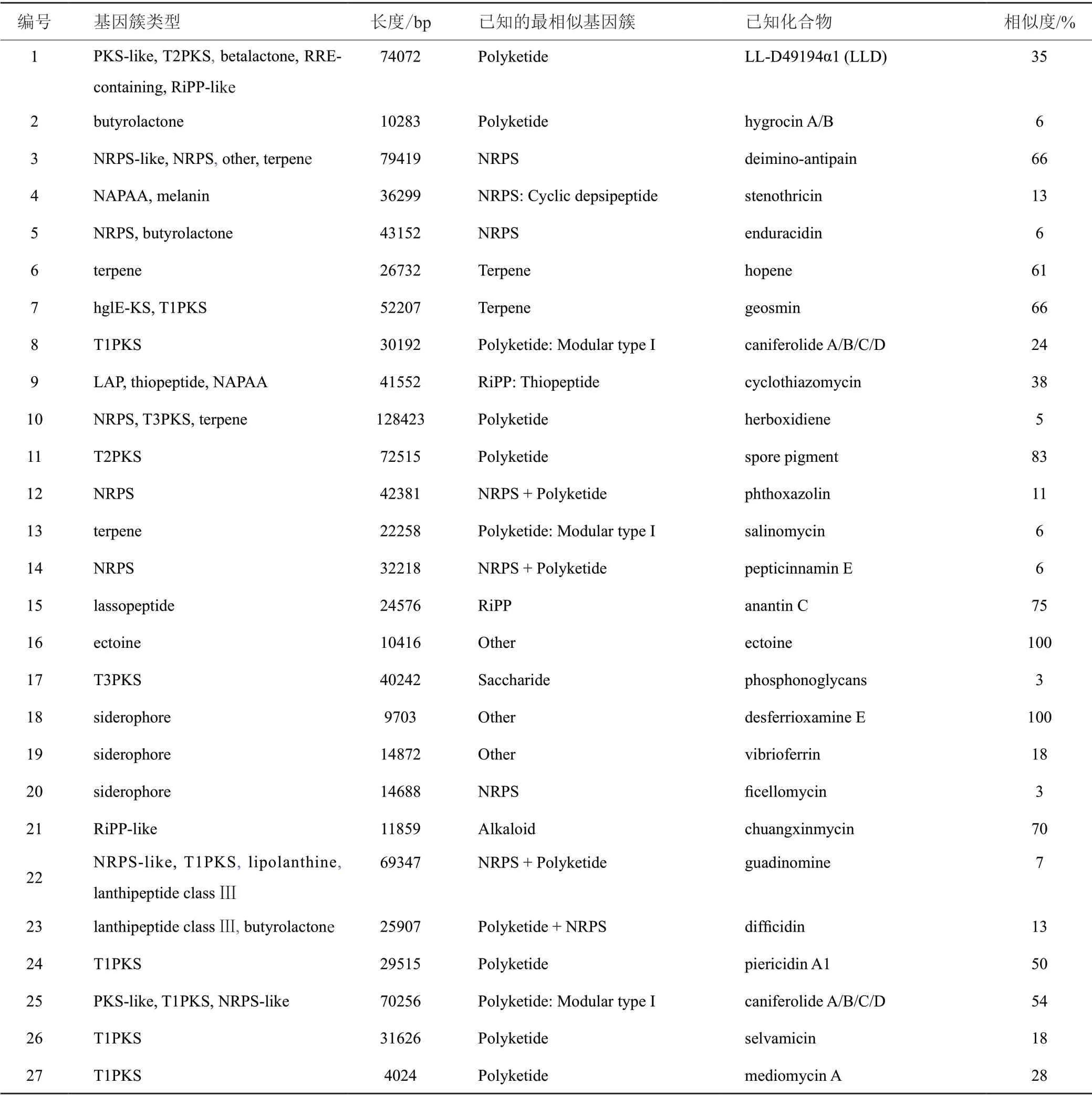
表3 M2020次級代謝產物生物合成基因簇Tab.3 Secondary metabolite biosynthesis gene clusters of M2020
此外,T1PKS(5451、18350和46353 bp)terpene(20950 bp) lanthipeptide class Ⅲ(22729 bp)RiPP-like(10227 bp和10842 bp) lassopeptide(22442 bp)other(32525 bp) butyrolactone (10342 bp和10920 bp)基因簇未匹配到同源基因簇的序列。
3 結論與討論
廣西的沃柑種植面積位居全國第一位,對廣西的農業經濟意義明顯。本研究針對沃柑真菌病害,從廣西南寧采集的沃柑葉片上分離到代號C1~C4的4株真菌,經初步鑒定,分別為大豆莖點霉、層出鐮刀菌、藤黑鐮孢菌和腐皮鐮刀菌。文獻調研表明,大豆莖點霉是破壞性極強的病原真菌,可導致棉花[26]、秋葵[27]、煙草[28]及大豆[29]等農作物患病。鐮刀菌是另一類分布廣泛的植物病原真菌,能侵染多種糧食和飼料作物的根、莖、葉及果實而引起根腐、穗腐和莖腐等病害[30]。其中腐皮鐮刀菌是由厚垣孢子可通過微小創面進入植物體的維管束中萌發,導致一系列植物腐爛病害[31]。本研究的回接實驗證實了大豆莖點霉C1和腐皮鐮刀菌C4能導致沃柑的健康葉片染病。
在此基礎上,利用廣西紅樹林特境來源的24株放線菌對4株病原真菌(代號C1~C4)進行拮抗活性篩選,其中鏈霉菌M2020對真菌C1~C4均表現出拮抗活性,其活性物質分布在發酵原液和酯相中。經16S rRNA基因序列比對分析,菌株M2020與鏈霉菌Streptomyces angustmyceticusNRRLB-2347T系統發育最為密切,相似率為100%。文獻調研表明,Streptomyces angustmyceticus對植物病原真菌具有高效廣譜抑制活性,如對大白菜葉斑病病原菌Colletotrichumsp.和Curvularia lunata的抑菌率達75.6%和69.5%[32],對植物病原真菌尖孢鐮刀菌黃瓜專化型(Fusarium oxysporumf.sp.cucumerinum,Foc)、禾谷鐮刀菌(Fusarium graminearum)和灰葡萄孢(Botrytis cinerea)均有很好的抑制作用[33]。為預測菌株M2020潛在合成產物,對其全基因組進行次級代謝產物生物合成基因簇比對分析。菌株M2020的基因組中預測出38個BGC,其中與已有基因簇相似度較高(30%~100%)的BGC有12個,對應合成具有抑菌活性的已知化合物有4類,即大環內酯caniferolide A/B/C/D、硫肽類抗生素環噻唑霉素cyclothiazomycin、創新霉素chuangxinmycin和聚酮類piericidin A1,菌株M2020可能會產生這些具有抑菌活性的化合物或其類似物。此外,有15個BGC與已有基因簇相似值低于30%,11個BGC未匹配到同源基因簇,這些未知BGC的存在暗示菌株M2020有產生新抗生素的潛力。值得一提的是,S.angustmyceticusNRRLB-2347T生產的一種核苷類抗生素狹霉素(angustmycins),具有很好的植物生長調節活性和一定抑菌活性[34]。總之,鏈霉菌M2020具有顯著且廣譜的抑制真菌活性效果,在沃柑真菌病害防治上得到了一定的驗證。然而,該菌株的抑菌活性是由何種次級代謝產物決定,是否產生狹霉素,其對田間沃柑真菌病害的藥效如何,是否對柑橘其他真菌病害有防治潛力等問題,均是亟須進一步研究的內容。
