肌肉生長抑制素在能量代謝中的調控作用
2022-08-24 05:32:52劉鵬胡亞婕孟令瀅劉洋徐彬
中國畜禽種業 2022年7期
劉鵬 胡亞婕 孟令瀅 劉洋 徐彬
(黑龍江八一農墾大學動物科技學院 163319)
肌肉生長抑制素(myostatin,MSTN)主要表達于肌肉組織,是一種特異性肌肉質量調節因子,參與哺乳動物骨骼肌生長發育調控[1]。當MSTN 遺傳缺失或突變時,小鼠和人的骨骼肌質量顯著增加、肌肉肥大[2]。此外,MSTN 也被報道參與心臟病理、葡萄糖和脂質代謝及某些疾病后期骨骼肌萎縮[3-4]。因此,本文主要就當前MSTN 的相關研究,對其在肌肉生長、能量代謝和相關疾病發生發展中的作用機制做一全面、系統的綜述。
1 MSTN 與肌肉生長
MSTN 屬于轉化生長因子β(transforming growth factor -β,TGF-β)超家族,雖與TGF-β 家族其他成員的同源性很低,但在結構、信號通路和功能上與TGF-β家族有許多相似之處(圖1A)[5]。轉錄翻譯后的無生物活性MSTN 前體蛋白仍需經2 次蛋白酶切才能轉變為具有生物功能的成熟蛋白,這是一個多步水解過程(圖1B)[6]。MSTN 信號主要是通過TGF-β/activin/nodal 途徑發揮其生理學功能,詳細信號轉導途徑如圖1C[7-8]。MSTN 是肌肉生成和出生后早期骨骼肌生長的主要調節劑,其通過激活骨骼肌組織內肌分化因子的轉錄活化來抑制肌細胞生長和發育,從而對肌纖維的形成產生影響[5]。MSTN 基因缺失小鼠比目魚肌中的快肌纖維比例顯著上升,并且在趾長伸肌中,肌纖維從氧化供能型轉向糖酵解型[9]。研究發現,MSTN 基因5′調控區序列多態性影響豬的早期骨骼肌和骨骼發育,且MSTN 表達上升,骨骼肌迅速發育[10]。由于MSTN 對骨骼肌早期生長具有重要調節作用,MSTN 基因缺失或蛋白表達降低會導致肌肉肥大,從而形成“雙肌”現象[11]。但MSTN 過量,同樣對骨骼肌的增殖分化及生長發育有阻礙作用[12]。過量的MSTN 會導致IGF-1/AKT/PI3K 信號受損,從而使FoxO1 激活增強[13]。過量的MSTN 信號以依賴轉錄因子Smad 3 的方式激活FoxO1 和萎縮素,介導泛素-蛋白酶體途徑的關鍵肌肉特異性蛋白降解,造成肌肉萎縮[14]。而運動能減輕MSTN 過量表達所引起的肌肉萎縮現象,如有氧和力量運動顯著減弱MSTN 的表達,并對肌肉萎縮有恢復作用[15]。
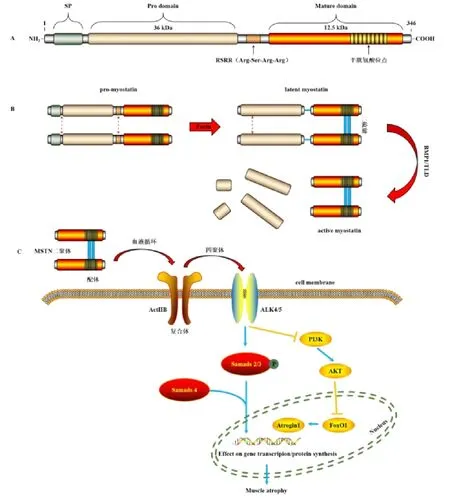
圖1 MSTN 蛋白結構、成熟過程和調控骨骼肌質量信息通路
2 MSTN 與能量代謝
2.1 MSTN 與脂肪沉積
MSTN 是肌肉內脂肪沉積的潛在調節劑,可以影響機體脂質代謝,通過抑制脂肪沉積而減少肥胖[16-17]。瘦素缺陷型肥胖小鼠和高脂肪飲食的野生小鼠肌肉和脂肪組織中MSTN 的mRNA 和蛋白水平增加[18]。此外,MSTN 可抑制前脂肪細胞的分化。MSTN 抑制IGF-1/AKT/PI3K 信號通路,提高FoxO1活性,從而阻礙前脂肪細胞分化[19]。有研究表明,在分化的3T3L1 前脂肪細胞給予重組MSTN 后,脂肪形成受到抑制[20]。同樣,MSTN 可以抑制BMP-7 誘導的脂肪細胞分化,從而抑制脂肪沉積[8]。因此,MSTN 可以通過抑制脂肪細胞分化,引起脂質合成障礙,從而減少脂肪沉積。而將MSTN 基因敲除會導致脂肪沉積率下降。將牛肌肉衛星細胞中MSTN 過表達或敲降,脂肪相關基因的表達均顯示由脂肪合成與脂肪分解的平衡向脂肪分解方向移動[21]。這些研究表明,MSTN 的過度表達和變異都會導致脂肪儲存減少,但其調控機制并不相同。目前關于MSTN 對脂肪合成及分解的作用機制知之甚少,有待進一步研究。
2.2 MSTN 與葡萄糖代謝
MSTN 與葡萄糖代謝之間存在密切關系,現有研究主要集中在MSTN 與胰島素抵抗、葡萄糖運載體蛋白4 等[22-23]。在產生胰島素抵抗的人體內,血液及骨骼肌中的MSTN 蛋白含量明顯上升[22]。在過表達MSTN 蛋白前肽的小鼠中,MSTN 蛋白活性受到抑制,即使高脂飼喂也未產生顯著性的肥胖及胰島素抵抗,表明MSTN 活性與胰島素抵抗存在一定相關性[24]。而過表達MSTN 的CHO 細胞注入裸鼠腿部能引發低血糖現象,且隨著濃度的上升,低血糖現象加重,表明MSTN 介導葡萄糖代謝紊亂[23]。胰島素調節關鍵信號分子AKT 也受到MSTN 調節,MSTN 缺陷小鼠AKT 及其磷酸化顯著高于野生型小鼠[25]。此外,MSTN 敲除能顯著提高葡萄糖運載體4 的表達,而葡萄糖運載體蛋白的不足會誘使葡萄糖代謝紊亂與胰島素抵抗[26]。
2.3 MSTN 與蛋白質代謝
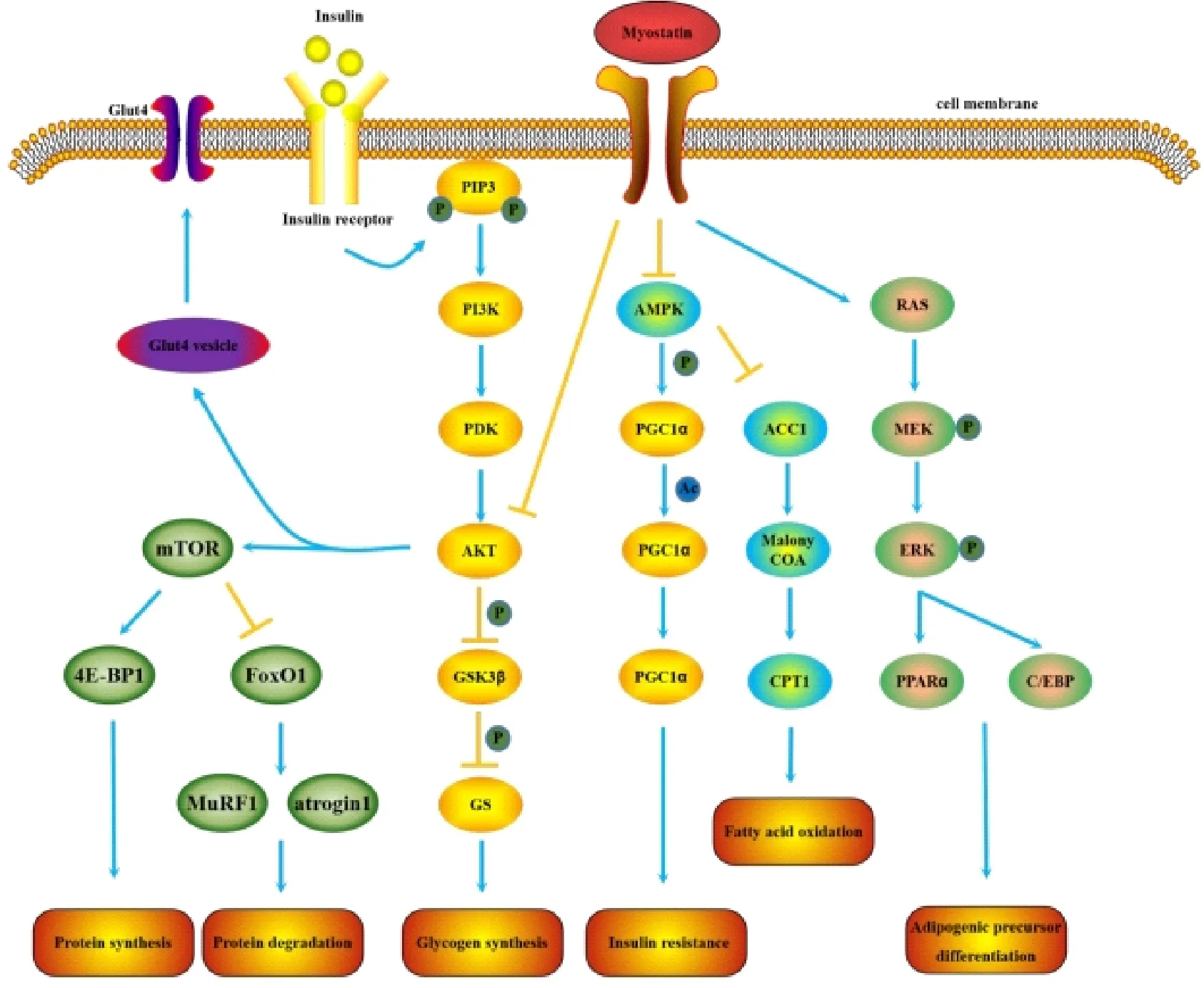
圖2 MSTN 參與葡萄糖代謝、脂肪代謝和蛋白質代謝
MSTN 基因既能參與蛋白質合成代謝,也能參與蛋白質分解代謝。研究發現,在MSTN 敲除小鼠和牛中PI3K/AKT/mTOR 信號通路上調[27]。用MSTN 抑制劑處理新生和成年大鼠,其肌肉中蛋白合成顯著升高。FoxO1 作為調控分解代謝的重要因子之一,可激活泛素-蛋白酶體及自噬溶酶體途徑。而MSTN 蛋白與其受體ActⅡB 結合后,抑制PI3K 活性和AKT磷酸化,從而促使FoxO1 入核。FoxO1 進入細胞核后,再激活MuRFl 與atrogin1,使蛋白質降解[28]。MSTN 敲除小鼠中atrogin1 含量和泛素-蛋白酶體途經的活性顯著低于野生型小鼠[29]。目前,對MSTN 與自噬溶酶體途徑的關注較少。此外,C2C12中MSTN 可促進自噬小體形成,并促進相關基因的表達[30]。MSTN 也可在體外引起鱒魚肌小管萎縮和蛋白降解相關的基因表達上調[31]。
3 MSTN 與疾病
MSTN 還與多種疾病的發生發展相關聯。研究表明,MSTN 在心血管和代謝病理中起重要作用,如心力衰竭、惡病質和胰島素抵抗及2 型糖尿病(type 2 diabetes,T2D)[32-33]。有研究發現,T2D 和非肥胖胰島素抵抗患者肌肉組織中MSTN mRNA 水平較高[34]。給正常小鼠外源注射MSTN 也能提高小鼠肌肉和肝臟的胰島素抵抗[22]。以肥胖和T2D 鼠為模型,抑制MSTN 可在一定程度上降低肥胖和糖尿病表型[35]。因此,抑制MSTN 信號可能是肥胖和糖尿病的一種全新的潛在防治方法。肝硬化、心力衰竭、慢性腎臟疾病、癌癥等疾病通常伴有肌肉萎縮。有研究表明,MSTN 在肝硬化后期表達升高,機體骨骼肌萎縮,而肝硬化又因骨骼肌質量喪失而加重[36]。這表明,疾病期間的肌肉損失常與循環MSTN 水平升高有關。慢性腎臟疾病小鼠肌肉中MSTN 的表達增加了2~3 倍,而注射抗MSTN 抗體后小鼠的體重和肌肉質量下降現象得到緩解[37]。由此可見,抑制MSTN 信號可以緩解消耗性疾病中伴隨的肌肉萎縮。
4 小結
MSTN 作為調節肌細胞的生長與分化關鍵蛋白,在骨骼肌中的功能研究已取得相當大的進展。還有部分研究探討了MSTN 對脂肪組織的影響,結果發現,MSTN 可以降低脂肪沉積率。因此,MSTN 與畜禽個體生長、發育和脂肪沉積有重要關聯,可以用于改善和調節畜禽個體發育情況及瘦肉和脂肪比例。當動物體成肌細胞和前體脂肪細胞形成,繼續成熟分化時,MSTN 對肌肉的抑制和脂質分解作用機制與前期作用機制的差異具有重大的研究意義。此外,MSTN 表達在很多系統性疾病發生發展的過程中起重要作用,通過調節MSTN 信號,可能對預防和治療某些疾病及其并發癥有效。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:10
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
