竹節(jié)參種質(zhì)資源及育種研究進(jìn)展
2022-08-24 08:08:38閔欣怡吳田澤鄭雅芹
世界中醫(yī)藥 2022年13期
閔欣怡 侯 聰 吳田澤 鄭雅芹 鄧 港 劉 霞
(武漢理工大學(xué)化學(xué)化工與生命科學(xué)學(xué)院,武漢,430070)
竹節(jié)參來源于五加科人參屬藥用植物竹節(jié)參PanaxjaponicusC.A.Meyer.,以干燥根莖入藥,又稱竹節(jié)三七、竹節(jié)人參、白三七等[1]。它具有人參和三七的特點(diǎn),具有散瘀、止血、消腫、祛痰、止咳等作用,被稱為“草藥之王”。藥理研究表明,竹節(jié)參對(duì)心血管系統(tǒng)、中樞神經(jīng)系統(tǒng)、免疫系統(tǒng)和消化系統(tǒng)均有保護(hù)作用[2-9]。長(zhǎng)期以來,竹節(jié)參的主要來源是野生資源。由于濫采,其棲息地遭到嚴(yán)重破壞,導(dǎo)致竹節(jié)參野生資源瀕臨滅絕。
1 資源分布
竹節(jié)參分布區(qū)域隨時(shí)間變化較大。《中國(guó)植物志》中記載竹節(jié)參廣泛分布于我國(guó)黃河流域以南濕潤(rùn)半濕潤(rùn)區(qū),分布于中國(guó)東北以及云南、貴州、陜西、湖北、四川、湖南、江西、浙江等省的北緯31°附近的地區(qū),越南、尼泊爾、緬甸、日本和朝鮮也有分布[10]。目前,野生竹節(jié)參資源匱乏,主要分布于西南地區(qū)以及湖北[11]。見表1。
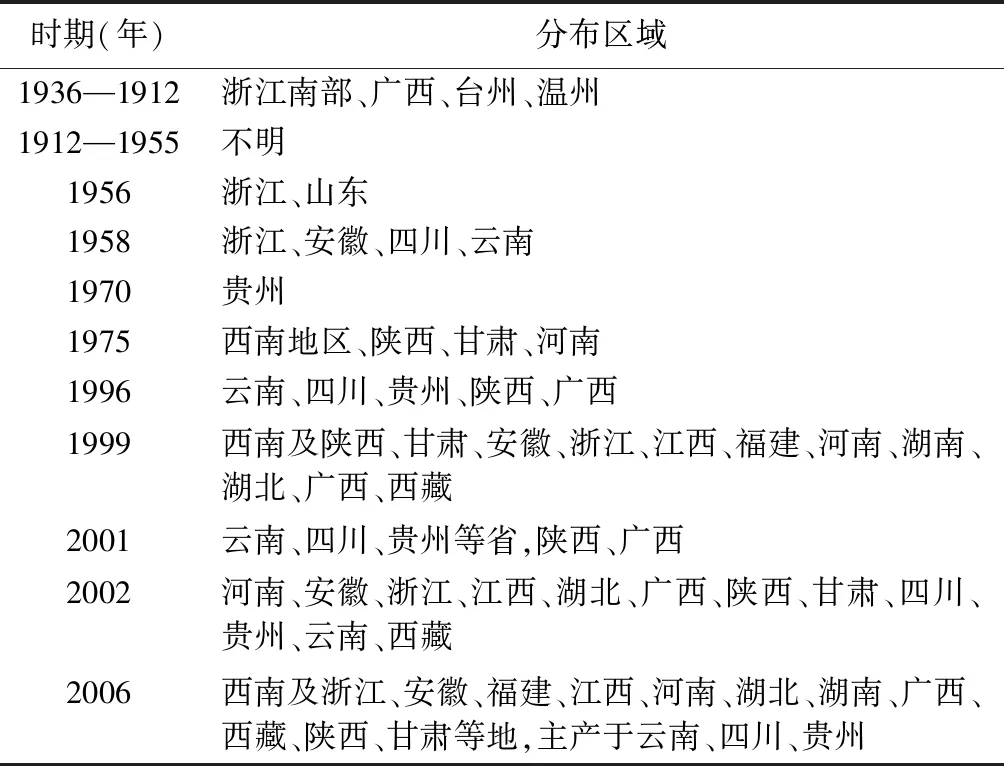
表1 竹節(jié)參分布區(qū)域變化
1.1 野生資源 竹節(jié)參野生資源匱乏,一方面由于濫采濫挖,另一方面在于其生長(zhǎng)的自然環(huán)境受到巨大威脅。在以往調(diào)查過的竹節(jié)參野生資源生長(zhǎng)地區(qū),近年來的調(diào)查顯示,曾經(jīng)野生竹節(jié)參的生長(zhǎng)地區(qū)如今已經(jīng)很難找到其生長(zhǎng)蹤跡[12-16]。
1.2 人工種植 竹節(jié)參目前的人工栽培技術(shù)仍處于剛起步階段,相關(guān)專業(yè)技術(shù)人員較少,規(guī)模化種植基地極少,且缺乏標(biāo)準(zhǔn)化種植技術(shù),導(dǎo)致目前竹節(jié)參種植產(chǎn)量少、效益低[17-21]。
2 種質(zhì)鑒定
2.1 性狀鑒定 竹節(jié)參產(chǎn)品略呈圓柱形,略呈彎曲狀,有些帶有肉質(zhì)側(cè)根。長(zhǎng)度為5~22 cm,直徑為0.8~2.5 cm。表面為黃色或黃褐色、粗糙,有致密的縱向皺紋和根的痕跡。有明顯的節(jié),節(jié)間長(zhǎng)0.8~2 cm,每個(gè)切片有1個(gè)凹陷的莖痕。質(zhì)地堅(jiān)硬,切片呈黃白色至淡黃褐色,有黃色點(diǎn)狀維管束排列成環(huán)狀。無臭、味苦,后略甜[22]。
2.2 顯微鑒定 竹節(jié)參橫截面木栓層有2~10個(gè)細(xì)胞。皮層稍寬,有一些分泌道。維管束外韌型,形成層呈現(xiàn)環(huán)狀。木質(zhì)部多數(shù)排列成2~4組的徑向鏈;木纖維通常為1~4束,有些還會(huì)有木化厚壁細(xì)胞存在。中央有髓質(zhì)。薄壁細(xì)胞含有大量草酸鈣簇晶體,直徑17~70 μm,并含有淀粉粒。粉末是黃白色到黃棕色。木纖維直徑約25 μm。草酸鈣團(tuán)簇更為常見,直徑為15~70 μm。網(wǎng)孔或邊緣口導(dǎo)管直徑為20~70 μm。偶爾可見樹脂束碎片,含有黃色的腫塊物。木栓組織碎片細(xì)胞多呈現(xiàn)矩形或不規(guī)則形,壁厚,淀粉顆粒數(shù)量眾多。
2.3 成分鑒定 Xie等[23]采用超高效液相色譜-四極飛行時(shí)間質(zhì)譜(Ultra-high Performance Liquid Chromatography-Quadrupole Time of Flight Mass Spectrometry,UPLC-QTOFMS)和多元統(tǒng)計(jì)分析技術(shù),對(duì)人參、三七、竹節(jié)參、西洋參和高麗參等5種藥用人參進(jìn)行了代謝產(chǎn)物分析。分析數(shù)據(jù)的主成分分析(Principal Component Analysis,PCA)表明,5種人參藥材可分離為5組不同的植物化學(xué)物質(zhì)。通過PCA的負(fù)荷圖確定了導(dǎo)致這些變化的化學(xué)標(biāo)志物,如人參皂苷Rf、20(S)-假人參皂苷F11、丙二酰-人參皂苷Rb1和人參皂苷Rb2,并通過飛行時(shí)間質(zhì)譜對(duì)各化學(xué)物質(zhì)的含量進(jìn)行了初步鑒定,并基于參考標(biāo)準(zhǔn)進(jìn)行了部分驗(yàn)證。結(jié)果表明,UPLC-QTOFMS對(duì)于快速分析草藥和其他天然產(chǎn)品中存在的一組代謝物是可靠的,并且適用于區(qū)分具有相似化學(xué)成分的人參混品。
竹節(jié)參中主要含有皂苷、糖、多炔、氨基酸、揮發(fā)油和無機(jī)元素等6種化學(xué)成分[24],其中主要活性成分為皂苷。
2.3.1 皂苷 皂苷是人參中研究最多的成分,其含量隨產(chǎn)地有很大的不同。竹節(jié)參以根莖入藥,其中粗皂苷的含量約23.6%,皂苷含量約5%[25]。目前從人參中分離出40多種皂苷,根據(jù)其結(jié)構(gòu)的不同分為齊墩果烷型、達(dá)瑪烷型、奧寇梯木醇型及甾醇型。竹節(jié)參中皂苷類型以齊墩果烷型皂苷為主。見表2。
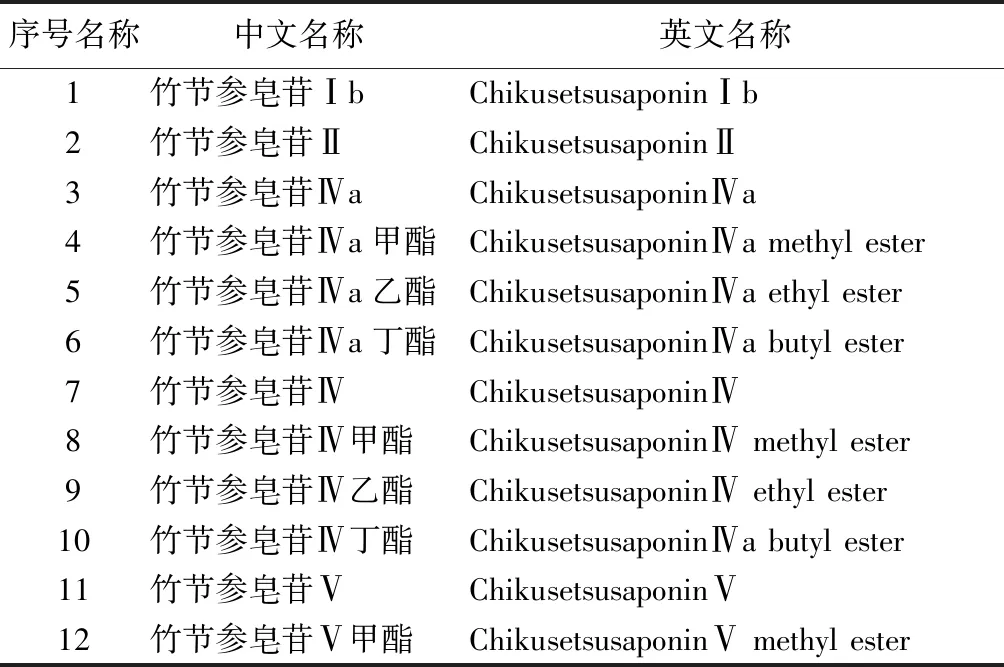
表2 齊墩果酸型化合物
2.3.2 糖 趙海霞等[26]測(cè)定了竹節(jié)參總糖含量、還原糖含量以及總多糖含量。見表3。Ohtani等[27]從日本生長(zhǎng)的竹節(jié)參中和分離出了2種具有激活網(wǎng)狀內(nèi)皮系統(tǒng)作用的多糖:竹節(jié)參多糖(Tochibanan)A和B。陳潔[28]等利用液相色譜-高分辨傅里葉變換離子回旋共振質(zhì)譜法和高效液相色譜(High Performance Liquid Chromatography,HPLC)-紫外檢測(cè)器法定量定性了竹節(jié)參多糖組分中的單糖,是由D-葡萄糖、L-鼠李糖、D-半乳糖醛酸、D-半乳糖和阿拉伯糖組成的。

表3 竹節(jié)參糖類含量
2.3.3 含氮化合物 陳永波等[29]使用HPLC和紅外(Infra-red,IR)光譜,在竹節(jié)參根莖中檢測(cè)出天冬氨酸、酪氨酸、精氨酸、丙氨酸和賴氨酸等17種氨基酸,8種氨基化合物。
2.3.4 多炔類 人參屬植物含有大量的C17多炔化合物[29],也能從竹節(jié)參中分離出來,其中3種多炔化合物首次在人參屬植物中分離出來。
2.4 分子鑒定 由于土壤、氣候、水等生長(zhǎng)環(huán)境對(duì)人參生長(zhǎng)狀況的影響,僅依靠經(jīng)驗(yàn),通過性狀鑒定人參相對(duì)困難。隨著分子生物學(xué)的發(fā)展,中藥鑒定開始進(jìn)入更加科學(xué)和準(zhǔn)確的階段,人參的現(xiàn)代研究變得更加真實(shí)和可靠。Tautz等[30]首次提出使用DNA序列作為生物分類系統(tǒng)的主要平臺(tái)。2003年,他首次提出了線粒體細(xì)胞色素C氧化酶亞基I(Cytochrome C Oxidase Subunit I,COI)的前半部分,作為DNA條形碼的基礎(chǔ),期望編碼所有生物物種,并將其命名為“DNA條形碼”。2004年,生命條形碼聯(lián)盟(Consortium for the Barcode of Life,CBOL)正式成立,旨在開發(fā)一套標(biāo)準(zhǔn)的DNA條形碼制備方法,建立一個(gè)全面的DNA條形碼庫(kù)。最近,生命條碼項(xiàng)目進(jìn)入了一個(gè)新的階段,啟動(dòng)了國(guó)際生命條碼(International Barcode of Life,iBOL)項(xiàng)目。iBOL是一個(gè)來自26個(gè)國(guó)家和地區(qū)的國(guó)際合作項(xiàng)目,旨在建立一個(gè)基于所有真核生物DNA條形碼文庫(kù)的自動(dòng)識(shí)別系統(tǒng)。截至2012年9月生命條形碼數(shù)據(jù)系統(tǒng)(The Barcode of Life Data System,BOLD,http://www.boldsystems.org/)數(shù)據(jù)庫(kù)擁有2 410 651個(gè)標(biāo)本記錄,1 737 588個(gè)標(biāo)本,160 117種生物,包括動(dòng)物、真菌、植物和原生生物,最終目標(biāo)是為全球生物鑒定活動(dòng)提供數(shù)據(jù)支持和服務(wù)。
陳鏡安等[31]利用ITS2序列對(duì)竹節(jié)參進(jìn)行定向測(cè)序,從GenBank下載近緣種和摻假種的ITS2序列。對(duì)ITS2數(shù)據(jù)庫(kù)進(jìn)行裁剪,得到24個(gè)物種的102條序列。應(yīng)用DAMBE程序檢測(cè)ITS2序列的替代飽和。利用MEGA 6.06軟件進(jìn)行序列比對(duì),計(jì)算GC含量,分析變異位點(diǎn),估算種內(nèi)和種間遺傳距離,最終構(gòu)建NJ樹。分析結(jié)果顯示,竹節(jié)參與其不相同的偽種或部分近緣種(秀麗假人參、三葉參、越南人參)之間存在較大差異。但該方法對(duì)竹節(jié)參及其部分近緣種(西洋參、人參、三七)的鑒別有一定的局限性。說明ITS2序列可作為竹節(jié)參及其不相同偽品鑒定的DNA條形碼之一,為竹節(jié)參及其近緣種間遺傳親緣關(guān)系的鑒定、竹節(jié)參與不相同摻假品種的鑒別以及竹節(jié)參的臨床安全性提供了依據(jù)。
3 遺傳多樣性
遺傳多樣性是生物多樣性的一個(gè)重要組成部分,它是生物體進(jìn)化和物種分化的來源。種內(nèi)遺傳多樣性是學(xué)者們經(jīng)常研究的遺傳多樣性。地球上所有生物體的遺傳信息的總和是廣泛的遺傳多樣性。
Choi等[32]通過140對(duì)引物的實(shí)驗(yàn)開發(fā)了70個(gè)表達(dá)序列Tag衍生的多態(tài)性簡(jiǎn)單序列重復(fù)標(biāo)記。這70個(gè)標(biāo)記在4個(gè)人參品種中均表現(xiàn)出可重復(fù)的多態(tài)性,其中19個(gè)標(biāo)記在6個(gè)人參品種。這些標(biāo)記在2個(gè)P之間雜交的F2群體中以1∶2∶1的孟德爾遺傳方式分離。人參品種“Yunpoong”和“Chunpoong”,表明這些是可復(fù)制和遺傳的可映射標(biāo)記。利用基因型數(shù)據(jù)進(jìn)行的系統(tǒng)發(fā)育分析顯示有3個(gè)不同的類群:人參-竹節(jié)參分化枝、西洋參和三七,相似系數(shù)為0.70。竹節(jié)參與人參2個(gè)物種具有相似的遺傳背景。人參品種分為3個(gè)小類群:一個(gè)獨(dú)立品種“Chunpoong”,一個(gè)亞類群有3份材料,包括2個(gè)品種“Gumpoong”和“Yunpoong”以及一個(gè)長(zhǎng)白種“Hwangsook”,另一個(gè)亞類群有2份材料,包括一個(gè)品種“Gopoong”和一個(gè)長(zhǎng)白種“Jakyung”。每對(duì)引物產(chǎn)生1到4條帶,表明人參基因組具有高度復(fù)制的古多倍體基因組結(jié)構(gòu)。
程揚(yáng)等[33]開發(fā)和鑒定了竹節(jié)參SSR分子標(biāo)記,深入研究了竹節(jié)參遺傳多樣性。基于竹節(jié)參轉(zhuǎn)錄組數(shù)據(jù),利用MISA軟件對(duì)SSR序列進(jìn)行搜索和分析。根據(jù)SSR兩翼序列設(shè)計(jì)SSR引物,并對(duì)反應(yīng)體系進(jìn)行優(yōu)化。以5個(gè)不同種群為模板,對(duì)SSR引物進(jìn)行多態(tài)性檢測(cè)。共發(fā)現(xiàn)12 244個(gè)SSR位點(diǎn),分布在66 403個(gè)unigenes基因10 299個(gè)單基因中,SSR頻率為15.51%。SSR重復(fù)序列最多的分別為AT/AT(44.87%)、AAG/CTT(24.00%)、AAAAT/ATTTT(19.28%)和AAAAAT/ATTTTT(6.54%)。最后,通過對(duì)竹節(jié)參樣品的引物多態(tài)性檢測(cè),獲得18對(duì)穩(wěn)定的SSR引物。
竹節(jié)參SSR位點(diǎn)豐富而密集,為竹節(jié)參及其近緣種的分子鑒定和遺傳多樣性研究奠定了分子基礎(chǔ)。
4 育種研究
4.1 繁殖方式 竹節(jié)參有無性繁殖和有性繁殖2種繁殖方式[34]。竹節(jié)參一般采用種子播種繁殖方法,可以在8月中旬,選擇在年末生長(zhǎng)強(qiáng)勁,無病蟲害,大顆粒,成熟早的4年以上的植物果實(shí)收集種子,去掉果皮,用0.3%高錳酸鉀溶液或3.5%~4%的甲醛水溶液浸泡10 min,沖洗后取出,再用濕沙保存,種子∶河沙為1∶4。在保存期間,應(yīng)注意防止?jié)裆掣稍铩R话銇碚f,濕沙子應(yīng)該揉成一個(gè)球。保存好的種子也需要在播種前挑選好,除去小或者保存過程中有霉變或失水的種子,然后用上述方法進(jìn)行1次消毒處理,即可播種。周益權(quán)等[35]對(duì)竹節(jié)參無性繁殖技術(shù)進(jìn)行了研究,以不同位置育苗的人參為研究對(duì)象,研究了不同基質(zhì)和頂芽保留對(duì)人參出苗和多芽形成的影響。結(jié)果表明,隨著齡期的增加,出苗數(shù)量和出苗率降低;有機(jī)質(zhì)含量高的基質(zhì)可促進(jìn)多芽的形成。頂芽的保留對(duì)竹節(jié)參復(fù)芽的形成有很大的影響。去頂芽處理顯著提高了竹節(jié)參的復(fù)芽率,最高為30.00%。通過處理頂芽和增加基質(zhì)養(yǎng)分可以提高竹節(jié)參的多梢發(fā)生率。陳士林等[36]在研究中公開了一種利用芽莖組織培養(yǎng)人參的方法,提高了竹節(jié)參人工栽培的成活率,增加竹節(jié)參產(chǎn)量的問題。
4.2 種植方法 林下栽培是竹節(jié)參種植生產(chǎn)的最重要的途徑之一。根據(jù)徐燃等[37]的研究,竹節(jié)參生產(chǎn)基地應(yīng)遠(yuǎn)離城市、公路、工業(yè)區(qū)及周邊污染源,生產(chǎn)環(huán)境、空氣質(zhì)量、種植土壤、灌溉水質(zhì)、地下水水質(zhì)均符合現(xiàn)行國(guó)家標(biāo)準(zhǔn),水資源水質(zhì)、空氣水質(zhì)、土壤水質(zhì)定期檢測(cè)評(píng)價(jià)。根據(jù)周益權(quán)等[38]的研究,選擇闊葉林或針寬混交天然次生林,遠(yuǎn)離重工業(yè)污染和沒有化學(xué)除草劑、稀疏而高大的樹木,在森林樹冠下開墾;高封閉的針葉林或大量的竹鞭表土不適宜。選擇陰坡或半陰坡,坡度為5%~25%的山坡中段種植。選擇松散、肥沃、濕潤(rùn)、排水良好,表面腐殖質(zhì)大于5 cm。氣候寒冷潮濕,年平均氣溫約10 ℃,極端最高氣溫小于35 ℃,極端最低氣溫大于-12 ℃;10 ℃以上的年活動(dòng)累積溫度大于2 700 ℃,年平均降水量大于1 500 mm。每年9月初至10月底種植前,砍伐0.6~0.7 m距離茂密的樹木和小灌木,清除1.5 m以下的雜草和灌木,清除林地清潔材料;準(zhǔn)備深度25~35 cm,清除土壤屑,施用殺蟲劑,使用有機(jī)肥,每隔667 m使用全腐豬、牛、羊糞施用2 000 kg,按自然地形逐耕混土,箱寬1~1.2 m,箱長(zhǎng)1.2 m,開溝起壟深15~25 cm,寬25~35 m。竹節(jié)參苗體形態(tài)自然、根較長(zhǎng)、無病、無機(jī)械損傷,單重量大于10 g,有發(fā)芽;第1種參苗為分苗,為根頂端,頂芽清晰生長(zhǎng)良好(該苗保證第2年出苗),然后剪長(zhǎng)2~4 cm。如果沒有頂芽,可以根據(jù)根芽的痕跡將每段切成1個(gè)芽,在苗圃中栽培1年,然后進(jìn)行下林移栽。第2種竹節(jié)參幼苗種植種子苗,種子幼苗大于2年生,單株重量在5 g以上,種植前用多菌靈溶液1∶800,或甲基溶液1∶800浸泡約45 min,或在塊莖苗切口位置浸入植物灰分種植到林地。人工栽培竹節(jié)參可以保持豐富的營(yíng)養(yǎng),一般來說,野生竹節(jié)參的產(chǎn)量以及品質(zhì)遠(yuǎn)低于栽培竹節(jié)參。
4.3 組織培養(yǎng) 竹節(jié)參的市場(chǎng)需求量日益增加,匱乏的野生資源無法滿足市場(chǎng)需求,對(duì)人參資源育種,人工栽培是解決這一問題的重要途徑,組織培養(yǎng)有利于實(shí)現(xiàn)規(guī)模、標(biāo)準(zhǔn)化種植,有利于種質(zhì)資源的有效利用,可以解決育種時(shí)間、幼苗短缺等問題。羅正偉等[39]以人參胚、莖和葉為外植體,研究分離培養(yǎng)和植物再生技術(shù),優(yōu)化培養(yǎng)條件,應(yīng)用正交法篩選竹節(jié)參最佳愈傷組織誘導(dǎo)分化培養(yǎng)基。
4.4 品質(zhì)提升 竹節(jié)參的次生代謝產(chǎn)物作為主要有效成分,利用基因工程技術(shù)對(duì)其代謝途徑進(jìn)行定向調(diào)控,提升相關(guān)有效成分的產(chǎn)量,從而提高竹節(jié)參的產(chǎn)品品質(zhì)。Rai等[40]描述了基于Illumina的RNA測(cè)序分析,以表征竹節(jié)參及其與其他人參屬植物的比較RNA測(cè)序和從頭轉(zhuǎn)錄組組裝。竹節(jié)參共產(chǎn)生135 235個(gè)單基因,其中78 794個(gè)(58.24%)單基因被NCBI nr數(shù)據(jù)庫(kù)注釋。5種組織的轉(zhuǎn)錄組分析和基因本體富集分析結(jié)果表明,盡管整個(gè)過程在所有組織中都是均勻保守的。然而,每個(gè)組織都有幾個(gè)獨(dú)特的單基因,在所研究的組織中,葉子顯示出最獨(dú)特的單基因。對(duì)竹節(jié)參轉(zhuǎn)錄組與其他人參屬物種的公開轉(zhuǎn)錄物組裝,即人參、竹節(jié)參和西洋參在所有人參屬物種中也表現(xiàn)出高度的序列相似性。竹節(jié)參轉(zhuǎn)錄組鑒定出編碼三萜主鏈生物合成途徑中所有酶的推定基因,并分別鑒定出24個(gè)和48個(gè)單基因,分別被注釋為細(xì)胞色素P450(Cytochrome P450,CYP)和糖基轉(zhuǎn)移酶(Glycosyltransferase,GT)。這些CYP和GT注釋的單基因在所有人參屬物種中都是保守的,并與參與三萜類主干生物合成途徑的其他轉(zhuǎn)錄物共同表達(dá)。確定的單基因是參與三萜皂苷生物合成的有力候選基因,可以作為未來驗(yàn)證研究育種的基礎(chǔ)。
Zhang等[41]對(duì)野生珠子參(竹節(jié)參的一種變種)的根莖節(jié)和節(jié)間進(jìn)行了比較代謝產(chǎn)物和轉(zhuǎn)錄組分析,以揭示它們?cè)谌圃碥丈锖铣芍械牟煌饔谩=Y(jié)果表明,節(jié)點(diǎn)是根莖中人參皂苷合成的關(guān)鍵部位。節(jié)點(diǎn)中齊墩果酸型人參皂苷的含量遠(yuǎn)高于節(jié)間。大多數(shù)類異戊二烯生物合成相關(guān)基因在淋巴結(jié)中高度表達(dá)。此外,還發(fā)現(xiàn)候選尿苷二磷酸糖基轉(zhuǎn)移酶(Uridine Diphosphate Glucuronosyltransferase,UGT)基因在節(jié)點(diǎn)和節(jié)間之間存在差異表達(dá)。這些均有助于更好地了解竹節(jié)參及其變種根莖中人參皂苷的代謝以及后續(xù)對(duì)相關(guān)基因進(jìn)行調(diào)控以幫助品質(zhì)的提升。
Tang等[42]通過比較竹節(jié)參莖尖的內(nèi)部解剖結(jié)構(gòu)、激素含量、淀粉含量和分子轉(zhuǎn)錄水平的差異,探討了根狀莖膨大的機(jī)制。Illumina測(cè)序產(chǎn)生了1 333個(gè)差異表達(dá)基因(Differentially Expressed Genes,DEGs),這些基因在2個(gè)樣本之間差異表達(dá)。通路富集分析表明,參與淀粉和蔗糖代謝、二萜生物合成和植物激素信號(hào)轉(zhuǎn)導(dǎo)通路的DEGs顯著富集。此外,淀粉測(cè)定結(jié)果表明,膨脹部分的淀粉含量顯著高于細(xì)長(zhǎng)部分的淀粉含量。此外,激素測(cè)定結(jié)果表明,赤霉素A1(GA1)、赤霉素A53(GA53)、吲哚-3-乙酸(IAA)和茉莉酸(JA)含量負(fù)相關(guān),而順式玉米素(cZ)含量與根莖膨大正相關(guān)。結(jié)果篩選出了幾個(gè)DEGs和潛在的根狀莖膨大候選基因,為人參屬植物膨大基因的鑒定以及后續(xù)提高產(chǎn)量等奠定了基礎(chǔ)。
Xu等[43]利用代謝產(chǎn)物分析,分析顯示齊墩果酸型和達(dá)馬拉型皂苷在竹節(jié)參的3種組織(幼根組織、次根組織、老根組織)中分布不均。單分子實(shí)時(shí)(Single Molecule Real-time,SMRT)測(cè)序和下一代測(cè)序(Next Generation Sequencing,NGS)數(shù)據(jù)揭示了與這2種皂苷的產(chǎn)生相關(guān)的不同的組織特異性轉(zhuǎn)錄組模式。在共表達(dá)網(wǎng)絡(luò)和層次聚類分析中,發(fā)現(xiàn)1個(gè)3-羥基-3-甲基戊二酰輔酶A還原酶(3-hydroxy-3-methylglutaryl-coenzyme A Reductase,HMGR)和2個(gè)1-脫氧-D-木酮糖-5-磷酸合成酶(1-deoxy-D-xylulose-5-phosphate Synthase,DXS)等轉(zhuǎn)錄物分別是與竹節(jié)參齊墩果酸和達(dá)馬烷型皂苷生物合成相關(guān)的關(guān)鍵基因。此外,還發(fā)現(xiàn)CYP和UGT家族蛋白作為皂苷生物合成相關(guān)基因的調(diào)節(jié)因子,表現(xiàn)出組織特異性表達(dá)模式。這些結(jié)果共同提供了竹節(jié)參的全面代謝組學(xué)和轉(zhuǎn)錄組學(xué)概述,為竹節(jié)參定向育種奠定基礎(chǔ)。
5 結(jié)論
在現(xiàn)代中藥研究領(lǐng)域中,人參皂苷等提取物的藥理作用豐富,對(duì)新藥研發(fā)具有重要的指導(dǎo)價(jià)值。它值得進(jìn)一步研究,在衛(wèi)生保健產(chǎn)品、食品等領(lǐng)域的價(jià)值也在逐步發(fā)展。目前,人參分布廣泛,具有較高的人工栽培價(jià)值,遺傳多樣性豐富,其潛在應(yīng)用價(jià)值值得進(jìn)一步開發(fā)。近年來,由于竹節(jié)參需求的增加,野生竹節(jié)參資源稀缺。除了保護(hù)竹節(jié)參資源和尋找新資源外,還可以從其他植物中尋找替代藥用資源,填補(bǔ)高活性成分的藥用資源的空白,解決市場(chǎng)供需矛盾。
