響應面法優化雞肝發酵工藝
2022-08-30 09:00:08馬晶晶吳瑀婕王道營徐為民
肉類研究 2022年8期
馬晶晶,吳瑀婕,李 良,黃 瑾,楊 靜,3,楊 彪,3,鄒 燁,*,王道營,*,徐為民
(1.江蘇省農業科學院農產品加工研究所,江蘇 南京 210014;2.江蘇大學食品與生物工程學院,江蘇 鎮江 212013;3.農業農村部農產品冷鏈物流技術重點實驗室,江蘇 南京 210095;4.南京農業大學食品科技學院,江蘇 南京 210095)
作為全球第二大肉雞生產和消費大國,2021年我國肉雞總產量為1 998.1 萬t。雞肝是雞屠宰加工過程中的主要副產品之一,因此每年會產生大量雞肝。雞肝中蛋白質的含量約占總成分的20%,與禽肉中的蛋白質含量接近,同時還具有平衡的氨基酸比例結構,富含VA、VB、VB、VE、鐵、硒等維生素及微量元素,營養價值高,對人體健康有益。然而,由于雞肝具有較重的腥苦味,在普通醬鹵過程中難以去除,較難被消費者接受,且雞肝的膽固醇含量較高,多食對健康無益,大多數雞肝以鮮銷或僅經簡單加工以動物飼料的形式被利用,精深加工利用率并不高,造成大量優質資源的浪費。
乳酸菌是一類能利用可發酵碳水化合物產生大量乳酸的細菌,屬于厭氧或兼性厭氧菌。其中植物乳桿菌()是一種廣泛分布于自然界、動物腸道及眾多發酵食品中的乳酸菌。鑒于其多種益生活性,植物乳桿菌在食品工業、飼料生產和臨床醫學中發揮著不可替代的作用。研究表明,植物乳桿菌能在短時間內產生大量酸類物質,使體系pH值下降至較低水平,從而抑制部分腐敗菌和雜菌的生長,這些酸類同時也是風味成分的前體物質,能與醇類反應生成酯類物質,有利于促進產品風味的形成。
雞肝蛋白質含量較高,可為乳酸菌的生長繁殖提供營養基質。雞肝中的蛋白質在微生物和酶的共同作用下降解為小肽和游離氨基酸等低分子質量產物,營養價值提高。隨著全球人口快速增長,對食物資源尤其是蛋白質資源的需求正不斷增加,開展雞肝副產物綜合利用的研究具有十分重要的意義,有利于提升對優質食物資源的綜合利用,為畜禽屠宰加工業的持續發展提供強有力的保障。
本研究以雞肝為原料,從植物乳桿菌LP1、發酵乳桿菌()LF1、枯草芽孢桿菌()BS1中選擇植物乳桿菌對雞肝進行發酵,通過優化得到植物乳桿菌發酵雞肝的最佳工藝,并測定雞肝發酵前后游離氨基酸的變化,為利用發酵雞肝制備寵物食品提供理論依據,拓展雞肝副產物的綜合利用思路。
1 材料與方法
1.1 材料與試劑
新鮮六和雞肝,購于南京孝陵衛菜市場。放置于冰柜中冷凍保存。植物乳桿菌LP1、發酵乳桿菌LF1、枯草芽孢桿菌BS1均由江蘇省農業科學院農產品加工研究所畜禽加工實驗室保藏。
MRS液體培養基 北京索萊寶科技有限公司;三氯乙酸(分析純) 上海麥克林生化科技有限公司;鹽酸(分析純) 國藥集團化學試劑有限公司;葡萄糖、氫氧化鈉(分析純) 四川西隴科學股份有限公司。
1.2 儀器與設備
PTX-FA210S電子天平 福州華志科學儀器有限公司;Five Easy Plus pH計 梅特勒-托利多(上海)有限公司;Zealway GI54DWS滅菌器 致微儀器有限公司;超凈操作臺 蘇州凈化設備有限公司;SPX-250B-Z生化培養箱 上海博迅實業有限公司醫療設備廠;ZQZY-75BS振蕩培養箱 上海知楚儀器有限公司;T-25數顯勻漿機德國IKA公司;Gen5全波長酶標儀 美國伯騰儀器有限公司;TG16-WS臺式高速離心機 湖南湘儀實驗室儀器開發有限公司;L8900氨基酸分析儀 日本日立公司。
1.3 方法
1.3.1 原料預處理
將冷凍的新鮮雞肝置于室溫解凍,解凍后用流水清洗,剔除肝葉表面的苦膽和經絡,然后放入料理機中勻漿(勻漿15 s,間隔10 s后再次勻漿15 s,重復勻漿3 次),將勻漿后的雞肝用無菌均質袋分裝,放置在-20 ℃冰箱冷凍備用。
1.3.2 雞肝發酵準備
稱取勻漿后的雞肝20 g至250 mL錐形瓶中,加入適量蒸餾水,調節漿液pH值,按照錐形瓶中的雞肝質量加入適量葡萄糖,混合均勻,于121 ℃滅菌15 min,得到無菌雞肝發酵培養基,備用。
1.3.3 發酵劑菌種的選擇
從實驗室提供的植物乳桿菌LP1、發酵乳桿菌LF1和枯草芽孢桿菌BS1中選擇一種作為發酵劑。
1.3.3.1 菌種活化及培養
參照王夢曼等的方法,稍作修改。將植物乳桿菌LP1、發酵乳桿菌LF1以2%(/)的接種量接種于MRS液體培養基,于37 ℃恒溫培養箱中靜置培養24 h;枯草芽孢桿菌BS1以2%的接種量接種于LB液體培養基,37 ℃、200 r/min搖床培養24 h。確定發酵菌株后,轉接2 代恢復活力,培養至活菌數為10CFU/mL,于4 ℃、8 000 r/min條件下離心10 min,棄去上層液體培養基,沉淀的菌泥用同體積的無菌生理鹽水洗滌3 次,將菌懸液于4 ℃保存,備用。
1.3.3.2 生長曲線及產酸曲線的測定
參照張玉的方法,并稍作修改。活化后的植物乳桿菌LP1和發酵乳桿菌LF1菌株以2%的接種量接種于50 mL MRS液體培養基,在37 ℃恒溫培養箱中靜置培養24 h,枯草芽孢桿菌BS1以2%的接種量接種于50 mL LB液體培養基,在37 ℃、200 r/min搖床培養24 h,間隔2 h取適量培養物測定600 nm波長處光密度(OD)及pH值,同時以空白MRS液體培養基和LB液體培養基作對照,平行測定3 次取平均值,繪制生長曲線及產酸曲線,根據菌種的生長與產酸情況選擇合適菌株作為發酵劑。
1.3.4 單因素試驗設計
選擇植物乳桿菌LP1為發酵菌種,將重懸后的菌懸液以一定的比例接種到雞肝發酵培養基中。利用三氯乙酸(trichloroacetic acid,TCA)能夠溶解小分子蛋白質或肽段的特性,將發酵后雞肝與15 g/100 mL TCA混合,離心取上清液測定280 nm波長處吸光度(),用于表征小分子蛋白質或肽段的含量,越大說明小分子肽和游離氨基酸的含量越高,發酵效果越好。
1.3.4.1 發酵初始pH值對的影響
按照1.3.2節,稱取20 g雞肝至250 mL錐形瓶中,以料液比1∶3.5(/)加入純水,調節pH值分別為4.0、4.5、5.0、5.5、6.0、6.5,滅菌冷卻后,以葡萄糖添加量6%、接種量1.5%的條件在37 ℃恒溫培養箱中發酵24 h,將雞肝發酵物與15 g/100 mL TCA混合均勻,靜置30 min,然后4 ℃、8 000 r/min離心10 min,取上清液稀釋至合適濃度(在2.5以內)測定。
1.3.4.2 葡萄糖添加量對的影響
以料液比1∶3.5向雞肝中加入純水,分別加入2%、4%、6%、8%、10%葡萄糖,調節pH值為5.5,滅菌冷卻后,接種1.5%的發酵菌種,其余步驟同1.3.4.1節。
1.3.4.3 接種量對的影響
以料液比1∶3.5向雞肝中加入純水,加入6%葡萄糖,調節pH值為5.5,滅菌冷卻后,分別接種0.5%、1.0%、1.5%、2.0%、2.5%、3.0%的發酵菌種,其余步驟同1.3.4.1節。
1.3.4.4 發酵時間對的影響
以料液比1∶3.5向雞肝中加入純水,加入6%葡萄糖,調節pH值為5.5,滅菌冷卻后,接種1.5%的發酵菌種,在37 ℃恒溫培養箱中發酵16、20、24、28、32 h,其余步驟同1.3.4.1節。
1.3.4.5 料液比對的影響
分別以料液比1∶2.0、1∶2.5、1∶3.0、1∶3.5、1∶4.0、1∶4.5向雞肝中加入純水,加入6%葡萄糖,調節pH值為5.5,滅菌冷卻后,接種1.5%的發酵菌種,其余步驟同1.3.4.1節。
1.3.5 響應面試驗設計
綜合1.3.4節單因素試驗結果確定各單因素取值范圍,選擇接種量()、發酵初始pH值()、料液比()為自變量,以為響應值,進行3因素3水平響應面試驗設計,采用Box-Behnken設計試驗方案,具體見表1。
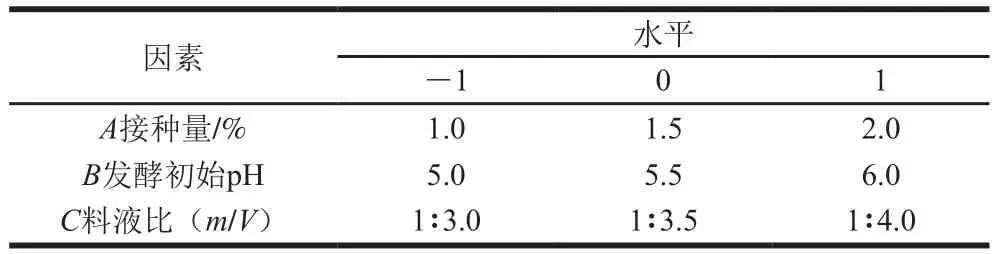
表1 響應面試驗因素水平設計Table 1 Codes and levels of independent variables in Box-Behnken design
1.3.6 發酵雞肝測定
取1 g發酵雞肝固液混合物,加入5 mL 15 g/100 mL TCA后混合均勻,靜置30 min,4 ℃、10 000 r/min離心5 min,將離心后得到的上清液適當稀釋,測定。
1.3.7 游離氨基酸測定
參照Yang Xue等的方法,稍作修改。稱取0.05 g凍干的發酵前后雞肝樣品,與5 mL 10 g/100 mL TCA溶液混合均勻,于4 ℃靜置2 h,并在4 ℃、10 000 r/min的條件下離心10 min,取2 mL離心后的上清液用6 mol/L NaOH溶液調節至pH值為2,并用pH 2、0.06 mol/L TCA溶液定容至10 mL,經0.22 μm的針筒式有機相濾膜過濾后,將濾液轉移至2 mL的上樣瓶中,采用氨基酸自動分析儀上機檢測。
1.4 數據處理
采用Excel 2019進行數據統計處理,用Origin 2018軟件進行作圖分析,用SPSS(IBM SPSS Statistics 19)軟件進行方差分析,<0.05為顯著性檢驗標準。采用Design-Expert 8.0.6軟件進行響應面優化試驗設計及數據分析處理。
2 結果與分析
2.1 發酵劑菌種的選擇
2.1.1 不同菌種的生長曲線
由圖1可知,植物乳桿菌LP1和發酵乳桿菌LF1顯示出類似的生長趨勢,2 種菌株的生長周期均較短,而枯草芽孢桿菌BS1的生長速率比植物乳桿菌LP1和發酵乳桿菌LF1小。0~4 h為植物乳桿菌LP1和發酵乳桿菌LF1的生長延滯期,此時菌種剛剛接種到新鮮培養基中,為了適應新環境,菌種需要對自身的代謝系統進行調整,菌種細胞數量及生物量增長率在延滯期處于最低水平;4~8 h為對數生長期,此時菌種已適應生長環境,生長速率達到最快;經過一段時間的生長后,隨著培養基中營養物質的消耗、代謝產物的積累,此時新增細胞數和死亡細胞數維持相對平衡,在生長曲線中表現為接近水平的線。枯草芽孢桿菌BS1的生長延滯期、對數期、穩定期分別為0~8 h、8~12 h及12 h后。植物乳桿菌LP1在對數期的生長速率高于發酵乳桿菌LF1和枯草芽孢桿菌BS1,且達到生長穩定期后,植物乳桿菌LP1的OD整體高于發酵乳桿菌LF1和枯草芽孢桿菌BS1。這說明植物乳桿菌LP1和發酵乳桿菌LF1較枯草芽孢桿菌BS1具有更強的環境適應性。
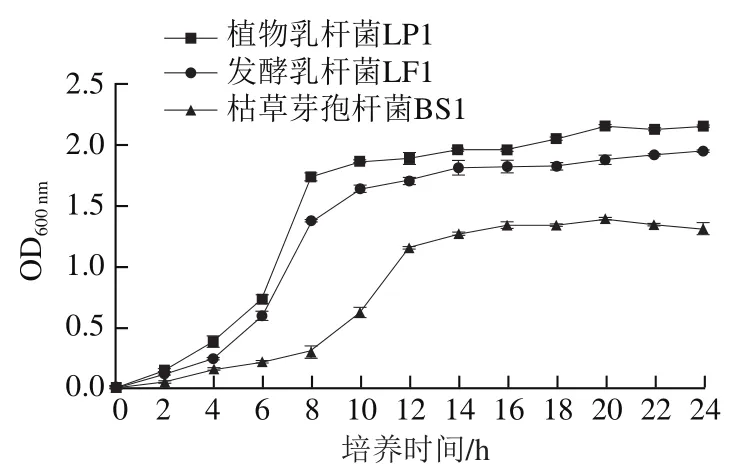
圖1 不同菌種的生長曲線Fig. 1 Growth curves of different strains
2.1.2 不同菌種的產酸曲線
由圖2可知,植物乳桿菌LP1和發酵乳桿菌LF1的產酸曲線也具有相似的趨勢,與圖1菌株生長曲線所體現的趨勢相符。經過延滯期對培養環境的適應,2 株乳桿菌培養基的pH值在生長4 h后迅速下降,進入對數期后代謝旺盛,開始大量產酸,生長8 h后進入穩定期,細胞代謝速率下降,并且已產生的酸類物質會對菌株的生長起到抑制作用,最終植物乳桿菌LP1的pH值穩定在3.7左右,發酵乳桿菌LF1的pH值穩定在4.0左右。與以上2 株乳桿菌相比而言,枯草芽孢桿菌BS1對數生長期培養基pH值下降的速率明顯降低,且最終pH值穩定在4.7左右。可以看出,3 種菌株中2 株乳桿菌的產酸能力遠高于枯草芽孢桿菌BS1,其中植物乳桿菌LP1的產酸能力最強。因此,綜合生長速率和產酸能力的結果,本研究最終選擇植物乳桿菌LP1作為發酵雞肝的發酵劑。
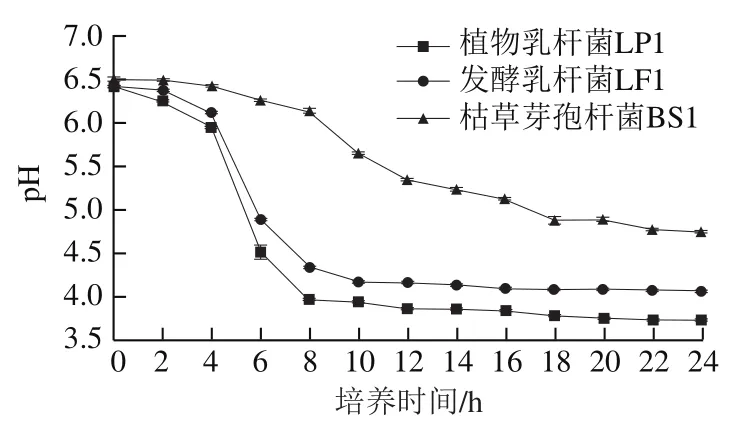
圖2 不同菌種的產酸曲線Fig. 2 Acid production curves of different strains
2.2 單因素試驗結果
2.2.1 發酵初始pH值對的影響
由圖3可知,隨著發酵初始pH值的增加總體呈現先升高后降低的趨勢,在發酵初始pH值為5.5時達到最高(<0.05),說明在該pH值條件下發酵雞肝中小分子肽和游離氨基酸的含量最高。部分低分子質量肽和游離氨基酸可以單獨呈味,也可以繼續反應形成其他風味化合物。雞肝蛋白質在植物乳桿菌LP1分泌的蛋白酶作用下降解,研究表明,生成的低分子質量肽和游離氨基酸不僅能起呈味增鮮作用,而且具有一定生理活性,能改善雞肝的風味,提高營養價值。與鄧愛華等優化地衣芽孢桿菌SD1482發酵魷魚下腳料時得到的結果一致,最佳發酵初始pH值同樣為5.5。研究發現,發酵初始pH值不僅會影響菌株的生長情況,還會影響酶的活性。為了促進發酵過程中菌株的生長,增強酶的活性,需要選擇合適的初始pH值。因此,本研究選取發酵初始pH 5.5為最優條件進行響應面試驗。
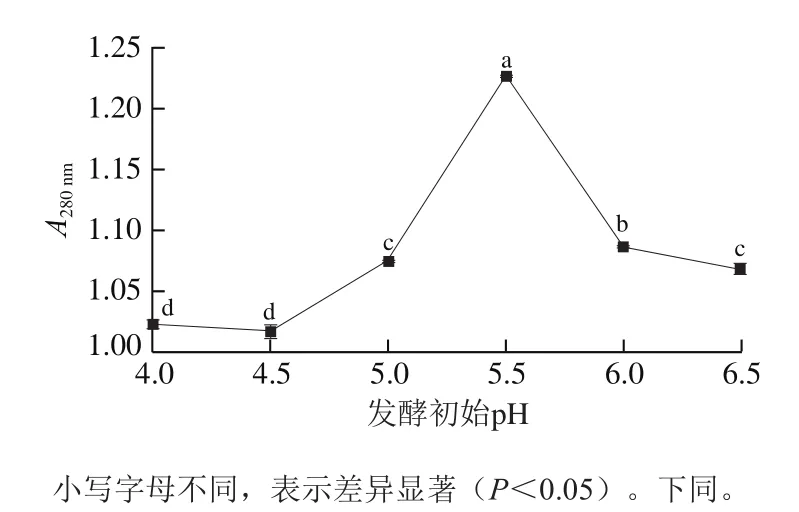
圖3 發酵初始pH值對A280 nm的影響Fig. 3 Effect of initial pH of fermentation medium on A280 nm
2.2.2 葡萄糖添加量對的影響
葡萄糖是乳酸菌生長繁殖必不可缺的碳源,而雞肝中的糖類以肝糖原形式存在,葡萄糖較之更有利于乳酸菌的利用,因此本實驗選擇向雞肝發酵培養基中添加葡萄糖。乳酸菌可將培養基中的碳源分解轉化為乳酸等有機酸,賦予發酵產品較強的酸味,這些酸類還可與醇類反應生成酯類物質,對產品特征風味的形成具有重要貢獻。
由圖4可知,首先隨著葡萄糖添加量增加而增大,在葡萄糖添加量為8%時達到最大值,隨后下降。葡萄糖添加量低于8%時,因碳源量無法滿足植物乳桿菌LP1正常生長所需而導致菌株生長速率緩慢,進而造成蛋白酶產量較低,蛋白質分解程度較低,較低。葡萄糖添加量繼續增加,出現下降,這是由于培養基中過高的葡萄糖含量會造成滲透壓過高,可能破壞菌株細胞結構,細胞的正常生理活動和代謝過程被擾亂,甚至導致菌體死亡,不利于發酵的進行,故選擇葡萄糖添加量為6%進行發酵。
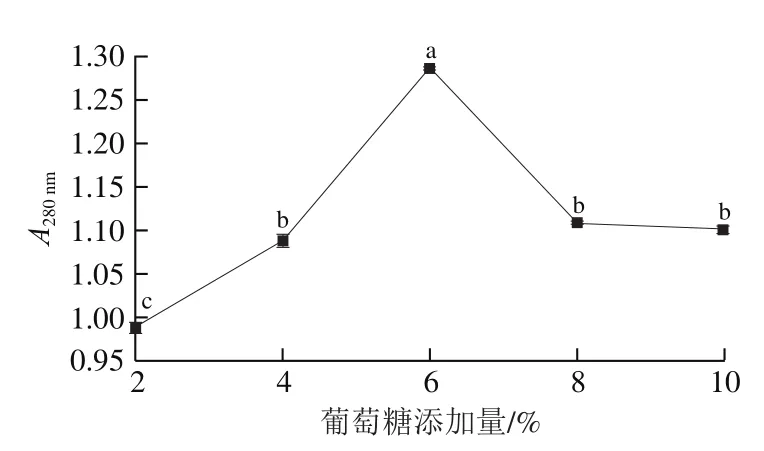
圖4 葡萄糖添加量對A280 nm的影響Fig. 4 Effect of glucose addition on A280 nm
2.2.3 接種量對的影響
由圖5可知:接種量在0.5%~1.5%時,呈上升趨勢;當接種量大于1.5%時,開始下降。這是因為在接種量過低時,菌株生長繁殖速率慢,植物乳桿菌LP1分泌的蛋白酶含量很少,分解蛋白質的能力較弱,故小肽及游離氨基酸的含量較低;而當接種量過大時,菌株迅速繁殖,菌體密度過大會導致與氧氣的接觸不足,不利于發酵的進行,另外,發酵劑用量過高不利于控制生產成本。同時植物乳桿菌LP1發酵產生大量的有機酸,使培養基處于較高的酸度環境,反而會抑制菌體部分代謝活動,影響產酶量及酶活性,從而導致接種量過大時發酵雞肝中小分子肽和氨基酸的含量隨之降低。因此,選擇接種量為1.5%進行發酵。李梓媛在研究乳酸菌和肉葡萄球菌混菌發酵制備羊肉香腸時,發現以8%的接種量進行發酵,菌株的產酸能力最強,有利于保證發酵香腸的安全性,使發酵香腸具有穩定的品質。
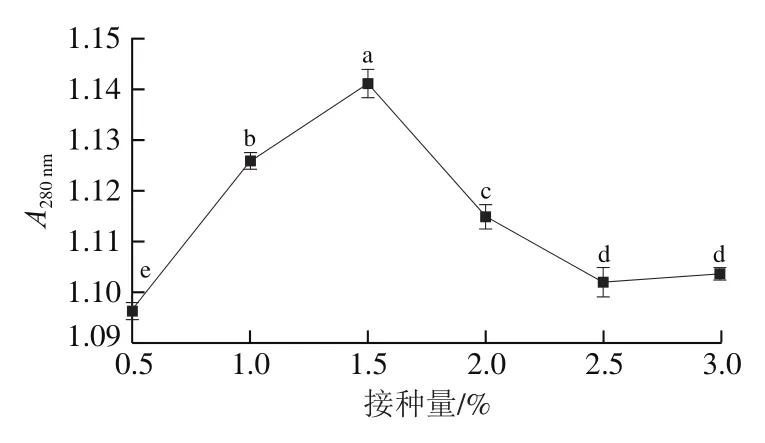
圖5 接種量對A280 nm的影響Fig. 5 Effect of inoculum size on A280 nm
2.2.4 發酵時間對的影響
由圖6可知,發酵16~24 h內,發酵雞肝的隨著發酵時間的延長而持續上升,20~24 h的急劇上升,說明大量的雞肝蛋白質被分解利用,小肽及游離氨基酸的含量迅速增加,可能由于在發酵24 h時植物乳桿菌LP1分泌的蛋白酶含量及酶活達到最大值,此時的發酵效果顯著。當發酵時間超過24 h,不再升高,隨著時間的延長甚至還出現輕微下降,當大部分雞肝蛋白被分解后,小分子肽和游離氨基酸的含量遠高于雞肝蛋白的含量,此時植物乳桿菌LP1會優先消耗小分子物質,造成小肽及游離氨基酸含量下降。另外,過長時間的發酵會導致乳酸等物質的積累,菌體的產酶量在進入平穩期后達到最大值,繼續發酵可能會影響蛋白酶的產量和活性。因此,綜合考慮生產成本以及避免過長時間發酵導致發酵效果的降低,選擇發酵時間為24 h。
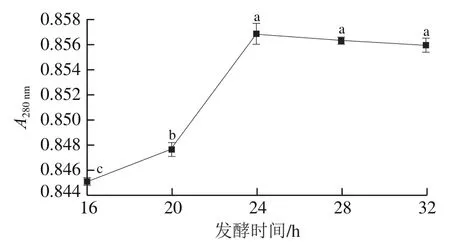
圖6 發酵時間對A280 nm的影響Fig. 6 Effect of fermentation time on A280 nm
2.2.5 料液比對的影響
雞肝作為植物乳桿菌生長繁殖的主要營養基質,控制合適的料液比能在節省生產成本的同時提高植物乳桿菌的生長速率。由圖7可知,料液比1∶2.0~1∶3.5時,隨著液體添加量的增加顯著增高(<0.05),在料液比為1∶3.5時達到最高,隨著液體添加量的繼續增加,開始下降,料液比過高會使生產成本提高,令狐青青等使用納豆芽孢桿菌()發酵魷魚碎肉,發現料液比過高會導致發酵液中氨基酸態氮含量下降,與本研究結果趨勢一致。當料液比達到1∶4.5時,與料液比為1∶4.0時相較無顯著變化,說明在料液比1∶4.0~1∶4.5范圍內,雞肝蛋白被分解生成小分子肽和游離氨基酸的量與被植物乳桿菌LP1菌體利用的量保持相對平衡,考慮到生產成本,選擇料液比1∶3.5進行下一步發酵工藝的優化。
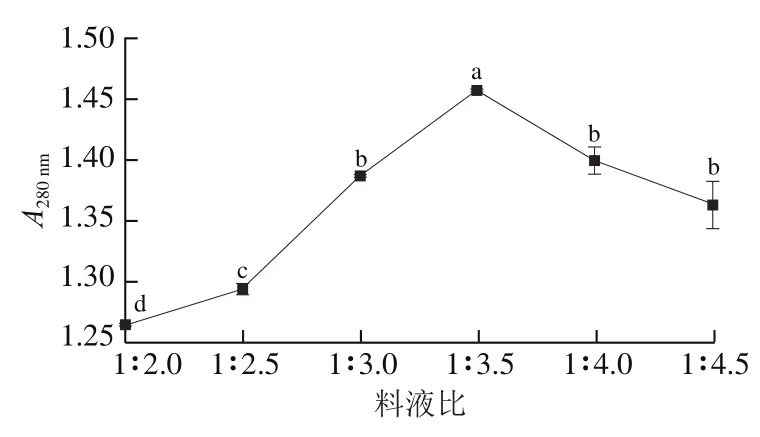
圖7 料液比對A280 nm的影響Fig. 7 Effect of solid-to-liquid ratio on A280 nm
2.3 響應面優化試驗結果
根據單因素試驗結果,接種量、發酵初始pH值和料液比對雞肝發酵的影響更為顯著,因此選擇該3 個因素對雞肝發酵工藝進行優化。
2.3.1 數學模型的建立及顯著性檢驗
利用Design-Expert 8.0.6軟件對表2響應面試驗數據進行回歸分析,得到與接種量()、發酵初始pH值()、料液比()3 個因素間的回歸方程為:=-21.196 47+2.477 85+5.206 20+3.608 65-0.120 00-0.181 00+0.086 00-0.389 70-0.478 70-0.537 70。該方程中各項系數的絕對值大小代表各因素對響應值的影響程度,系數的正負則反映各因素對響應值影響的方向。由一次項系數可以得出對于發酵物的影響程度依次為>>,即發酵初始pH值>料液比>接種量。

表2 響應面試驗結果Table 2 Experimental design and results for response surface analysis
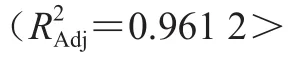
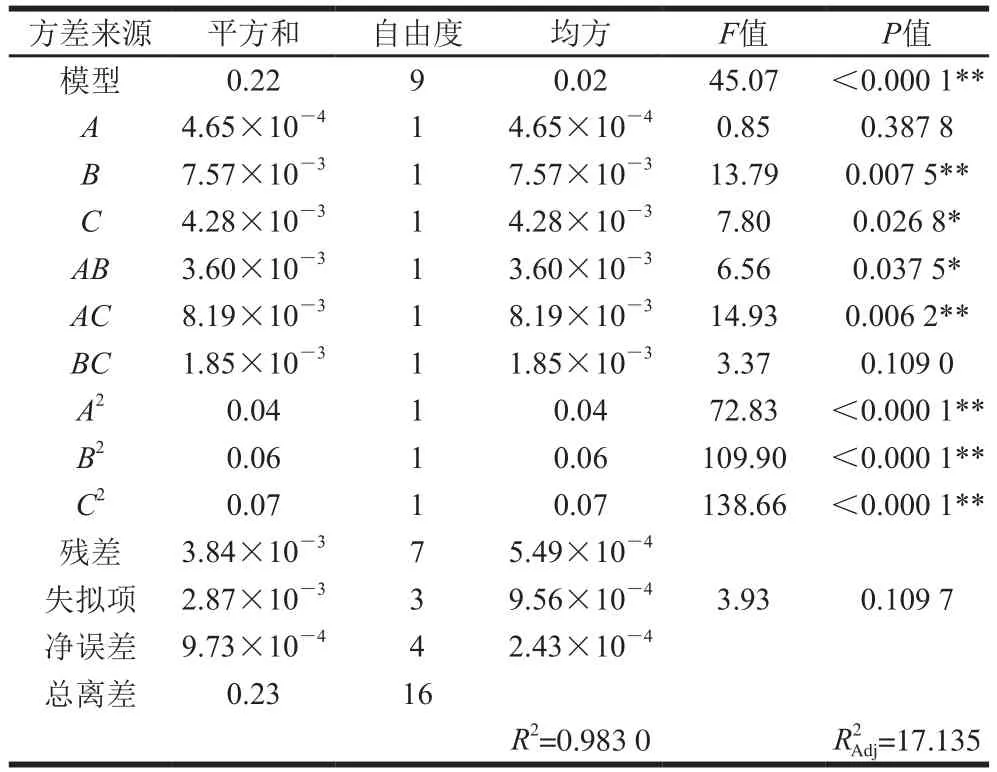
表3 回歸模型顯著性系數檢驗Table 3 Analysis variance of quadratic polynomial model and significance test
通過顯著性分析可知,模型中一次項中的發酵初始pH值影響極顯著(<0.01),料液比影響顯著(<0.05),接種量影響不顯著;二次項系數均影響極顯著(<0.01);交互項中,接種量與料液比交互作用極顯著(<0.01),接種量與發酵初始pH值的交互作用顯著(<0.05),發酵初始pH值與料液比交互作用不顯著。
2.3.2 因素交互作用分析
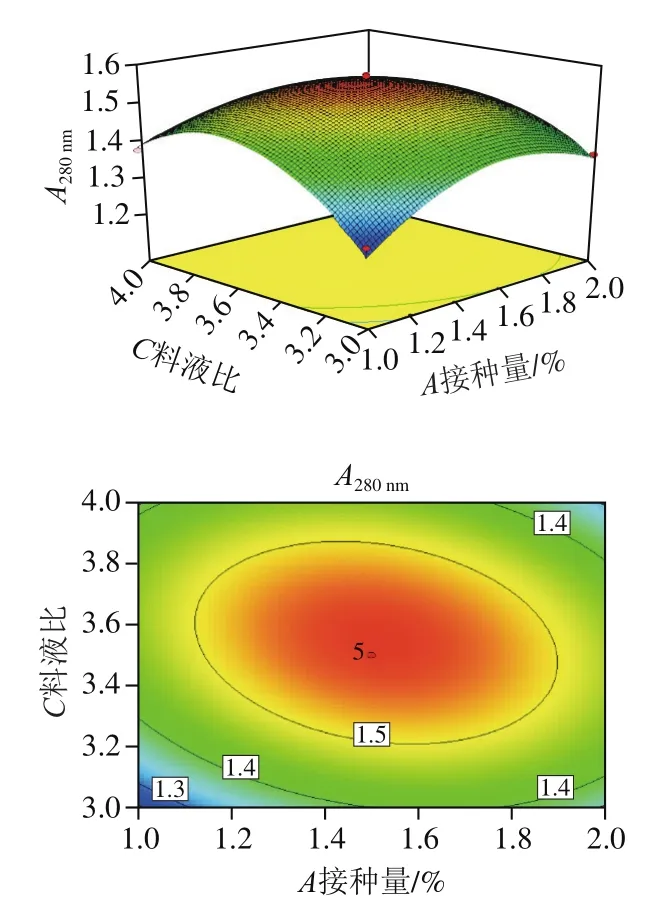
圖9 接種量和料液比對A280 nm影響的響應面圖及等高線圖Fig. 9 Response surface and contour plots showing the effects of inoculum size and solid-to-liquid ratio on A280 nm
分析接種量、發酵初始pH值、料液比中任意一個因素為零水平時,其余2 個因素對發酵雞肝的交互影響。為了分析兩因素間交互作用的程度,需要通過響應面陡峭程度、三維等高線的形狀來進行直觀反映。其中響應值與各自變量之間的變化程度可通過響應面坡度的陡峭情況來分析。等高線的形狀呈橢圓形,表明兩因素之間的交互作用顯著,而等高線趨于圓形則說明交互作用較弱。可以看出,圖8~9的響應面具有較陡峭的坡面,等高線呈現橢圓形,說明接種量和發酵初始pH值、接種量和料液比的交互作用對發酵雞肝有顯著影響。圖10響應面坡面較緩,等高線形狀趨于圓形,可知發酵初始pH值和料液比的交互作用對發酵雞肝的影響不大,這與表3方差分析的結果一致。

圖8 接種量和發酵初始pH值對A280 nm影響的響應面圖及等高線圖Fig. 8 Response surface and contour plots showing the effects of inoculum size and initial pH of fermentation medium on A280 nm

圖10 發酵初始pH值和料液比對發酵雞肝A280 nm影響的響應面圖及等高線圖Fig. 10 Response surface and contour plots showing the effects of initial pH of fermentation meduim and solid-to-liquid ratio on A280 nm
2.3.3 雞肝發酵工藝參數的優化和模型驗證
以為響應值,通過Design Expert 8.0.6軟件分析得到植物乳桿菌LP1發酵雞肝的最佳工藝條件為:發酵初始pH 5.57、料液比1∶3.55、接種量1.5%。在此條件下,發酵物的預測值為1.559。經過3 次驗證實驗(實驗條件保持高度一致),得到發酵雞肝的實際值為1.537,與預測值的吻合度達到98.59%,因此,該模型能夠有效應用于植物乳桿菌LP1發酵雞肝工藝的優化。
2.4 最優工藝條件下發酵雞肝游離氨基酸組成測定結果
植物乳桿菌LP1在發酵過程中分泌的蛋白酶可將雞肝蛋白分解為小分子質量的肽和大量游離氨基酸,這些小肽和游離氨基酸本身具有呈味作用,同時還可與其他成分進一步反應作為風味化合物的前體物質,對發酵雞肝風味的形成有重要作用。通過對游離氨基酸組成的測定分析,可知發酵前后雞肝中游離氨基酸的種類及變化情況。測得的17 種氨基酸中,根據機體是否合成及合成速率是否滿足需求,分為必需氨基酸(Thr、Val、Met、Ile、Leu、Phe、Lys、His)和非必需氨基酸(Asp、Ser、Glu、Gly、Ala、Cys、Tyr、Arg、Pro);按照呈味不同可分為鮮味氨基酸(Asp、Glu)、甜味氨基酸(Ser、Ala、Thr、Pro、Gly)和苦味氨基酸(Val、Leu、Ile、Tyr、Lys、Arg、Phe)等。
由表4可知,發酵雞肝的游離氨基酸總含量顯著高于未發酵雞肝(<0.05),同時必需氨基酸的總含量經過發酵后顯著增加(<0.05),說明發酵能促使蛋白質分解為大量游離氨基酸,其中蘇氨酸、異亮氨酸、賴氨酸、組氨酸、天冬氨酸、絲氨酸和精氨酸含量分別提高58.67%、49.18%、65.71%、51.62%、57.39%、80.82%和53.93%,從而提高雞肝的營養價值。Li Chunsheng等使用戊糖片球菌30-7和30-15發酵羅非魚香腸,發現戊糖片球菌30-15能夠顯著提高羅非魚香腸中鮮味和甜味游離氨基酸的含量,改善產品滋味。

表4 發酵前后雞肝游離氨基酸組分含量的變化Table 4 Changes in free amino acid composition of chicken liver before and after fermentation g/100 g
3 結 論
通過對植物乳桿菌LP1、發酵乳桿菌LF1和枯草芽孢桿菌BS1生長曲線及產酸曲線的測定,得出植物乳桿菌LP1的生長速率最快、產酸能力最強,故選擇植物乳桿菌LP1作為發酵劑對雞肝進行單菌發酵,同時可利用植物乳桿菌LP1產酸抑制雜菌的生長,提高安全性。以單因素試驗為基礎,選擇發酵初始pH值、接種量和料液比進行3因素3水平響應面試驗,對植物乳桿菌LP1發酵雞肝的工藝進行優化,得到最優工藝參數為:發酵初始pH 5.57、料液比1∶3.55、接種量1.5%。結果顯示,雞肝發酵后游離氨基酸和必需氨基酸總含量均顯著增加(<0.05),進一步說明發酵可促進蛋白質分解為大量游離氨基酸,提高雞肝的營養價值。發酵雞肝是制備寵物貓食品的優質原料,拓展了雞肝副產物綜合利用的新途徑。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
