哀牢山紫莖澤蘭入侵群落主要物種種間聯結及環境解釋
2022-09-02 08:46:30姜倪皓張世浩張詩函
生態環境學報 2022年7期
姜倪皓,張世浩,張詩函
1.楚雄師范學院資源環境與化學學院,云南 楚雄 675000;2.河池學院化學與生物工程學院,廣西 河池 546300;3.楚雄師范學院教育學院,云南 楚雄 675000
種間關系是生態學研究的重要內容。種間聯結是不同物種在空間分布上的相互關聯性,用以描述由于群落生境差異造成的不同物種分布的相互關系,它是植物群落重要的數量和結構特征之一,是群落結構形成和演化的重要基礎(Haukisalmi et al.,1998)。研究群落內主要物種的種間聯結特征,有助于正確認識群落的組成和功能及演替趨勢,對植物資源的保護和利用、生物多樣性保護以及植被恢復與重建等具有重要的意義(劉海等,2017;江煥等,2019)。采用種間聯結性等數量生態學方法探討生物入侵后群落中物種間的聯結性是研究入侵生物和本土物種間相互作用的重要切入點,當前國內對外來植物入侵群落的種間聯結的報道相對較少(郭連金等,2009;郭佳琦等,2021)。在群落尺度上,微地形因子和土壤養分,水分等微生境因子的差異及它們之間的協同作用會導致生境異質性,進而影響植物的空間分布(謝玉彬等,2012;吳昊,2015),研究物種種間聯結性及物種分布與環境的關系有助于了解優勢物種對資源利用的生態習性及其相互關系,也有助于揭示群落的水平空間配置情況和物種分布的主導環境因子,對生態系統的科學經營管理等具有重要意義(陳佳卉等,2021)。
紫莖澤蘭(Ageratinaadenophora)是菊科(Compositae)紫莖澤蘭屬(AgeratinaSpach)的多年生叢生型半灌木狀草本植物,是中國外來入侵物種中危害最嚴重的植物之一(Wang et al.,2006;Gui et al.,2008)。紫莖澤蘭入侵影響生物群落的結構與功能,打破了群落內物種共存的生態格局(陳旭等,2021),給入侵地區的生態系統和環境造成了嚴重的危害。哀牢山位于中國西南部云南省的云貴高原、橫斷山脈和西藏高原之間,是云南重要的氣候與生態類型的分界線。該區域擁有典型的亞熱帶森林生態系統,生物多樣性較豐富,植物區系地理成分復雜(王高升等,2008)。筆者在哀牢山東麓的野外調查中發現,紫莖澤蘭早已入侵該區域并在該區域有迅速擴散蔓延的趨勢,這將造成該區域植物群落物種多樣性喪失、自然生態系統服務功能下降等破壞性影響,嚴重威脅該區域生態系統的穩定性。
目前,關于紫莖澤蘭入侵群落植物種間聯結性及物種分布與環境的關系的研究仍相對缺乏(劉海等,2017),尤其針對哀牢山地區紫莖澤蘭群落尚無相關的研究報道。本研究以哀牢山紫莖澤蘭群落為研究對象,利用χ2檢驗、方差比率法、種間聯結系數、Spearman秩相關系數檢驗等數量分析方法,對研究區紫莖澤蘭入侵群落主要植物的種間聯結性進行定量分析。采用冗余分析(RDA),方差分解,隨機森林模型和結構方程模型共同探究環境因子對主要物種分布及入侵的影響。旨在探究以下3個科學問題:(1)紫莖澤蘭與伴生物種的種間關系和相互影響如何?(2)影響群落物種分布的主要環境因子是什么?(3)影響入侵植物入侵強度指數、群落可入侵性指數和群落穩定性指數的主要環境因子是什么?這些科學問題的合理解答將為該區域紫莖澤蘭入侵后植物群落的管理以及本土生物多樣性保護提供理論依據。
1 研究區概況和研究方法
1.1 研究區域概況
雙柏縣處于滇中干旱地區,屬環境敏感區域和生態環境脆弱地區,同時雙柏縣亦是哀牢山國家級自然保護區的核心區域。本研究區位于哀牢山北段東麓的雙柏縣愛尼山鄉安龍哨森林管護點所轄的林區,地理坐標為 101°56′—101°57′E、24°44′—24°45′N(圖1)。該區域屬北亞熱帶高原季風氣候,年平均降雨量927 mm,年平均氣溫15 ℃。土壤類型以紅壤為主。
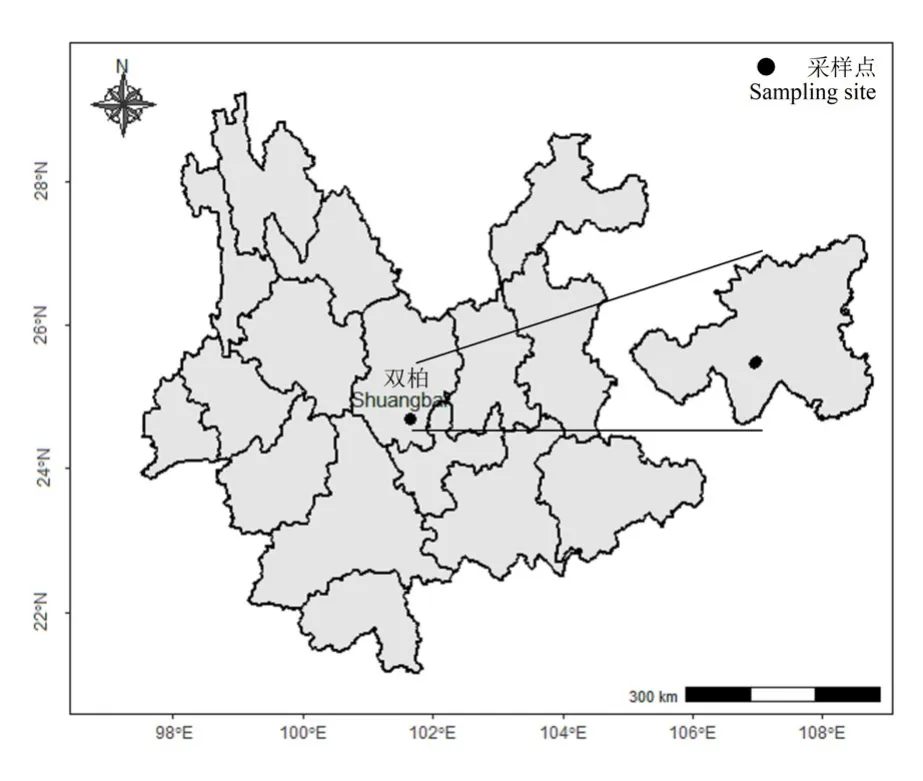
圖1 研究區位置Figure 1 Location of study area
研究區域的喬木層以云南松(Pinus yunnanensis)為主,混生少量滇木荷(Schimanoronhae)和麻櫟(Quercusacutissima)。該區域植被長期維持自然生長發育狀態,對紫莖澤蘭入侵也未采取任何防控措施,紫莖澤蘭呈現蔓延的趨勢。
1.2 研究方法
1.2.1 樣地設置與調查
2020年7月,在紫莖澤蘭典型分布區域內(相對豐度>50%),設置垂直于群落邊緣的3條垂直樣帶,每條樣帶內設置10個20 m×20 m的標準樣地,測定每個標準樣地內胸徑大于1 cm的立木的種類、數量、基徑、胸徑、樹高等因子,記錄郁閉度、海拔等群落和環境因子信息。同時在每個樣地內分別沿對角線設置3個1 m×1 m的樣方,在90個樣方內調查紫莖澤蘭及其他灌草層物種的株數、株高和蓋度等特征。坡向的原始記錄是以朝東為起點(即為0),順時針旋轉的角度表示。坡向數據處理時,采取每45°為1個區間的劃分等級制的方法,數字越大,表示越向陽、越干熱(張峰等,2003)。
1.2.2 土壤理化性狀測定
采集每個小樣方中心0—20 cm土層的土壤樣品,將每個樣地中3個小樣方土壤混合,土樣過0.25 mm篩后保存,土壤理化性質分析均為常規分析方法(黃慶陽等,2020)。土壤含水量采用烘干法。土壤容重采用環刀法。采用電位法(水土比為2.5∶1)測量土壤pH值。電導率由手持電導率儀測定。有機質含量測定用重鉻酸鉀容量法-外加熱法測定。全氮測定采用半微量開氏法。全磷測定采用鉬銻抗分光光度法。全鉀測定采用原子吸收分光光度法。土壤中有效氮含量測定采用堿解擴散法。有效磷測定采用紫外/可見分光光度計。有效鉀測定采用 1 mol·L-1乙酸銨浸提法。
1.2.3 重要值及生態位特征測定
采用重要值反映物種在群落中的優勢程度,剔除偶見種后,基于相對頻度、相對多度、相對密度和相對高度的相關數據計算重要值,取相對重要值大于 1.0且出現頻率最高的 10個物種進行后續分析。重要值計算公式(張金屯,2018)如下:

式中:
B(sw)i——物種i的Shannon-Wiener生態位寬度指數,值域為[0, lnr];
Oik——物種i和k的生態位重疊指數;
Cik——物種i與物種k的生態位相似比例,Cik值域為[0, 1];
Pij——物種i在第j資源位上的重要值;
Pkj——物種k在第j資源位上的重要值。
1.2.4 總體聯結性檢驗
采用零聯結模型導出方差比率(Rv)法檢驗多物種間的總體關聯性(Schluter,1984;張金屯,2018)。Rv>1表示物種間總體上表現為正關聯,Rv<1表示物種間總體上表現為負關聯。采用統計量(W)來檢驗多物種間的關聯程度,χ20.95<W<χ20.05表示關聯不顯著,反之則表示關聯顯著。計算公式為:
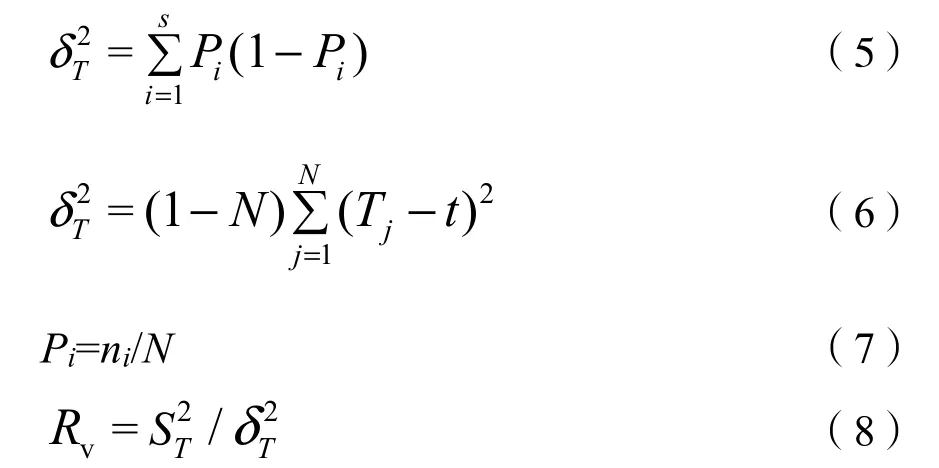
式中:
Pi——物種i出現的頻率;
S——物種總數;
N——總樣方數;
Tj——樣方j內出現的物種數;
t——各樣方內物種數的平均值;
ni——物種i出現的樣方數。以Rv值作為群落內主要物種間的總體聯結指數,在獨立性零假設條件下,Rv的期望值為1。Rv>1表示物種間總體上表現為正關聯,Rv<1表示物種間總體上表現為負關聯。采用統計量(W)來檢驗多物種間的關聯程度,W=N×Rv;χ20.95<W<χ20.05表示關聯不顯著,反之則表示關聯顯著。
1.2.5 種間聯結性檢驗
采用χ2統計量檢驗對種間聯結性進行定性研究,應用SPSS 19.0計算Yates連續性校正χ2值。Yates的連續性校正公式如下(張金屯,2018):

式中:
a——2個物種共同出現的樣方數;
b、c——僅有1個物種出現的樣方數;
d——2個物種均出現的樣方數。當χ2<3.841,即P>0.05,種間聯結性不顯著;當 3.841<χ2<6.635,即0.01<P<0.05,種間聯結性顯著;當χ2>6.635時,即P<0.01,認為種間聯結性極顯著。χ2值本身沒有負值,本研究使用V比值判斷種間關聯性。V=(a+b)-(b+c)/(a+b+c+d),當V>0時,種間關聯性為正聯結;當V<0時,種間關聯性為負聯結。
1.2.6 種間關聯程度測定
采用聯結系數Ca對χ2統計量檢驗結果進行進一步檢驗。計算公式如下(張金屯等,2018):當ad≥bc時,Ca=(ad-bc)/[(a+b)(b+c)];當ad<bc時,且d≥a時,Ca=(ad-bc)/[(a+b)(a+c)];當ad<bc時,且d<a時,Ca=(ad-bc)/[(b+d)(d+c)];Ca值域為[-1,1],Ca值越趨向1,表明種間正聯結性越強;Ca值越趨近-1,表明種間負聯結性越強;Ca值為0,表明種間相對獨立。
1.2.7 種間關聯程度測定
采用優勢種的重要值作為 Spearman秩相關分析的數量指標。Spearman秩相關系數計算公式(張金屯,2018):

式中:
n——樣方數;
xik和xjk——種i和種j在樣方k中的秩。
rs(i,j)——值域為[-1, 1],正值表示正相關,負值表示負相關。
1.2.8 入侵植物入侵強度指數及群落可入侵性指數
采用入侵植物入侵強度指數(Invasion intensity index of alien invasive plants,Iii)表征入侵植物入侵強度,Iii值越大表示入侵植物的入侵強度越高(Wang et al.,2020)。采用群落可入侵性指數(Community invasibility index,Cii)表征群落可入侵性水平,Cii值越大表示群落可入侵性越高(Wang et al.,2020)。
入侵植物入侵強度指數(Iii),計算公式如下:

群落可入侵性指數(Cii),計算公式如下:

式中:
Pimax——所有調查樣方內入侵植物相對豐度最大值;
Pi——調查樣方內入侵植物的相對豐度。Cii值越大表示群落可入侵性越高。
1.2.9 群落穩定性
群落穩定性采用以下兩種計算方法:
(1)群落穩定性指數(community stability index)以物種種群密度變異系數(coefficient of variation,CV)的倒數ICV表示,計算公式如下(Wang et al.,2020):

式中:
μ——調查樣方內各植物種類的平均密度;
σ——調查樣方內各植物種類密度的標準差(SD)。ICV值越大表示群落穩定性越高。
(2)使用鄭元潤(2000)改進后的M.Godron穩定性測定方法測定紫莖澤蘭入侵群落的穩定性。先計算出群落中主要物種的頻度,按相對頻度的大小順序逐步累積,與主要物種總物種數的倒數累積一一對應,建立模糊散點平滑曲線模型。繪制經過(0, 100)、(100, 0)的直線,該線段與平滑曲線的交點即為所求解。交點坐標越接近(20, 80),表示群落越穩定。
1.2.10 統計分析
以優勢植物相對重要值為基礎,利用 Canoco 5.0軟件對物種重要值矩陣進行去趨勢對應分析(detrended correspondence analysis,DCA),DCA排序前 4個軸中最大值為 2.7。然后進行冗余分析(RDA),通過前向選擇(forward selection)法和蒙特卡洛檢驗(Monte Carlo test,循環次數499)篩選影響顯著(P<0.05)的環境因子,探討環境因子對物種分布的影響。基于約束性排序結果,通過多元線性逐步回歸方法分析環境因子對群落穩定性、群落可入侵性及入侵植物入侵強度的相對影響,影響程度以標準化的偏回歸系數確定。
基于R(4.1.2)軟件,運用“Vegan”數據包里的“Varpart”功能進行方差分解分析。為了驗證逐步回歸分析結果,利用隨機森林模型分析環境因子對群落穩定性、群落可入侵性及入侵植物入侵強度的相對重要性,為了判斷隨機森林模型的可靠性,對每個模型進行十折交叉驗證。
入侵植物與環境因子間的關系較復雜,為厘清各環境因子的貢獻,引入結構方程模型分析各環境因子對紫莖澤蘭入侵的影響。根據已知的土壤因子與紫莖澤蘭入侵及群落穩定性之間的因果關系,借助Amos 7.0軟件構建初始模型。選用擬合指數(GFI>0.9)、漸進殘差均方和平方根(RMSEA<0.08)、顯著性概率值(P>0.05)等指標評價模型優度(王酉石等,2011)。最終選取全磷、有效鉀、土壤pH值、土壤溫度作為土壤因子,構建出最優結構方程模型。利用模型中各路徑標準化系數,定量表示不同因素對的相對群落穩定性指數、群落可入侵性指數及入侵植物入侵強度指數的相對作用大小。
2 結果與分析
2.1 紫莖澤蘭與伴生物種的生態位分析
根據 90個樣方統計資料,紫莖澤蘭入侵群落共有29種植物。其中,喬木17種,灌木4種,草本植物6種,蕨類植物2種,隸屬21科27屬。本研究選取了群落中重要值大于1.0且出現頻率最高的10個物種作為主要物種(表1)。重要值排在前3位的物種依次為紫莖澤蘭(Ageratinaadenophora)(28.90%)、里白(Diplopterygiumglaucum)(22.31%)和虎杖(Reynoutriajaponica)(7.89%)。生態位寬度較大的物種依次為紫莖澤蘭(0.79)、里白(0.76)和芒萁(Dicranopterisdichotoma)(0.59)。
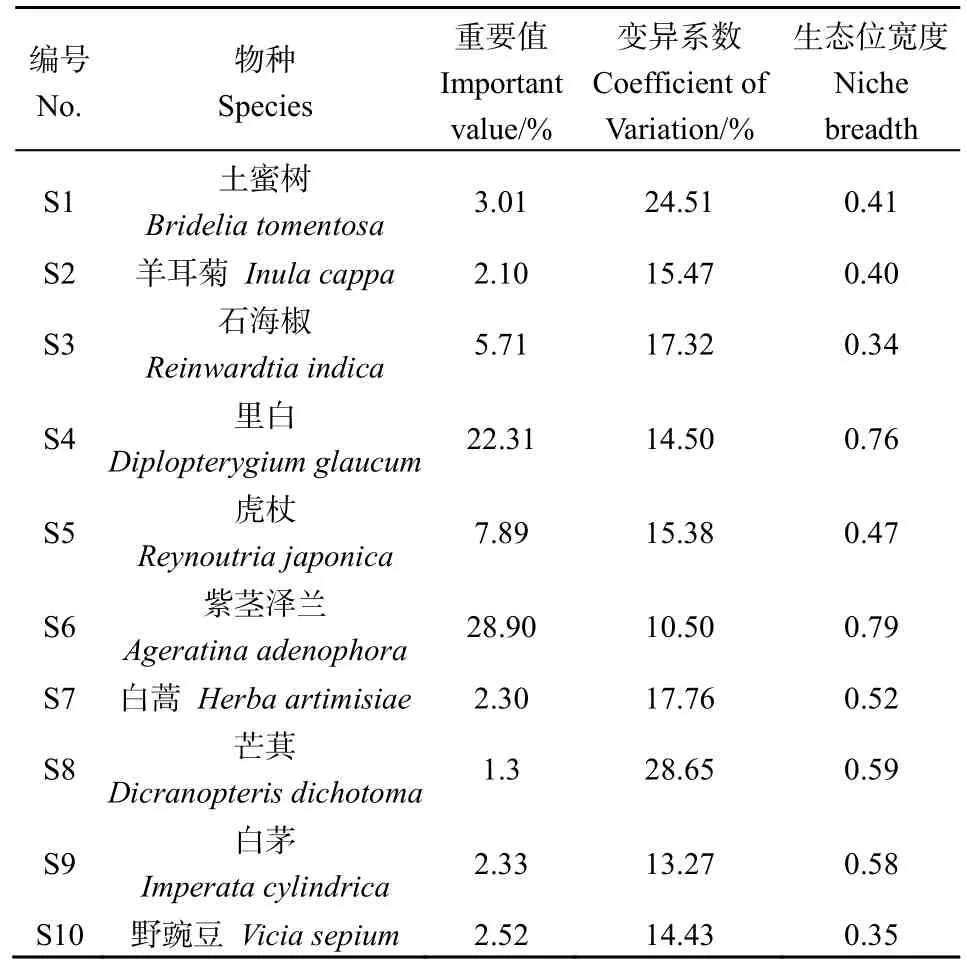
表1 草本層優勢種重要值及其生態位寬度Table 1 Importance value and niche breadth of dominant species in herbaceous layer
生態位相似比例能夠反映同一群落的不同種群對環境資源利用的相似性和物種生物學特性。紫莖澤蘭群落主要物種的生態位相似性系數平均值為 0.312,與紫莖澤蘭生態位相似性最高的為里白(Cik=0.642)。生態位相似性系數為0的種對有19個(42.22%),生態位相似性系數在 (0, 0.5]區間的種對共11個(24.44%)(表2),表明大部分物種對環境資源的需求有較大差異。
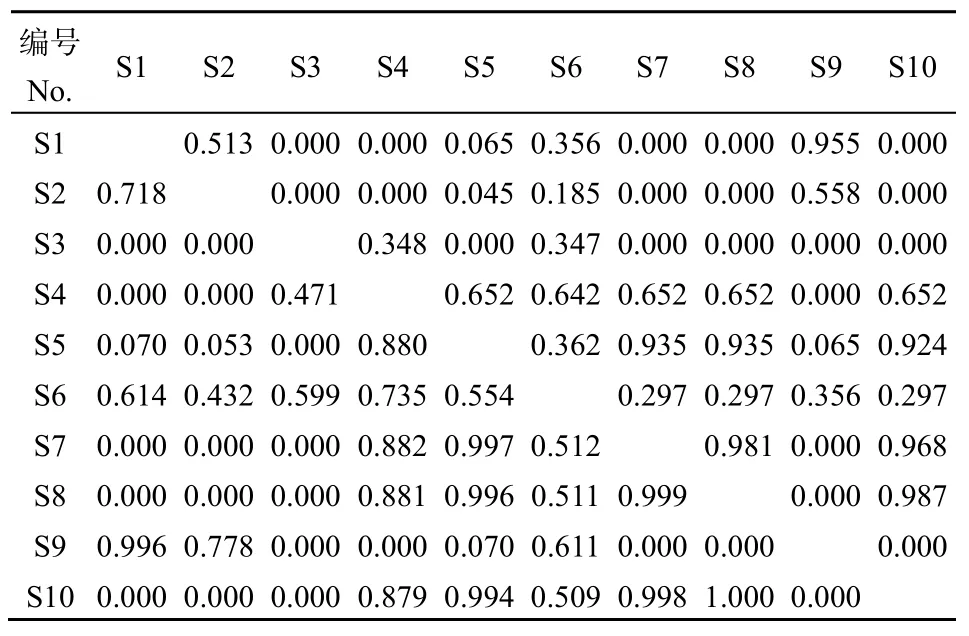
表2 紫莖澤蘭群落主要優勢種間的生態位相似性比例(對角線上)和生態位重疊指數(對角線下)Table 2 Niche similarity (above the diagonal) and niche overlap (below the diagonal) of dominant plant species in Ageratina adenophora community
紫莖澤蘭群落主要物種的生態位重疊指數的范圍在 (0, 1]區間,生態位重疊指數平均值為0.394。與紫莖澤蘭生態位重疊最高的為里白(Oik=0.735)。生態位重疊指數在0.5以上的種對有21對,占總數的46.67%(表2),且其中8對發生在紫莖澤蘭與伴生植物間,這些種對在環境資源不足時會產生激烈的競爭。
2.2 群落總體聯結性和種間聯結分析
紫莖澤蘭群落主要物種間的總體聯結性計算結果如表3所示,總體聯結性的方差比率Rv=0.181<1,表明主要物種間總體呈負聯結狀態。而檢驗統計量(W)未落入界限χ20.95<W<χ20.05,表明Rv顯著偏離1,即多物種間整體負聯結具有統計學意義。研究表明,哀牢山紫莖澤蘭群落主要物種間的總體聯結性表現為顯著負聯結。
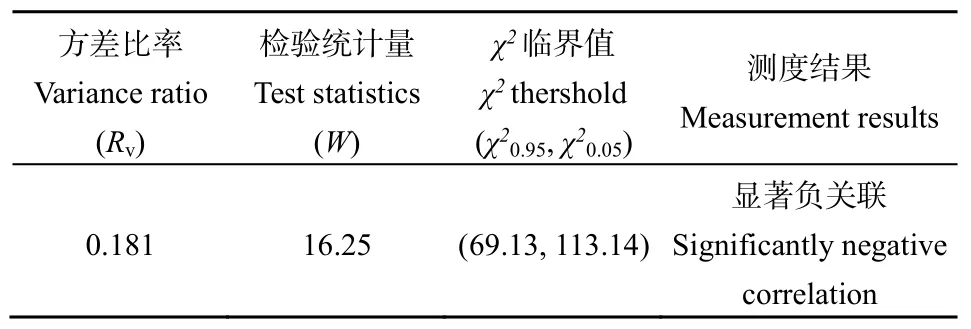
表3 主要物種間的總體聯結性Table 3 Overall interspecific associations among dominant species
χ2檢驗結果表明(圖2A),在10個主要物種構成45個種對,呈不顯著正聯結的種對有7對,占總種對數的 15.56%;11個種對呈極顯著或顯著正聯結,占總種對數的24.44%;呈不顯著負聯結的種對有15對,占總種對數的33.33%;呈極顯著或顯著負聯結的種對有12對,占總種對數的26.67%。正負聯結種對的比值為0.67,負聯結占優勢,這與群落整體關聯性呈負關聯的結果相一致。聯結系數Ca(圖2B)和Spearman秩相關分析(圖2C)結果的正負聯結比分別為0.67和0.73,表明種對間負聯結占據相對優勢地位,優勢種間聯結比較松散,各自趨向于獨立分布。
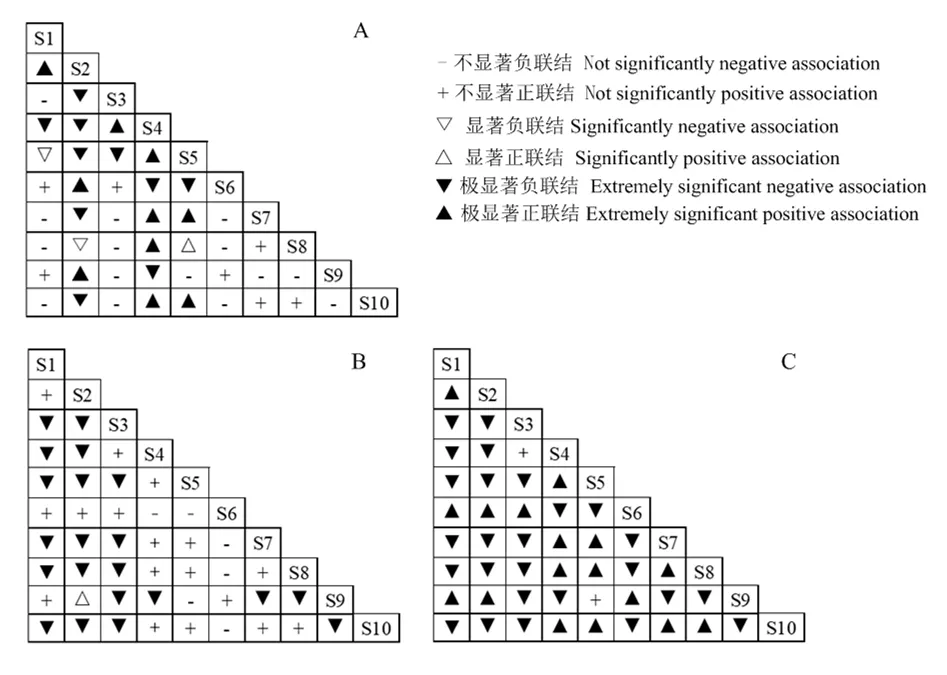
圖2 主要物種的χ2檢驗(A)、聯結系數(Cr)(B)和Spearman秩相關分析(C)半矩陣圖Figure 2 Semi-matrix diagram of χ2-test coefficient (a),association coefficient (b) and Spearman’s rank correlation coefficients of interspecific association among main species
綜上所述,本研究調查區域內的紫莖澤蘭與土蜜樹(Brideliatomentosa)、羊耳菊(Inulacappa)、石海椒(Reinwardtiaindica)及白茅(Imperata cylindrica)之間具有一定的正聯結,而紫莖澤蘭與里白和虎杖的種間聯結均呈極顯著負聯結。
2.3 群落穩定性分析
M.Godron穩定性計算結果見表 4,群落穩定性坐標為(40.25, 59.75),與理想群落穩定點(20,80)相比向下移動了28.63歐氏距離,說明群落處于不穩定狀態。

表4 群落穩定性分析結果(M.Godron法)Table 4 Results of community stability(M.Godron’s method)
2.4 環境因子與物種分布的關系
RDA排序結果顯示,前2個排序軸累積解釋了物種環境關系總方差的78.65%,第1排序軸所含的生態信息豐富,對變異的解釋率較高(76.40%)(表5)。前2軸的特征根之和占總特征根值的78.65%,說明RDA前2軸在解釋物種組成變異中起主要作用。對入侵群落主要物種組成影響顯著的環境因子為有效鉀、土壤溫度、土壤pH值(表6)。其中,有效鉀與第1軸呈極顯著正相關,土壤溫度與第2軸呈顯著正相關,土壤pH值與第2軸呈極顯著負相關(圖3A),這 3個環境因子分別解釋了群落組成 47.00%、19.50%和 13.70%的變異(表 6)。紫莖澤蘭(S6)重要值與全磷、全鉀和土壤溫度均呈較大負相關,而與有效磷、有效鉀和有效氮呈較大正相關。

表5 物種分布與入侵相關指數沿環境因子的RDA排序結果Table 5 RDA calculation of species distribution and invasion-related indexes along environmental gradients
方差分解結果表明(圖4A),土壤因子與地形與生物因子之間有不同程度的協同作用,共同解釋了 13.90%的群落物種組成變異。土壤因子是群落物種組成變異最主要的解釋因子,能夠解釋22.30%的變異。其中,土壤因子對群落物種組成變異的單獨解釋力最大(22.30%);其次為地形與生物因子,解釋 5.60%的變異,另有 58.20%的變異未能被解釋。
2.5 紫莖澤蘭入侵強度、群落可入侵性及群落穩定性與環境因子的關系
RAD前兩軸累計包含了80.69%的解釋率(表5),其中,第1排序軸所含的生態信息豐富,對入侵強度指數、群落可入侵性指數、群落穩定性指數的變異的解釋率較高(79.18%)。決定RDA第1軸的環境因子為總磷(r= -0.911,P=0.002),解釋率為65.7%(表6),沿著RDA第1軸從左往右,全磷含量逐漸下降。入侵強度指數、群落可入侵性指數與全磷呈顯著負相關關系。群落穩定性指數則與總磷呈一定的正相關,而與有效鉀呈一定的負相關(圖3B)。
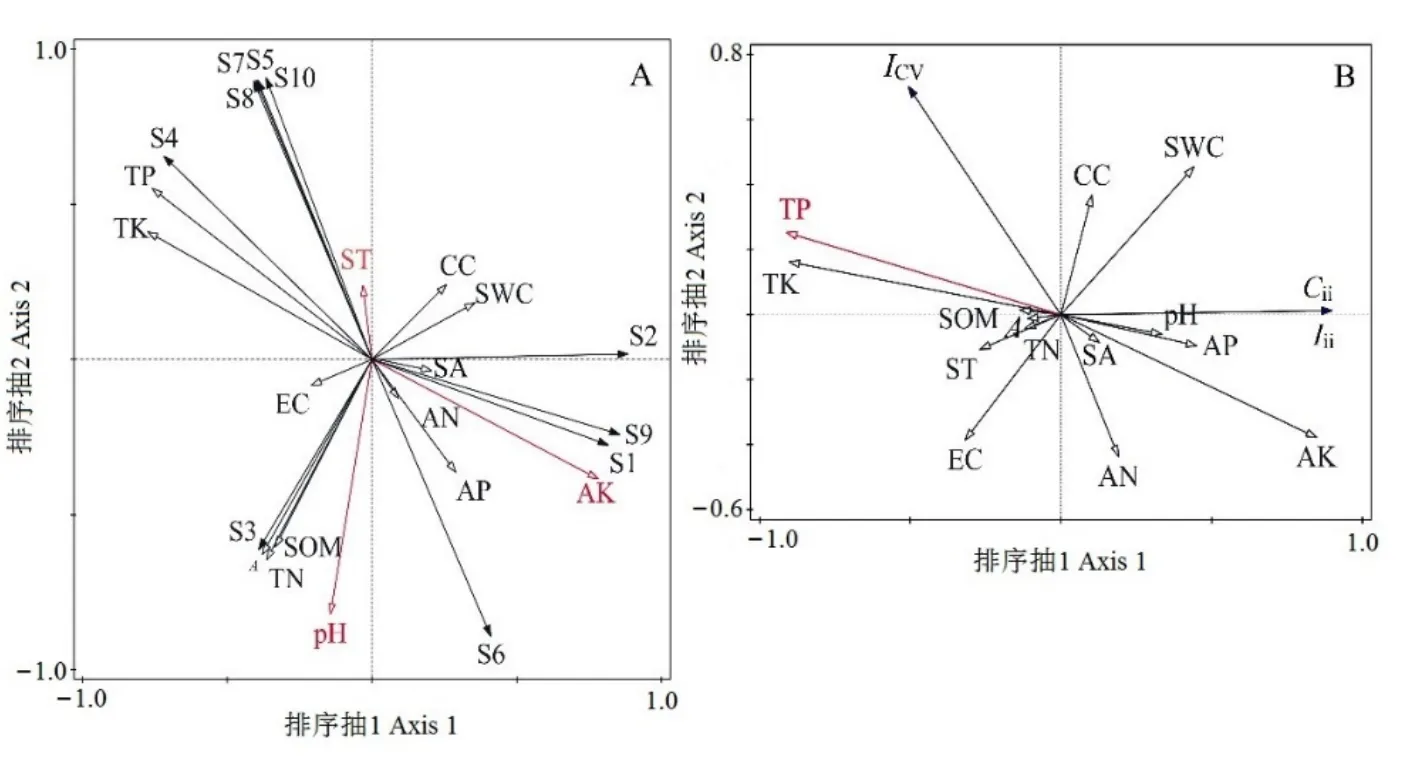
圖3 物種分布(A)和入侵相關指數(B)沿環境因子的RDA二維排序圖Figure 3 Two-dimensional RDA ordination diagram of species (A) and invasion-related indexes (B) along environmental gradients
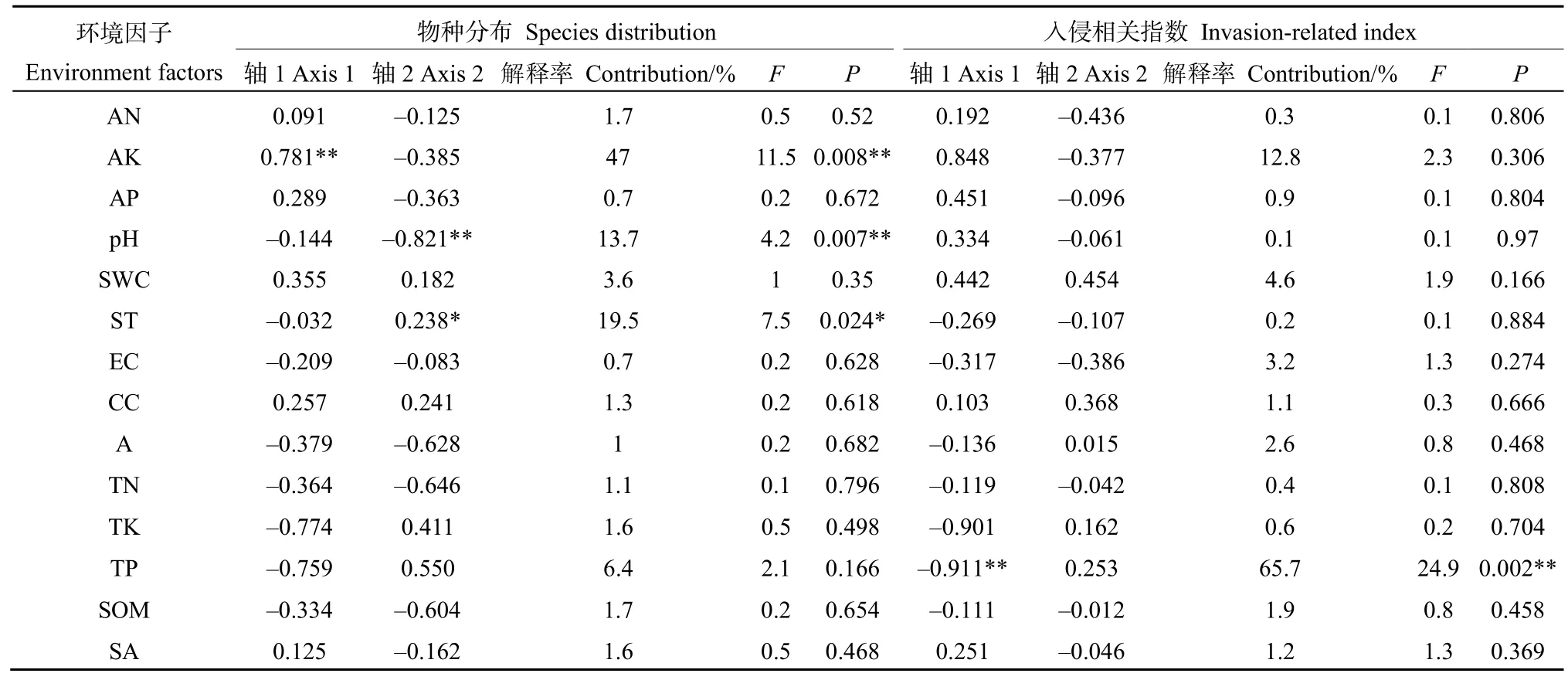
表6 環境因子與RDA排序軸之間的相關系數Table 6 Correlation coefficients between RDA axis and environment factors
方差分解分析結果表明(圖4B),土壤因子和地形與生物因子對入侵相關指數變異的單獨解釋率分別為17.00%和8.90%,兩者交互作用解釋率為4.10%。另有 70%的變異未得到解釋,表明存在較多未知因素影響入侵相關指數。
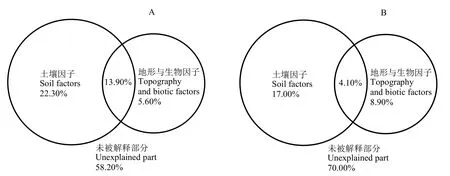
圖4 環境因子對物種分布(A)和入侵相關指數(B)的解釋率Figure 4 Explanatory power of different environmental factors for species distribution (A)and invasion-related indexes (B)
逐步回歸模型分析結果顯示(表 7),全磷與入侵植物入侵強度指數呈顯著負相關,決定系數r2值表明全磷含量能解釋入侵植物入侵強度指數63.90%的變化。全磷與群落可入侵性指數呈顯著負相關,全磷對群落可入侵性指數的解釋率為66.50%。有效鉀與群落穩定性指數呈顯著負相關,對群落穩定性指數的解釋率為44.20%。

表7 入侵相關指數與環境因子的逐步回歸分析Table 7 Stepwise regression analysis of intrusion-related indexes and environmental factors
隨機森林模型結果表明,在所有變量中,全磷對入侵強度指數(圖5A)和群落可入侵性指數(圖5B)的相對重要性最高,而有效鉀對群落穩定性指數的相對重要性最高(圖5C),這些結果與回歸模型所得結果一致。
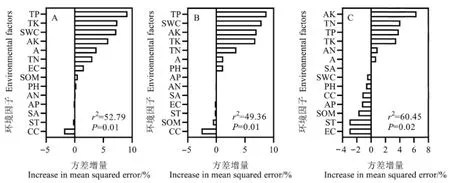
圖5 環境因子對入侵植物入侵強度指數(A)、群落可入侵性指數(B)、群落穩定性指數(C)的重要性Figure 5 Importance of environmental factors to invasion intensity index of alien invasive plants(A), community invasibility index (B) and community stability index (C), respectively
2.6 結構方程分析
結構方程結果表明,影響入侵植物入侵強度指數、群落可入侵性指數及群落穩定性指數的土壤因子相同,但相對作用大小存在差異。對入侵植物入侵強度指數有顯著影響的土壤因子是土壤全磷(P<0.05),其直接路徑系數為-0.833(圖 6A)。土壤全磷對群落可入侵性指數的影響顯著(P<0.05),其直接路徑系數為-0.836(圖 6B)。土壤有效鉀對群落穩定性指數存在直接效應,路徑系數為-0.669(圖6C)。此外,土壤全磷對有效鉀有顯著影響(P<0.01),路徑系數為-0.858(圖6)。

圖6 入侵植物入侵強度指數(A)、群落可入侵性指數(B)、群落穩定性指數(C)與環境因子結構方程模型Figure 6 Structural equation model for invasion intensity index of alien invasive plants (A), community invasibility index (B), community stability index (C) and environmental factors
3 討論
3.1 紫莖澤蘭群落主要物種生態位和種間聯結
在本研究中,主要物種的生態位寬度與重要值排序基本一致。紫莖澤蘭占據最大的重要值和生態位寬度。另外,紫莖澤蘭重要值的變異系數最小(10.50%),說明其在不同樣地中的分布相對均勻。在調查區域內,紫莖澤蘭在群落中占主導地位,具有較強的競爭力以及適應環境、利用資源的能力,這也是紫莖澤蘭快速穩定占領新生境、入侵成功的重要原因(鄧丹丹等,2015;李霞霞等,2017)。紫莖澤蘭群落總體生態位重疊程度不高,而生態位相似性分析結果表明大部分物種對環境資源的利用差異較大,因此,當前群落內大部分物種間的競爭尚不激烈(鄭曉陽等,2018;黃慶陽等,2020),在環境資源充裕時尚能共存。伴生種中里白和虎杖的重要值較大,同時二者與紫莖澤蘭生態位重疊指數均大于 0.5。有研究表明,里白對南方紅壤環境具有獨特的適應機制,且該物種與芒萁一樣都屬于南方紅壤侵蝕區優良的水土保持植物(熊平生等,2012;李小飛等,2013);虎杖為中國傳統中藥材,具有廣泛的藥用價值(Fabian et al.,2021),因其具有較強的生長繁殖能力,虎杖在歐洲多國被認定為入侵植物(Stefanowicz et al.,2017)。本研究中,里白和虎杖在紫莖澤蘭群落中保持較強的競爭力,這與二者具有較強的生長和繁殖能力及較強的生境適應性有關。
通常,處于演替初期的群落,物種間的關聯性較弱,甚至表現出較強的負關聯,而正聯結的種對越多,物種間關聯性越強,群落越穩定(郭忠玲等,2004;劉潤紅等,2020)。本研究中,主要物種間的總體聯結性表現為顯著負聯結,且χ2檢驗、聯結系數AC測定以及Spearman秩相關系數檢驗結果均表明紫莖澤蘭群落中負相關的種對數遠大于正相關的種對數,表明群落尚未達到穩定。種間正聯結有2種情況:(1)一個物種對另一物種產生依賴;(2)生態學特征和生存環境的差異和互補性,使各物種相互兼容。本研究中,紫莖澤蘭與土蜜樹、羊耳菊、石海椒等優勢灌木之間呈現出一定的正聯結。另外,里白和虎杖之間呈現出一定的正聯結,二者與草本層的白蒿(Herbaartimisiae)、芒萁及野豌豆(Viciasepium)等本土草本之間亦呈現一定的正聯結。推測本研究中大部分種對間的正聯結關系屬于第二種情況。物種之間存在空間或資源的排斥爭奪造成種間負關聯(郭佳琦等,2021),本研究中里白和虎杖與紫莖澤蘭的種間聯結均呈極顯著負聯結,二者與紫莖澤蘭之間存在著較大的排斥性,表明二者與紫莖澤蘭之間存在空間、環境資源等的競爭。通常認為,種間聯結度與生態位重疊指數呈正相關(吳佳夢等,2019),即種間正聯結越強,生態位重疊越大,反之亦然。亦有研究認為,種間關聯程度與生態位重疊值并不具有一致性(吳佳夢等,2019;姜倪皓等,2021),由資源競爭引起的負關聯生態位重疊值較高(陳旭,2008),本研究中亦得到類似結果,這也表明從生態位理論來解釋入侵植物與本土植物優勢種種間關聯性存在一定的局限性。
競爭替代控制是入侵植物主要的生態控制方法,亦是目前普遍認可的控制方法(Li et al.,2015)。在兼顧生態效益和經濟效益的前提下,綜合群落可入侵性-物種功能群多樣性假說(Naeem et al.,2000)、替代控制植物選擇標準(Li et al.,2015)及本文結果,本研究建議將具有較強生長能力、適應性較強、兼具生態或經濟效益作為本研究區域選擇替代控制植物的重要標準。未來或可考慮在研究區域內利用本土植物里白和虎杖對哀牢山地區紫莖澤蘭進行替代控制。在紫莖澤蘭已入侵的群落中優先栽培里白和虎杖,既可以抵抗紫莖澤蘭的入侵,又能在資源匱乏時與其形成利用性競爭,遏制紫莖澤蘭的蔓延。同時,里白還能發揮水土保持功能,而虎杖作為一種藥用價值很高的中藥藥材,或可成為林區經濟發展新的增長點。此外,盡管虎杖作為在當地自然生態系統中自然定居的本土植物,亦有研究表明在歐洲多地河岸區,虎杖的落葉分解產生了植物毒素,進而對河岸和溪流不同植物群落產生正向和負向雙方面的影響(Cybill et al.,2020),因此,亟待開展相關的生態風險監測體系及替代控制植物配置模型的應用研究。
3.2 環境因子對物種分布及紫莖澤蘭入侵的影響
入侵植物可改變入侵地的生態環境,從而利于自身生長或抑制其他植物生長(Holmgrem et al.,1997;萬方浩等,2011)。土壤養分在紫莖澤蘭植物入侵過程中起著重要作用,研究表明紫莖澤蘭入侵群落的土壤有機質、全氮、有效磷、有效鉀含量顯著提高,紫莖澤蘭入侵顯著提高了土壤養分,且有效養分對紫莖澤蘭入侵較敏感(Hu et al.,2019;劉潮等,2007;吳天馬等,2007;鄧丹丹等,2015)。另外,亦有研究發現,紫莖澤蘭入侵群落的土壤全磷、有效磷、有效鉀含量均顯著下降(Sun et al.,2013;吳天馬等,2007)。本研究中,紫莖澤蘭重要值與全磷和全鉀均呈較強的負相關,而與有效鉀呈較強的正相關。紫莖澤蘭可直接利用的速效元素的含量增加,對紫莖澤蘭生長發育有促進作用,紫莖澤蘭這種改變土壤營養環境的能力為其在貧瘠生境的順利入侵提供了有利條件(Hu et al.,2019;劉潮等,2007;鄧丹丹等,2015)。另一方面,本研究結果亦表明,在多數情況下,紫莖澤蘭入侵對土壤養分的影響較復雜,沒有表現出一致的規律性。
本研究中,對紫莖澤蘭群落主要物種組成影響顯著的環境因子為有效鉀、土壤溫度、土壤pH值。強大入侵力源于入侵植物的強大的養分競爭特性,而這將導致土壤養分被大量消耗,從而影響本地植物的生長(Stefanowicz et al.,2017;鄧丹丹等,2015)。有研究表明,紫莖澤蘭植株的鉀含量較高(劉潮等,2007),同時紫莖澤蘭在生長過程中對土壤中的含磷和含鉀的化合物具有很強的活化作用,并且對磷、鉀具有較強的吸收能力和較大的需求量(Hu et al.,2019;牛紅榜等,2007)。本研究發現,紫莖澤蘭的入侵強度指數和群落可入侵性指數均與有效鉀呈較強的正相關。因此,推測紫莖澤蘭將土壤中大量的全鉀轉換成有效鉀,然后被其吸收利用,促使其在與本地植物群落的競爭中取勝,從而利于其成功入侵(Hu et al.,2019;鄧丹丹等,2015)。本研究中,逐步回歸分析、隨機森林模型及結構方程模型的結果均表明,群落穩定性指數與土壤有效鉀含量呈極顯著負相關,紫莖澤蘭入侵地的物種多樣性隨入侵程度加重而降低(張修玉等,2010),以致植物功能群單一化,進而導致群落穩定性下降。
南方土壤酸性強,活性鐵、鋁含量高,土壤對磷固定作用強,有效磷含量低(Yan et al.,2020;李杰等,2011)。本研究樣地土壤全磷含量均值為1.708 g·kg-1,遠高于中國表層土壤全磷含量均值0.78 g·kg-1(Tian et al.,2010),但是可能由于樣地所處區域屬于酸性紅壤地區(李杰等,2011),土壤的固磷作用較強,導致土壤磷有效性降低,磷難以被植物利用而成為植物生長的主要限制因子。本研究中,全磷與紫莖澤蘭入侵強度指數和群落可入侵性指數均呈極顯著負相關關系。此外,本研究發現,土壤全磷與有效鉀呈較強的負相關,有研究表明在耕作土中土壤有效鉀含量隨著磷施用量的增加而降低(鄭昭佩等,2004),但是在自然土壤條件下,尚無法對此作出很好的解釋。基于本研究結果,推測紫莖澤蘭入侵促進土壤全鉀向植物可利用的鉀素形態轉化,而有效鉀含量提高則促進紫莖澤蘭生長,導致土壤pH提高(劉潮等,2007),進而提高磷素的有效性(李杰等,2011),最終導致土壤全磷含量降低。
本研究中,紫莖澤蘭重要值與土壤pH呈較強的正相關,而紫莖澤蘭的入侵強度指數和群落可入侵性指數均與土壤pH呈一定的正相關。研究表明,紫莖澤蘭凋落物分解產生的酸性物質較少,其群落下土壤pH較高,這有利于土壤酶發揮作用,利于其對養分的吸收(劉潮等,2007)。土壤過度酸化,將會使土壤理化性質變劣,不利于土壤養分的保存和積累(Mccoll et al.,1991)。本研究區主要的喬木樹種為云南松(Pinusyunnanensis),有研究表明云南松通過根際酸化來適應低磷環境(戴開結等,2005)。另外,研究表明,虎杖群落能顯著降低土壤 pH 值(Dassonville et al.,2011;Stefanowicz et al.,2017),在本研究中亦觀察到了類似的結果,而土壤pH值降低將進一步降低磷等元素的有效性(文亦芾等,2005),進而可能限制紫莖澤蘭的蔓延。
本研究樣地均處于海拔1600 m以上的高山地區,氣溫低,晝夜溫差大,而紫莖澤蘭重要值與土壤溫度呈一定的負相關。研究表明,紫莖澤蘭種子萌發溫度范圍較廣,在亞適宜的低溫下,萌發率和萌發指數均高于本地植物(張麗坤等,2014)。寒冷冬季,在本地植物葉片凋落、處于休眠和半休眠狀態時,紫莖澤蘭仍保持大面積的葉片與活躍的光合作用(Wang et al.,2013),另外紫莖澤蘭表現較強的耐低溫逆境能力(Gong et al.,2010),這些生物學特性將促進紫莖澤蘭利用其他物種不能利用的資源,進而有利于其向更高海拔區域擴展。
植被郁閉度在70%以上,紫莖澤蘭幾乎無法入侵(何永福等,2005)。本研究中,紫莖澤蘭重要值和郁閉度幾乎無相關關系,本研究樣地主要喬木為云南松,且郁閉度均值為73%,針葉林特殊的葉形使得針葉林林下光照強度仍顯著高于闊葉林,因此光照未成為紫莖澤蘭入侵的主要限制因子(朱文達等,2013;宋紫玲等,2019)。
4 結論
(1)紫莖澤蘭入侵群落中,紫莖澤蘭的重要值和生態位寬度均最大。伴生種中里白和虎杖的重要值較大,同時二者與紫莖澤蘭的生態位重疊指數均大于0.5。
(2)入侵群落總體呈現顯著負關聯,群落處于非穩定狀態。紫莖澤蘭與里白和虎杖均具有極顯著的負聯結關系。未來或可考慮利用里白和虎杖對哀牢山地區的紫莖澤蘭進行替代控制。
(3)對紫莖澤蘭入侵群落主要物種組成影響顯著的環境因子為有效鉀、土壤溫度、土壤pH值。
(4)土壤全磷與入侵植物入侵強度指數及群落可入侵性指數呈顯著負相關,有效鉀與群落穩定性指數呈顯著負相關。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
