基于產量相關性狀SSR分子標記的大豆雜種優勢群劃分
2022-09-13 07:41:28關哲允曹士亮王玉民林春晶彭趙麗梅李志剛張春寶
作物雜志 2022年4期
關鍵詞:大豆
雷 蕾 關哲允 曹士亮 王玉民 林春晶彭 寶 劉 鵬 趙麗梅 李志剛 張春寶
(1內蒙古民族大學農學院,028042,內蒙古通遼;2吉林省農業科學院大豆研究所/農業農村部雜交大豆育種重點實驗室,130033,吉林長春;3黑龍江省農業科學院玉米研究所,150086,黑龍江哈爾濱)
大豆作為我國重要的糧食、油料以及飼料兼用作物,近年來的生產總量無法滿足日益攀升的消費需求。目前我國是世界上最大的大豆消費國,也是全球最大的進口國,供需關系失衡,僅靠國內生產難以滿足需求,大豆高度依賴進口的局面難以改變[1]。這其中的主要原因是我國大豆單產低,效益比較差,同時耕地面積難以增加,因此提高大豆的單產水平迫在眉睫。雜種優勢普遍存在于生物界,在許多動植物上均得以體現,并且被認為是提高作物產量的最有效途徑之一[2]。雜種優勢在水稻、玉米、高粱、油菜和棉花等農作物的生產上廣泛應用并獲得巨大成功,世界農業生產得到了極大發展[3-5]。雜交大豆同樣具有較強的雜種優勢,隨著世界上第1個大豆雜交種“雜交豆1號”育成并通過審定[6],大豆雜種優勢利用進入新階段。目前我國在該領域一直處于國際領先水平,截至2021年底,在東北及黃淮海大豆主產區已審定39個大豆雜交品種[1]。雖然目前已有一批大豆雜交種育成并通過審定,但是在雜交種實際選育過程中仍存在大量雜交組合盲目配制的問題,導致雜交種的產出率低,部分雜交組合的雜種優勢不強,限制了強優勢雜交種的產業化應用。借鑒玉米和水稻等作物的雜交育種經驗,只有將來源不明的種質資源劃分到正確的雜種優勢群,才能更好地利用雜種優勢模式,提高育種效率,培育出更多的新品種,將雜種優勢最大效應[7]發揮在雜種優勢群內,進行親本遺傳改良和創制,在不同雜種優勢群間進行雜交組合配制,是強優勢雜交種選育的重要理論基礎。
簡單序列重復(simple sequence repeats,SSR)作為一種多態性高、重復性好且較穩定的分子標記,在雜種優勢群劃分和遺傳多樣性分析中被廣泛應用。如Senior等[8]利用70個SSR標記對94份美國玉米自交系材料進行雜種優勢類群的鑒定分析,證實雜種優勢類群的鑒定結果與系譜吻合;聶永心等[9]利用70個SSR分子標記對20個玉米自交系劃分了5個雜種優勢群,聚類分析結果與系譜基本一致,表明SSR標記可以進行玉米自交系遺傳變異分析,并用于雜種優勢群的劃分;羅小金等[10]利用36個SSR分子標記將58份秈型水稻劃分為3個雜種優勢類群,并發現類群間組合優勢強于類群內組合;張濤等[11]利用44個與水稻產量性狀基因緊密連鎖的功能基因標記對三系雜交水稻親本進行遺傳多樣分析,將雜交水稻親本劃分為保持系群和恢復系群,并且證明了功能基因標記與普通分子標記相比具有更高的DNA多態性檢測效率,能夠準確、可靠地劃分類群;于海至[12]利用49個SSR分子標記對199份油菜強優勢雜交種親本進行聚類分析,最終將大部分親本劃分為2個雜種優勢群,分析后發現遺傳距離中等偏大的親本間進行組配,雜種優勢較為明顯。在大豆中也開展了利用SSR標記進行雜交大豆親本間遺傳多樣性的分析。吳倩等[3]利用SSR分子標記遺傳距離預測大豆親本間雜種優勢,最終發現SSR分子標記對大豆親本間雜種優勢預測未能達到顯著水平;白志元等[13]也利用了SSR標記對32份雜交大豆親本進行了遺傳多樣性分析,經聚類分析發現,親本間可以根據地理來源進行類群劃分。但上述研究僅僅是通過多態性標記進行分群聚類,不同群內和群間雜交是否具有雜種優勢的研究尚未深入展開。
本研究利用篩選獲得的14個在雜交大豆親本間具有較好多態性的產量性狀連鎖SSR標記,對2002-2022年在東北春大豆主產區審定的30個雜交種所用的43份親本材料進行親緣關系分析和遺傳多樣性評價,探討親本間地理來源及遺傳距離對強優勢雜交種創制的影響,為今后指導親本選育方向及提高雜交組合組配效率提供理論依據。
1 材料與方法
1.1 試驗地概況與材料
試驗材料為吉林省農業科學院雜交大豆團隊育成的30個春大豆雜交種的親本材料,包括來源于中國東北、美國和意大利的不育系配套保持系20份與恢復系23份,共計43份。所有試驗材料于2021年5月種植在吉林省長春市范家屯鎮雜交大豆核心育種基地(124°83′E,43°43′N)。試驗地行長 3m,行距65cm,株距15cm。對所有試驗材料進行編號,每份試驗材料種植1行。進行常規田間管理,并在試驗田四周設置保護行,在行頭插牌做標記。選取43份親本材料V2期(第2個3小葉時期)的新鮮葉片置于液氮中運輸,于-80℃冰箱保存。
1.2 SSR標記篩選
對公開發表文獻中與大豆籽粒大小、單株粒重、每莢粒數、單株莢數、百粒重、分枝數和主莖節數等產量性狀相關SSR分子標記進行檢索,選取66個緊密連鎖的SSR標記,其引物序列送北京六合華大基因科技有限公司合成。篩選多態性好、重復性高、帶型清晰的分子標記用于對43份親本材料進行多態性的檢測。
1.3 基因組DNA提取及PCR檢測
使用Nu Clean Plant Genomic DNA Kit(北京康為世紀生物科技有限公司)提取試驗材料葉片基因組DNA,并用1%瓊脂糖凝膠電泳以及微量分光光度計(thermo,NanoDrop 2000)對提取的基因組DNA進行濃度和質量檢測,檢測合格的DNA放置于-20℃冰箱保存。
利用SSR分子標記對親本DNA進行PCR擴增。PCR反應體系為15μL,包括2×ES Taq Master Mix For Page(北京康為世紀生物科技有限公司)7.5μL、ddH2O 4.5μL、上下游引物各 1μL、模板 1μL。PCR反應程序為95℃預變性5min;95℃變性30s,退火溫度為42℃~60℃,72℃延伸30s,循環33次;72℃延伸5min,最終4℃保存。利用8%聚丙烯酰胺凝膠電泳進行檢測,電泳前在PCR產物中加入變性劑3μL[14],95℃變性10min。變性后的樣品置于凝膠點樣孔恒定電壓300V電泳2h,電泳結束后在400mL水中加入Super GelRed熒光染料(蘇州宇恒生物科技有限公司)試劑40μL,染色10~20min,通過紫外凝膠成像儀(UVP,GelDoc-It 310)成像,記錄條帶數據。
1.4 數據處理
根據電泳條帶遷移的位置對試驗結果進行記錄,同一位置條帶以同一數字進行記錄,缺失條帶記為“9”,利用Excel記錄試驗結果。利用Power Marker v3.25軟件,采用算術平均非加權方法(unweighted pair-group method with arithmetic means,UPGMA)對親本材料進行聚類分析,獲得聚類樹狀圖。利用Power Marker v3.25[15]計算每個標記的多態性信息含量(polymorphism information content,PIC),PIC=1-∑Pi2,Pi表示群體中含有第i個等位變異的比例[16]、等位變異數(allele number,Na)、遺傳多樣性系數(genetic diversity index,GI)、主效基因頻率(major alle frequency,MAF)和遺傳距離(genetic distance,GD)。根據SSR標記對43份親本材料檢測到的基因型及利用Power Marker v3.25軟件得到的分群結果,利用R語言ggplot 2作主坐標分析(principal co-ordinates analysis,PcoA),最終將各點PcoA結果坐標繪制在二維平面圖上,制成散點圖。
2 結果與分析
2.1 多態性SSR分子標記篩選
通過對文獻檢索并合成的66個與產量相關性狀緊密連鎖的SSR分子標記,首先利用4份不同來源地的親本進行初篩,最終篩選出14個多態性好、帶型清晰、檢測結果穩定的SSR標記用于進一步分析,具體分子標記信息見表1。

表1 14個SSR標記信息Table 1 14 Primers information table
2.2 基于SSR多態性數據的聚類分析
根據PowerMarker v3.25軟件通過Nei's距離利用UPGMA將43份材料的SSR標記多態性數據進行聚類分析。根據聚類結果(圖1),43份親本被聚為2個類群。其中類群Ⅰ中包含27份材料,大部分為中國來源,國外材料有美國JLCMS179B、JLCMS254B和JLCMS164B與意大利JLR83;類群Ⅱ中包含16份材料,均為美國來源。從表2可以看出,2002-2022年東北春大豆區審定的30個雜交種中,25個雜交種的親本屬于2個類群,另有4個為類群Ⅰ內雜交獲得,1個為類群Ⅱ內雜交獲得。說明多數的雜交種親本來源分屬于不同的雜種優勢類群。
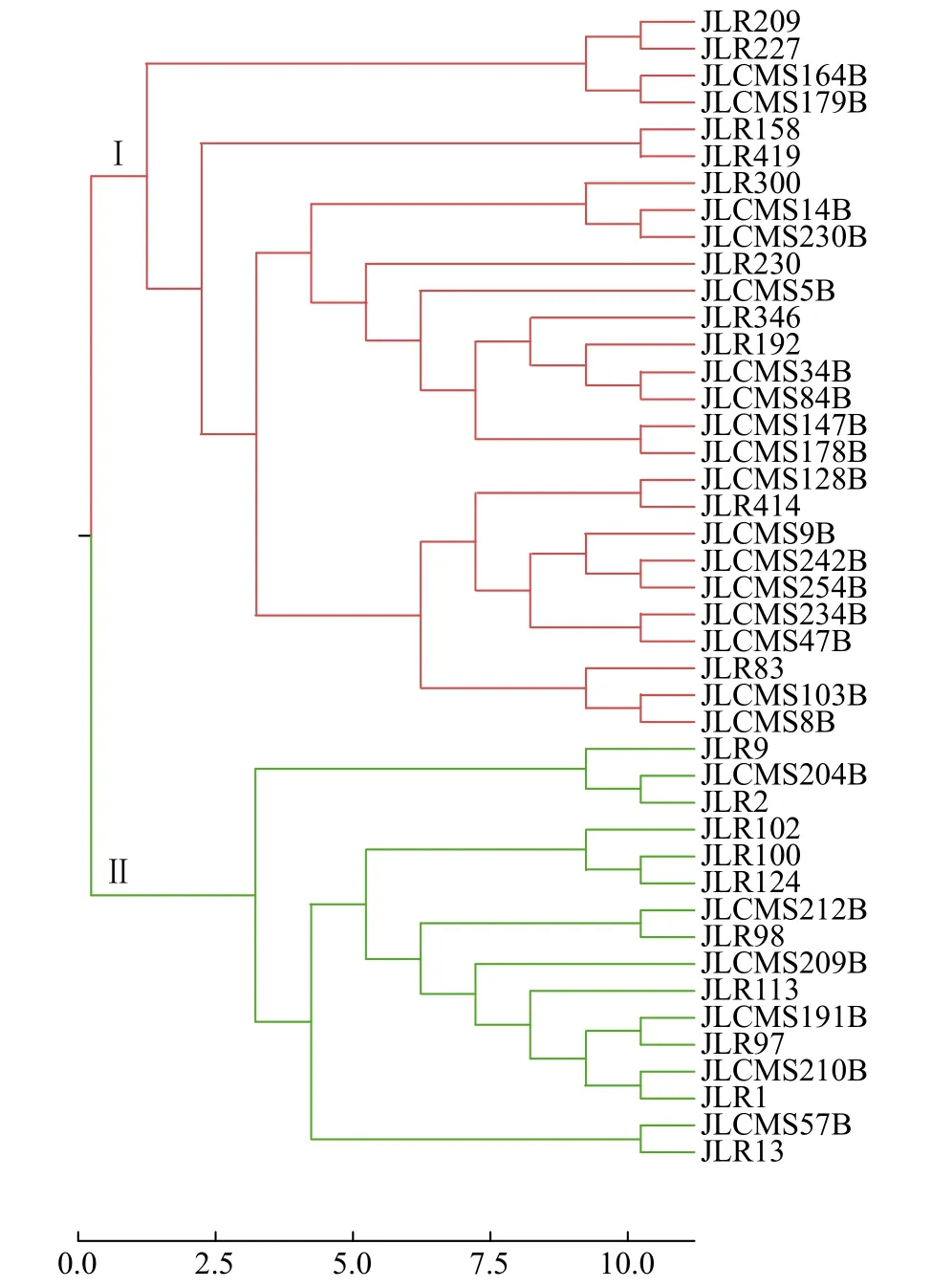
圖1 SSR標記對親本材料的UPGMA聚類分析Fig.1 UPGMA cluster of parent materials based on SSR markers
另外,根據表2中所列親本間的地理來源信息,26個雜交種屬于國內與國外親本雜交獲得,僅有2個為美國親本間雜交獲得,2個為國內親本間雜交獲得。雜交種親本來源既滿足不同類群又符合不同地理來源的雜交種個數為23個,說明分子標記劃分的雜種優勢類群結果與地理來源分布基本一致。分子標記劃分的雜種優勢類群結果與地理來源分布不一致的情況主要表現為,雜交種親本均來自美國,但分子標記分析結果可以劃分為類群Ⅱ(吉育612),分別屬于美國和中國的吉農H1和吉育667以及均屬于中國的吉育635和吉育653被劃分到了類群Ⅰ。
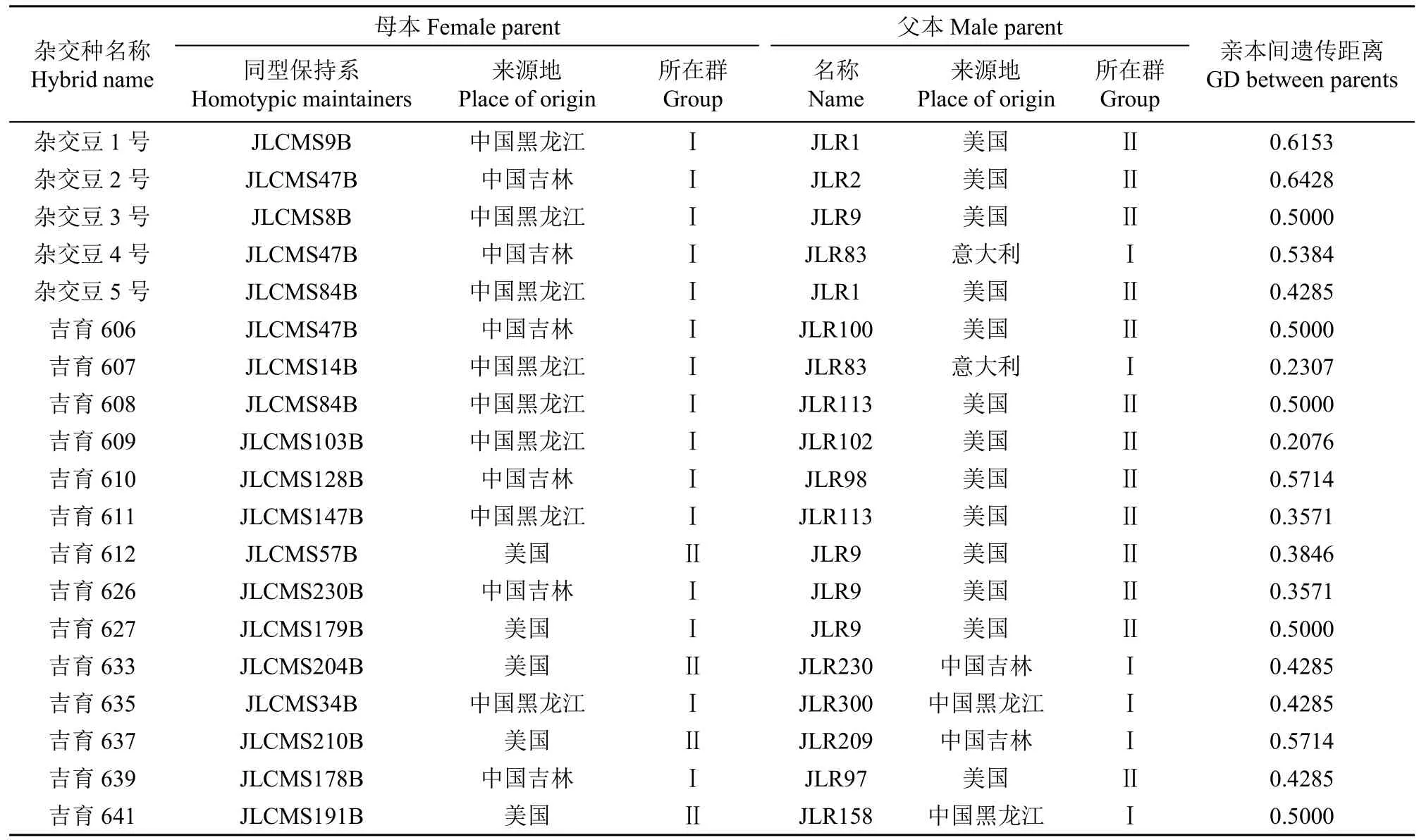
表2 雜交種親本分群結果Table 2 Results of parental strata of hybrid species
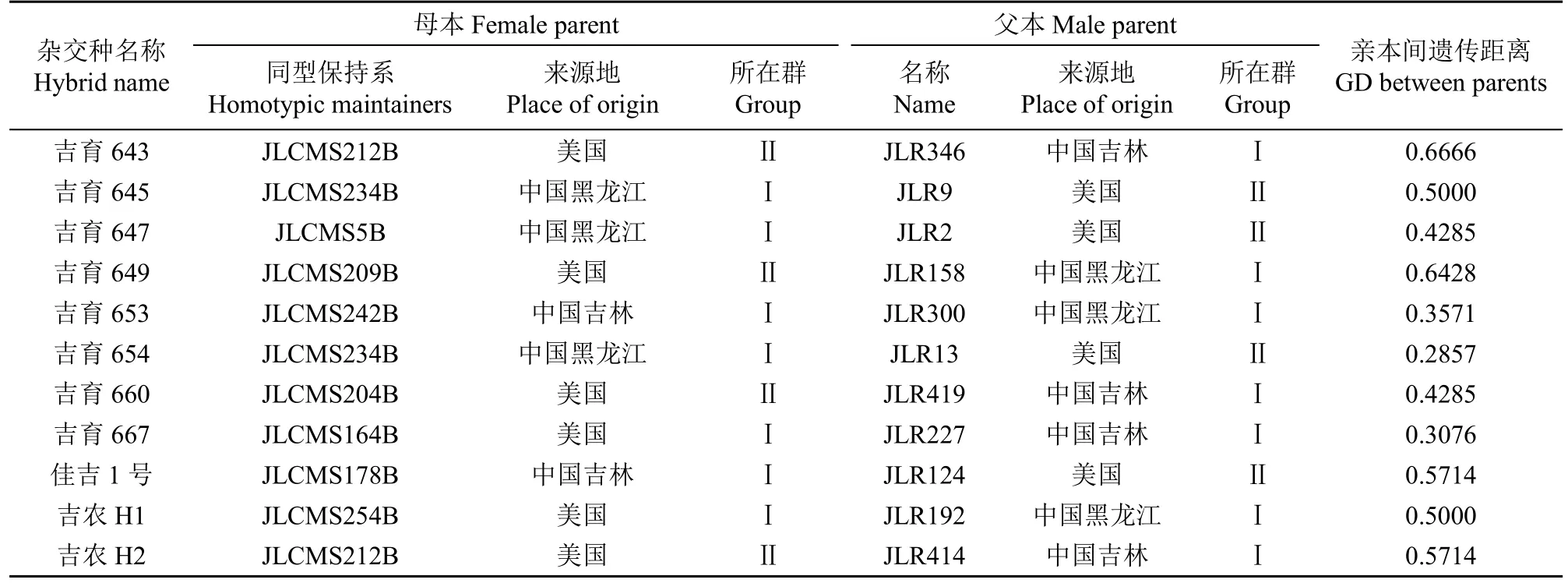
續表2 Table 2(continued)
進一步對雜交種親本間GD的分布情況(圖2)進行分析發現,43份親本間的GD均值為0.3845,30個雜交種成對親本間的GD均值為0.4650,親本間GD在0.5~0.6的組合頻率最高,共有12個,占所有組合的40.00%。親本間GD在0.4~0.5的組合有6個,占總數的20.00%,親本間GD在0.3~0.4的組合有5個,占總數的16.67%。推測GD在0.4~0.6時雜種優勢利用效率較高。考慮到大豆雜種優勢利用時,為減少組配的盲目性,優先組配親本GD在0.5~0.6的雜交組合。
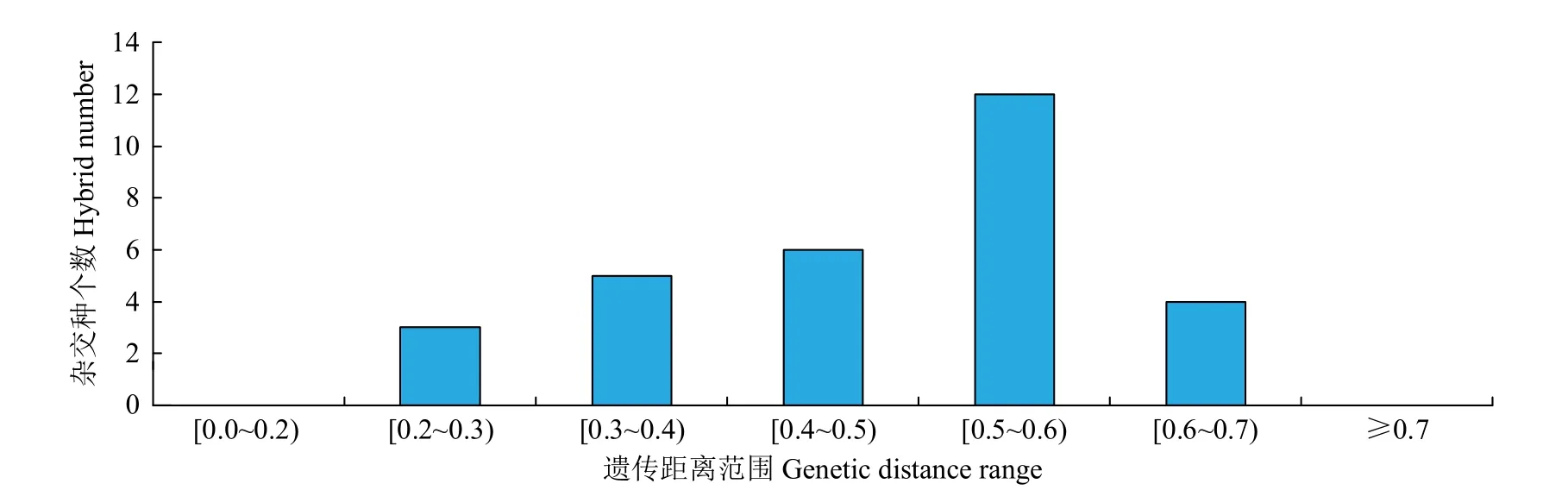
圖2 SSR標記分析雜交種親本遺傳距離分布Fig.2 Genetic distance distribution of hybrid parents based on SSR markers
2.3 遺傳多樣性分析
進一步對43份親本材料進行遺傳多樣性分析(表3和圖3)發現,總群的MAF均值為0.6871,范圍在0.4419~0.9302;類群Ⅰ均值為0.7233,范圍在0.4074~1.0000;類群Ⅱ的均值為0.7622,范圍在0.5000~1.0000。總群的Na均值為2.2143,范圍在2.0000~4.0000;類群Ⅰ的均值為2.0714,范圍在1.0000~4.0000;類群Ⅱ的均值為1.9286,范圍在1.0000~2.0000。總群的GI均值為0.4043,范圍在0.1298~0.6101;類群Ⅰ的均值為0.3449,范圍在0.0000~0.6475;類群Ⅱ均值為0.3243,范圍在0.0000~0.5000。總群的PIC均值為0.3280,范圍在0.1214~0.5272;在類群Ⅰ中,PIC均值為0.2801,范圍在0.0000~0.5720;在類群Ⅱ中,PIC均值為0.2612,范圍在0.0000~0.3750。總群GD范圍在0.0000~0.6923,類群Ⅰ的范圍在0.0000~0.6153,類群Ⅱ的范圍在0.0000~0.4285。上述結果表明,類群Ⅰ的遺傳多樣性水平高于類群Ⅱ。
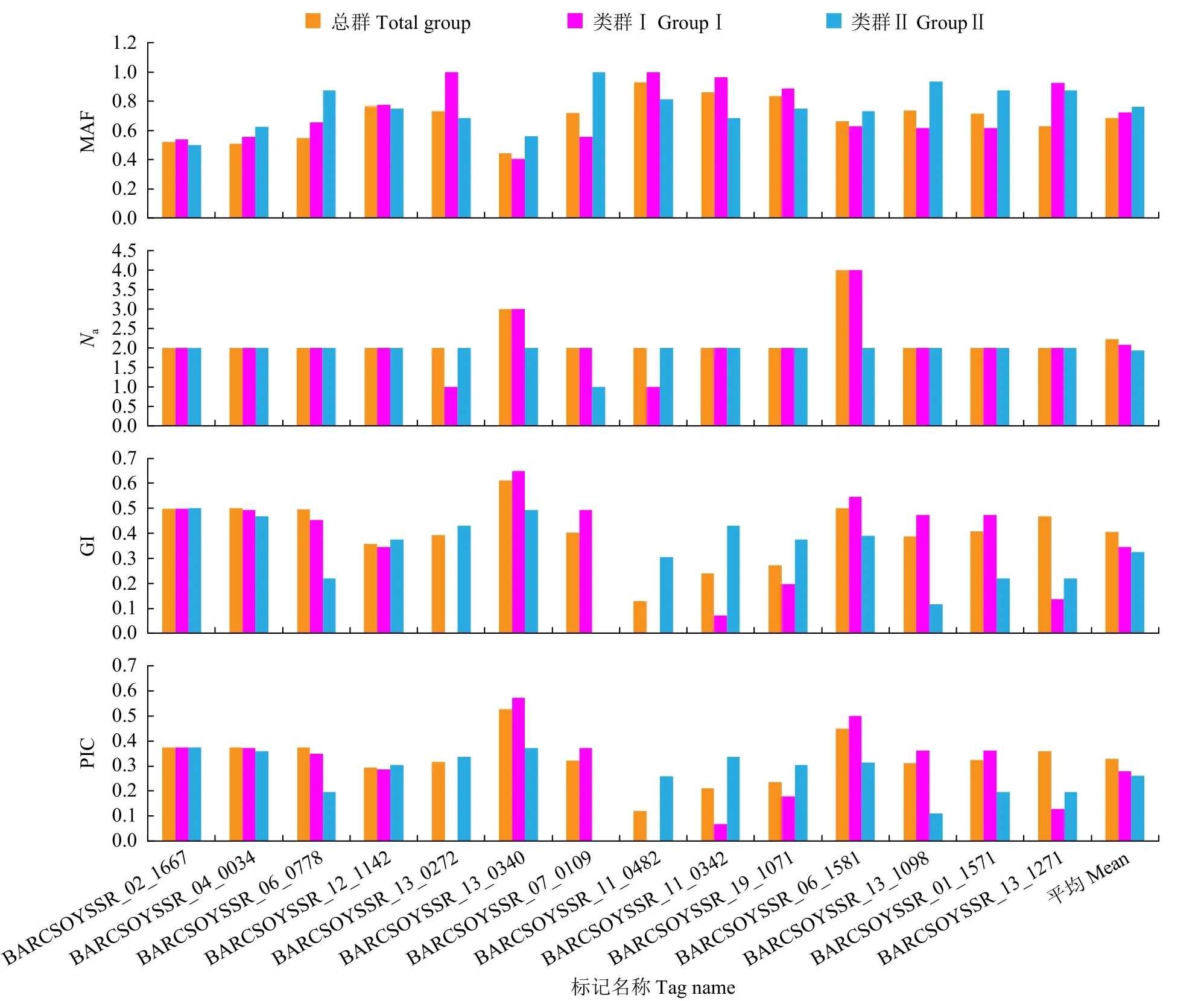
圖3 SSR標記對群體間的遺傳多樣性分析Fig.3 Analysis of genetic diversity between groups by SSR markers
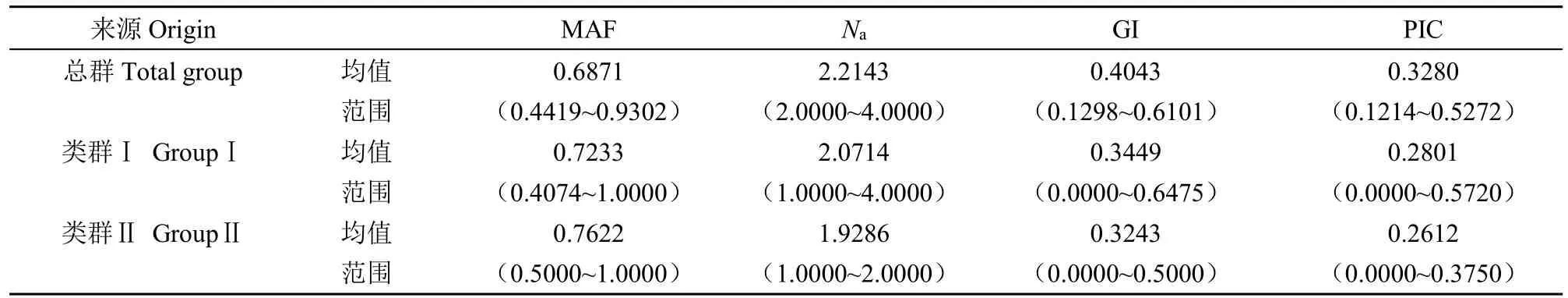
表3 SSR標記對不同群試驗材料的遺傳多樣性分析Table 3 Genetic diversity analysis of test materials by SSR markers on different groups
從圖4中可知,利用PC1和PC2主成分將43份親本清晰地劃分為2個集合,集合Ⅰ包含23份材料,70%以上為中國東北材料;集合Ⅱ包含20份材料,全部為美國材料;有8份材料在2集合交匯位置聚集,這部分材料中存在混合血緣材料。這與SSR標記聚類圖的分析結果較為一致,證明中國東北與國外材料之間配制的雜交組合存在較強的雜種優勢。材料間遺傳距離分布范圍較廣,散點密集度較小,進一步表明中國東北春大豆區審定的雜交種的親本材料分化程度較高,遺傳差異較大,種質資源背景較為豐富。
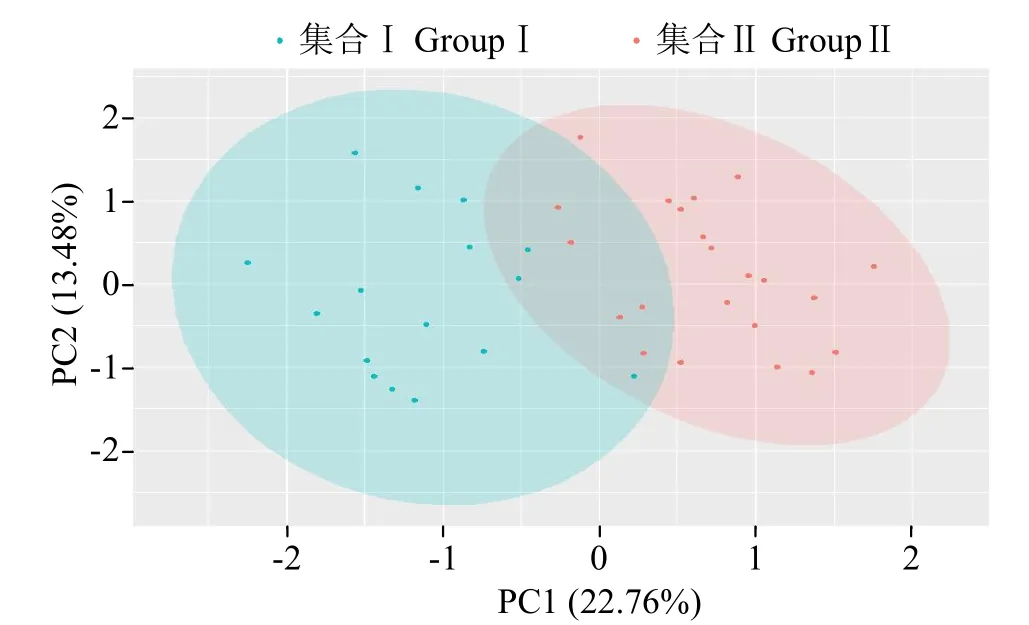
圖4 基于SSR標記檢測基因型及分群結果的PcoA散點圖Fig.4 PcoA scatter plot based on SSR marker detection of genotype and clustering results
3 討論
本研究利用SSR分子標記根據遺傳距離對30個審定大豆雜交種的43份親本材料進行聚類分群,劃分結果與親本地理來源結果基本一致,相同地理來源的試驗材料基本可以聚在一個大群中,這與鐘文娟等[29]和魏苗等[30]的研究結果相同,表明SSR標記聚類結果與地理來源之間存在一致性。個別不同來源材料會聚類到一個類群,可能是由于跨地區引種導致的遺傳背景聚合所致,類似的結果在糜子聚類分群研究中也同樣出現[31];強海平等[32]利用SSR標記對中國和美國來源的紫花苜蓿進行聚類分析時也發現,由于每個品種存在多個親本參與雜交的過程,導致部分品種間存在一定的親緣關系,聚類時個別材料出現偏差。也可能是因為本研究所用SSR標記數量未能覆蓋大豆全部20條染色體,對個別親緣關系較為復雜的親本無法進行精準判斷所致。
從分群結果來看,30個審定春大豆雜交種中有26個為中國東北與國外材料雜交組配育成,且上述雜交種親本大多分屬于不同遺傳類群,也印證了遺傳背景差異越大,雜種優勢越強的理論[33]。另外,也不乏國內或美國材料內部的組配育成了強優勢雜交種,這表明同一來源且屬于同一類群間的親本同樣會育成強優勢雜交種,但概率會較低。根據43份親本聚類分群結果來看,80%的雜交種的親本分屬于不同類群,大部分雜交種均由中國和國外材料進行組配獲得。但個別材料如來源于美國的JLCMS164B和JLCMS254B被聚在中國東北材料群中,推測其可能由中國引進尚未被改良或尚存大部分中國材料遺傳背景,所以通過SSR標記分析將其與中國材料聚入同一類群。
Lippman等[34]認為育種者應該努力增加親本種群之間的遺傳距離,以最大化雜種優勢。但Moll等[35]卻提出,親本不應超過最佳GD,以避免由于有害的上位效應而降低雜種優勢。然而Boeven等[36]最近的研究證實,雜種優勢隨親本間GD的增加而穩定增加,父母本群體之間可以盡可能保持距離,后代優勢不會因此而下降。本研究計算了43份春大豆雜交種親本間的GD,發現所用雜交種親本的GD在0.4~0.6時,雜種優勢利用程度較高,這與Moll等[35]的結論較為一致,然而本研究中仍有部分雜交種親本的GD在0.6~0.7。因此,雜種優勢是否存在最佳GD或者會隨親本間GD的增加而增加,仍有待于進一步開展研究。
4 結論
利用14個大豆產量性狀連鎖SSR分子標記對2002-2022年中國東北春大豆產區審定的30個雜交種的43份親本材料進行雜種優勢群劃分和驗證,通過UPGMA法將43份親本材料劃分為2個類群,不同地理來源種質資源親緣關系較近,聚類結果與地理來源基本一致。已審定強優勢雜交種的親本80%以上分屬上述2個類群,且為中國東北與國外材料雜交獲得,表明GD較遠且存在地理遠源的雜交大豆親本材料之間進行雜交組配,會產生較強的雜種優勢。
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
