甜蕎莖稈重心高度和抗折力的遺傳分析
2022-09-13 07:41:48胡丹
作物雜志 2022年4期
胡丹
(甘肅省種子總站,730030,甘肅蘭州)
甜蕎(Fagopyrum esculentum Moench)起源于中國,因其生育期短、適應性廣和抗逆性強等特點,在作物的生產布局中具有特殊的地位,是主要的填閑復播和備荒救災作物[1]。我國是甜蕎的生產大國,種植面積和產量居世界第2位。甜蕎營養物質豐富,被營養學家稱為21世紀最有前途的綠色食品[2-3],甜蕎含有較高的類黃酮、蛋白質、維生素、脂肪酸、礦物質和粗纖維等物質[4],具有降“三高”、防癌抗癌、抗過敏、利尿和鎮咳等功效[5-8]。莖稈易倒伏和彎曲是當前甜蕎生產中存在的主要問題,輕者減產,重者絕收,極大地制約了甜蕎增產[9-10]。因此研究甜蕎抗倒伏性狀的遺傳規律對實現其高產、穩產具有重要意義。
莖稈重心高度和抗折力是影響作物倒伏的重要指標,許多研究[11-16]表明,在一定范圍內,莖稈重心高度較低的甜蕎品種,其莖稈抗折力參數大,倒伏指數小,抗倒伏能力強;邵慶勤[17]和楊文飛等[18]研究表明,小麥重心高度和機械強度與其倒伏密切相關。袁圓等[19]研究表明,油菜莖稈抗折力是影響其倒伏的重要因素。從本質上明確品種特征特性的遺傳規律,才能指導育種實踐。蓋鈞鎰等[20]提出的植物數量性狀主基因+多基因混合遺傳模型分離分析方法已廣泛應用于多種作物多種性狀的遺傳分析。目前關于甜蕎莖稈重心高度和抗折力遺傳分析的研究鮮見報道。本文利用主基因+多基因混合遺傳模型分離分析方法對甜蕎莖稈重心高度和抗折力進行分析,初步闡明其遺傳效應和機制,為加快甜蕎抗倒伏品種選育和降低生產上倒伏率提供理論依據。
1 材料與方法
1.1 試驗材料
酉蕎2號(抗倒伏品種)為本課題組育成品種;烏克蘭大粒蕎(易倒伏品種)由重慶市蕎麥產業技術體系創新團隊提供。種子發芽率均在90%以上。
1.2 試驗方法
試驗材料種植在西南大學歇馬科研基地(106°18′14″~106°56′53″E,10°3′53″~29°39′10″N),試驗田土壤為沙壤土。2014年秋季,在隔離區內配制兩親本正、反交組合,初花期去除異類型花,成熟收獲后得到正、反交種F1代;2015年春季F1自交以及與雙親回交分別獲得F2、B1和B2的種子;2015年秋季,將這2個組合6個世代的種子分別播種進行抗倒伏相關性狀的測定。設置保護行,P1、P2和F1種植4行,B1、B2和F2種植10行,行長3m,行距0.33m,株距0.20m。成熟期在田間隨機選取300株植株,用直尺量取莖稈基部至該莖(含、葉和鞘)平衡支點的距離,記為莖稈重心高度;取基部第2節間,剝除葉鞘,置于SH-100數顯式推拉力計,將植株拉斷瞬間的力記為抗折力。
1.3 遺傳模型分析
采用蓋鈞鎰等[20]提出的 P1、P2、F1、B1、B2、F2多世代聯合遺傳分析方法,運用章元明等編寫的SIN-A.EXE和SIN-B1.EXE等13個程序,設定參數esp=0.0001,variance coefficient=2,設計1對主基因(A)、2對主基因(B)、多基因(C)、1對主基因加多基因(D)和2對主基因加多基因(E)共24個模型的AIC值,利用AIC準則選擇AIC值最小或較小的1個或幾個備選模型,再通過一組適合性測驗(U12、U22、U32、nW2和Dn)選擇統計量達到顯著水平個數最少的模型作為最優模型,并估計主基因和多基因效應值和方差等遺傳參數。
2 結果與分析
2.1 莖稈重心高度和抗折力表型分布
不同世代莖稈重心高度和抗折力見表1。從表1中可以看出,雙親的莖稈重心高度和抗折力均存在明顯差異,正反交F1、B1、B2和F2莖稈重心高度和抗折力均介于雙親之間,但B1和B2這2個回交世代的莖稈重心高度和抗折力更偏向于回交親本,F2代的莖稈重心高度和抗折力更偏向于高親。從變異系數看,正交中,莖稈重心高度的不分離世代群體(P2和F1)的變異系數低于分離世代群體(B1和B2),說明B1和B2群體的遺傳多態性較高,但F2群體遺傳多態性較低;莖稈抗折力的不分離世代群體(P1、P2和F1)的變異系數高于分離世代群體(B1和B2),說明分離世代群體的遺傳多態性較低。反交中,莖稈重心高度的不分離世代群體(P1和F1)的變異系數高于分離世代群體(B1、B2和F2),說明分離世代群體的遺傳多態性較低;莖稈抗折力的不分離世代群體(P2和F1)的變異系數低于分離世代群體(B1和B2),說明分離世代群體的遺傳多態性較高。

表1 正、反交組合6個世代的莖稈重心高度與抗折力Table 1 Culm gravity height and snapping resistance in the six generations from reciprocal combinations
從F2代正、反交莖稈重心高度和抗折力的次數分布(圖1和圖2)可以看出,正、反交莖稈重心高度和抗折力均呈連續性分布,具有典型的數量遺傳特征。正交F2世代莖稈重心高度具有正態分布現象,表明F2世代可能無主基因存在,屬于受多基因遺傳效應影響的數量性狀;正、反交F2世代莖稈抗折力具有明顯的多峰現象,表明莖稈抗折力性狀屬于受主基因遺傳效應影響的數量性狀。

圖1 F2世代莖稈重心高度的次數分布Fig.1 Frequency distributions of culm gravity height in F2generations

圖2 F2世代莖稈抗折力的次數分布Fig.2 Frequency distributions of culm snapping resistance in F2generations
2.2 莖稈重心高度和抗折力主基因+多基因遺傳分析
利用植物數量性狀主基因+多基因混合遺傳模型分析方法對正、反交組合的莖稈重心高度和抗折力進行基因聯合分析,共獲得24種遺傳模型的極大對數似然函數值和AIC值(表2和表3)。根據AIC準則,選取AIC值最小及與最小AIC值比較接近的3組遺傳模型作為備選模型。在正交組合,D-0、E-0和E-1為莖稈重心高度遺傳的備選模型,B-1、E-0和E-1為莖稈抗折力遺傳的備選模型;在反交組合,C-0、D-0、E-1為莖稈重心高度遺傳的備選模型,B-1、E-0和E-1為莖稈抗折力遺傳的備選模型。
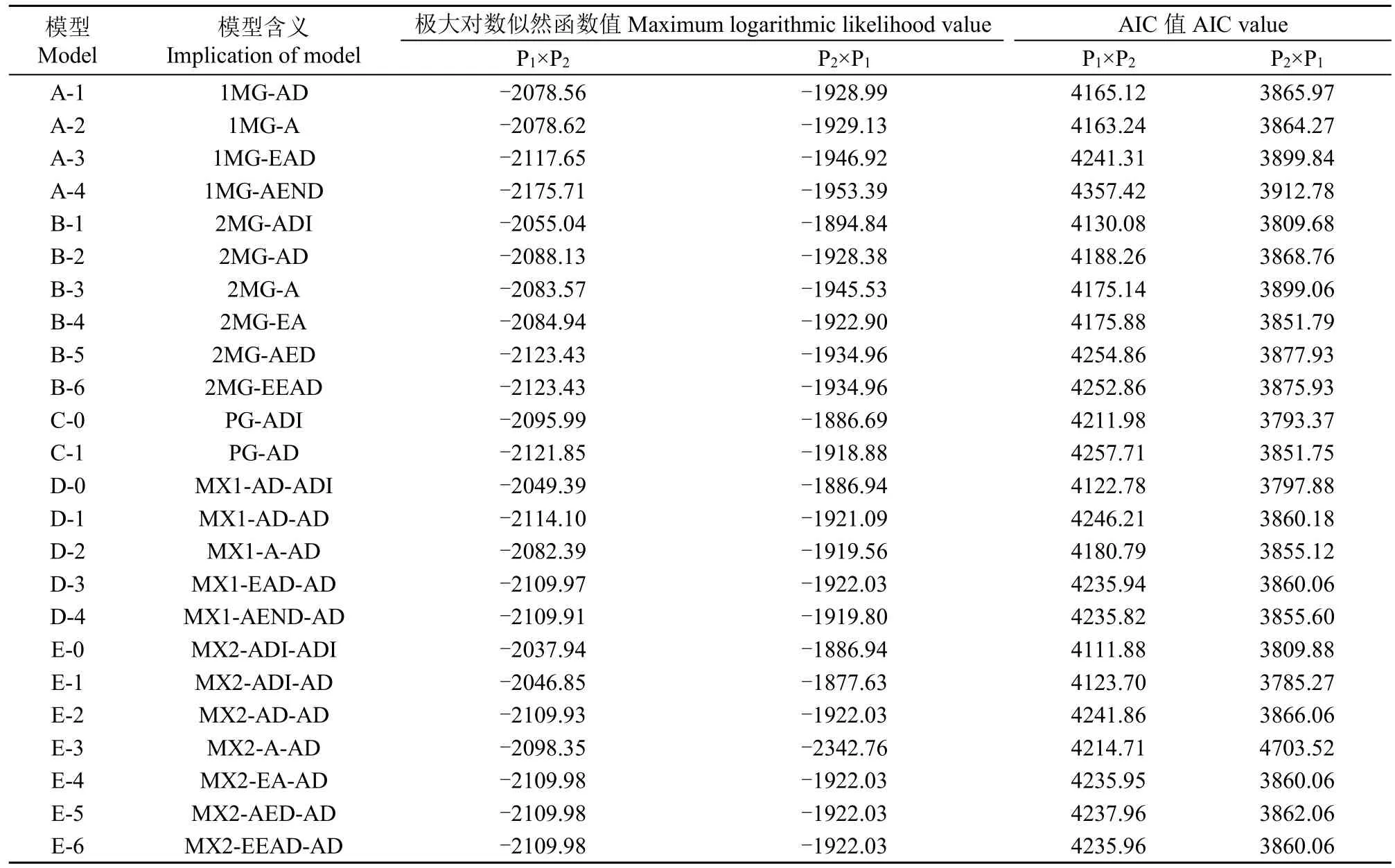
表2 正、反交組合莖稈重心高度性狀的AIC值和極大對數似然函數值Table 2 AIC values and maximum log likelihood estimated values for culm gravity height in reciprocal crosses
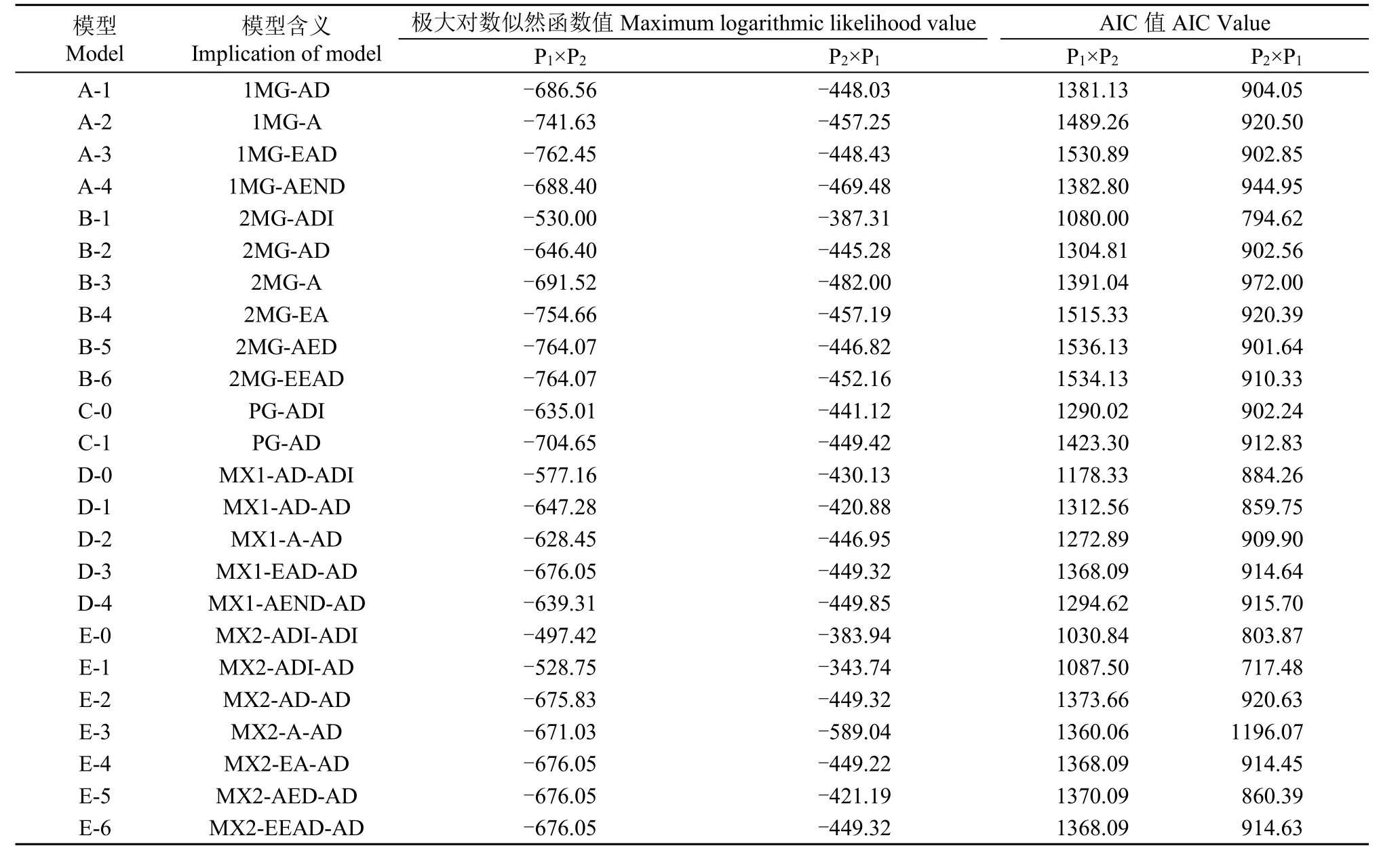
表3 正、反交組合莖稈抗折力性狀的AIC值和極大對數似然函數值Table 3 AIC value and maximum log likelihood estimated values for culm snapping resistance in reciprocal crosses
對備選模型進行適合性測驗,選擇AIC值較小和統計量達到顯著水平個數最少的模型為最適模型(表4和表5),對正交組合,莖稈重心高度的最佳遺傳模型為D-0模型,即1對加性-顯性主基因+加性-顯性-上位性多基因模型,莖稈抗折力的最佳遺傳模型為E-0模型,即2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型;對反交組合,莖稈重心高度和抗折力的最佳遺傳模型均為E-1模型,即2對加性-顯性-上位性主基因+加性-顯性多基因模型。

表4 正、反交組合后代莖稈重心高度備選遺傳模型的適合性檢驗Table 4 Test for goodness-fit of selected genetic model for culm gravity height in reciprocal crosses

表5 正、反交組合后代莖稈抗折力備選遺傳模型的適合性檢驗Table 5 Test for goodness-fit of selected genetic model for culm snapping resistance in reciprocal crosses
2.3 莖稈重心高度和抗折力最適遺傳模型遺傳參數估算
根據選擇的最佳模型D-0、E-0、E-1得出的極大似然估計值計算得到正反交莖稈重心高度和抗折力的一階(表6)和二階遺傳參數(表7)。正交組合中控制莖稈重心高度的1對主基因加性效應大于顯性效應,且作用相反,主基因遺傳率僅在B1世代存在,為10.00%,選擇率低,多基因遺傳率在F2世代中最高,為48.00%,選擇率較低,各群體的環境變異占表型變異的51.21%~100.00%,平均為78.40%,遺傳變異占表型變異的0.00%~48.79%,平均為21.60%,說明環境對正交莖稈重心高度性狀影響極大。反交組合中控制莖稈重心高度的2對主基因加性效應相等(da=db=-3.30),顯性效應分別為ha=0.08,hb=-2.01,小于加性效應,ha/da=-0.02,hb/db=0.61,表現為加性,加性效應的互作效應i為-2.75,顯性效應的互作效應l為-1.79,第1對主基因加性效應與第2對主基因顯性效應的互作效應jab=3.34,第2對主基因加性效應與第1對主基因顯性效應的互作效應jba=5.44,效應值最大。多基因的加性效應為-1.76,顯性效應為-0.50,小于主基因的加性效應和顯性效應,且作用相反,主基因遺傳率在B1世代中最高,為80.22%,選擇率高,多基因遺傳率僅在B2世代存在,為6.20%,各群體的環境變異占表型變異的41.28%~54.00%,平均為47.57%,遺傳變異占表型變異的46.00%~58.72%,平均為52.43%,說明環境對甜蕎莖稈重心高度性狀影響較大。

表6 莖稈重心高度和抗折力一階遺傳參數估計值Table 6 Estimates of first order genetic parameters for culm gravity height and snapping resistance in reciprocal crosses

表7 莖稈重心高度和抗折力二階遺傳參數估計值Table 7 Estimates of second order genetic parameters for culm gravity height and snapping resistance in reciprocal crosses
正交組合中控制莖稈抗折力的2對主基因加性效應相等(da=db=0.59),顯性效應相近,分別為ha=-0.17,hb=-0.14,小于加性效應,ha/da=-0.29,hb/db=0.23,表現為加性,加性效應的互作效應i為0.56,效應值最大,顯性效應的互作效應l為0.23,第1對主基因加性效應與第2對主基因顯性效應的互作效應jab=0.16,第2對主基因加性效應與第1對主基因顯性效應的互作效應jba=-0.15,主基因遺傳率在F2世代高,為86.13%,選擇率高,多基因遺傳率在各世代均不存在,各群體的環境變異占表型變異的14.49%~58.82%,平均為39.59%,遺傳變異占表型變異的41.18%~85.51%,平均為60.41%,說明環境對甜蕎莖稈抗折力性狀有一定的影響。反交組合中控制莖稈抗折力的2對主基因加性效應也相等(da=db=-0.40),顯性效應分別為ha=0.32,hb=0.10,小于加性效應,ha/da=-0.81,hb/db=-0.26,表現為加性。加性效應的互作效應i為0.43,顯性效應的互作效應l為-0.81,效應值最大,第1對主基因加性效應與第2對主基因顯性效應的互作效應jab=0.20,第2對主基因加性效應與第1對主基因顯性效應的互作效應jba=0.73,同時,受多基因效應的影響,多基因的加性效應為0.34,顯性效應為0.51,大于主基因顯性效應,且作用相同。主基因遺傳率在F2世代中最高,為91.75%,選擇率高,所以在F2群體中選擇效率最好,多基因遺傳率在B2世代最高,為13.43%,各群體的環境變異占表型變異的17.24%~21.74%,平均為19.94%,遺傳變異占表型變異的78.26%~82.76%,平均為80.06%,說明環境對甜蕎莖稈抗折力性狀影響不大。
3 討論
作物的抗倒伏性狀表現為復雜的數量性狀遺傳特征。Stojsin等[21]研究表明,玉米莖稈倒伏性狀以加性和顯性效應為主,且加性效應的估計值較高;李得孝等[22]研究與玉米倒伏相關的9個莖稈性狀,遺傳分析結果表明,多數性狀主要受基因的加性和非加性效應,其中莖稈橫折強度等性狀主要受基因非加性效應控制;梁康逕等[23]研究發現,水稻莖稈抗倒性受基因加性、顯性、加性×加性、上位性和與環境互作效應的影響,其中顯性效應與環境互作影響較大;顧慧等[24]研究發現,甘藍型油菜抗壓力性狀的遺傳主要受2對主基因控制,伴有多基因的修飾作用但效應較小,2對主基因都以加性效應為主,主基因間存在明顯的互作效應,F2群體的主基因遺傳率為54.71%,多基因遺傳率只在B1群體中檢測到,為10.56%,遺傳變異平均占表型變異的53.43%,環境變異平均占表型變異的46.57%;李洪戈等[25]研究發現,甘藍型油菜莖稈強度性狀受2對主基因和微效多基因共同控制,以主基因遺傳為主;目前國內外對甜蕎抗倒伏性狀的遺傳分析較少,李英雙等[26]對甜蕎的株高和莖粗的遺傳分析發現,株高和莖粗的遺傳主要受2對加性-顯性-上位性主基因控制,同時伴有加性-顯性多基因的修飾作用,株高遺傳率在80%以上,莖粗遺傳率在80%以下。
本研究表明,甜蕎莖稈重心高度主要受加性和顯性效應控制,且加性效應大于顯性效應,主基因遺傳率大于多基因遺傳率,主基因遺傳率在B1世代中最高,各群體的環境變異占表型變異的49.32%~77.00%,平均為62.98%,遺傳變異占表型變異的33.00%~50.68%,平均為37.02%,說明環境對甜蕎莖稈重心高度性狀的影響極大。這與張倩[27]對油菜莖稈重心高度主要受顯性效應控制,不受加性效應影響的遺傳分析結果不同。甜蕎莖稈抗折力主要以加性效應為主,2對主基因間存在明顯的基因互作效應,主基因遺傳率大于多基因遺傳率,多基因遺傳率在F2世代沒有檢測到,主基因遺傳率在F2世代最高,為88.94%,選擇率高,應在早期世代進行選擇,各群體的環境變異占表型變異的18.11%~38.03%,平均為29.76%,遺傳變異占表型變異的61.97%~81.89%,平均為60.24%,說明環境對甜蕎莖稈抗折力性狀有一定的影響,這與顧慧等[24]對油菜抗壓力性狀的遺傳主要受2對主基因控制且伴有多基因修飾、主基因以加性效應為主、多基因遺傳率只在B1群體中檢測到、遺傳變異大于環境變異的研究結果相似。
4 結論
莖稈重心高度最佳遺傳模型為1對加性-顯性主基因+加性-顯性-上位性多基因模型和2對加性-顯性-上位性主基因+加性-顯性多基因模型,以加性效應為主,主基因遺傳率大于多基因遺傳率,環境變異大于遺傳變異,降低甜蕎莖稈重心高度可以有效提高抗倒伏能力。
莖稈抗折力性狀的最佳遺傳模型為2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型和2對加性-顯性-上位性主基因+加性-顯性多基因模型,以加性效應為主,2對主基因間存在明顯的基因互作效應,主基因遺傳率大于多基因遺傳率,多基因遺傳率在F2世代沒有檢測到,主基因遺傳率在F2世代最高,為88.94%,選擇率高,在甜蕎抗倒伏品種選育上可在早期世代進行選擇,來減少工作量和提高育種效率。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
