擬南芥MGT6調控高鎂條件下Ca2+/Mg2+動態平衡的機制研究
2022-09-13 08:15:28易世鳴王春平周冰玉孫翔宇
生命科學研究 2022年3期
關鍵詞:植物
易世鳴,王春平,李 慧,周冰玉,孫翔宇
(湖南省微生物研究院,中國湖南 長沙 410009)
鎂離子(Mg2+)是植物細胞質中最豐富的游離二價陽離子,在植物的生長發育中有著十分重要的作用[1~3]。它在植物體中能夠作為許多酶的輔助因子參與一系列生理生化反應,同時也作為葉綠素的中心元素參與光合作用[4~8]。
缺鎂引起淀粉和蔗糖等碳源在植物葉片中積累,高濃度的蔗糖能夠抑制編碼葉綠素a/b結合蛋白2的基因CAB2的表達,影響植物光合作用[9~10]。但植物細胞中過量的Mg2+也會抑制植物正常生長發育。當植物中Mg2+過量時,葉片的葉緣部分焦枯,葉尖萎縮泛黃,在更嚴重的情況下,整個葉片組織全部淡黃,葉肉組織逐漸變為褐色,直至壞死,導致植株的生長發育受阻[11]。高鎂脅迫嚴重影響作物生長[12]。但目前的研究主要針對植物缺鎂生理,而對高鎂毒害引起的植物生理變化及其機理卻了解較少。
有研究表明,濃度較高的Mg2+會促進細胞外部鈣離子(Ca2+)被Mg2+所取代,影響細胞壁的穩定性和細胞質膜的滲透性,從而影響植物生長發育[13]。但到目前為止,有關植物細胞鈣鎂離子(Ca2+/Mg2+)動態平衡的機制報道較少。
鎂轉運蛋白6(magnesium transporter 6,MGT6)是植物中第一個被報道定位于細胞質膜,并在低鎂條件下介導植物根部吸收Mg2+的蛋白質[14]。本研究發現,MGT6的突變體和RNA干擾(RNA interference,RNAi)植株在高鎂條件下出現葉片變黃的高鎂敏感表型,并且該表型能被一定濃度的Ca2+拮抗,說明擬南芥MGT6能通過調控高鎂脅迫下鈣鎂離子動態平衡來維持植物正常生長發育。該研究將為解析植物響應高鎂脅迫的信號網絡奠定基礎。
1 材料與方法
1.1 植物材料
實驗所用的材料為擬南芥(Arabidopsisthaliana)野生型(Columbia生態型,Col-0),種子購買自擬南芥生物資源中心(Arabidopsis Biological Resource Center,US)。擬南芥MGT6基因(At3g58970)的mgt6 T-DNA插入突變體(Salk_203866C)和MGT6 RNAi(#2、#8、#12)種子均為本實驗室保存,并已被鑒定為純合突變體。
1.2 培養基的配制
MS固體培養基(100 mL):4 mL 25×A液,1 mL 100×B-C液,1 mL 100×D液,1 g蔗糖,雙蒸水定容至100 mL;用10 mol/L NaOH溶液調節pH至5.8,再加入0.8 g(0.9%)擬南芥培養專用瓊脂,121℃高溫滅菌2 h。培養基A液、B-C液和D液參照文獻[14]配制。
1/6 MS固體培養基(300 mL):2 mL 25×A液,0.5 mL 100×B-C液,0.5 mL 100×D液,3 g蔗糖,雙蒸水定容至300 mL;調節pH至5.8,再加入2.8 g(0.9%)擬南芥培養專用瓊脂,121℃高溫滅菌2 h。
1.3 擬南芥溫室培養
用75%乙醇處理擬南芥相關種子5 min,進行表面消毒。春化處理后,將其點植于MS培養基上,于光照培養箱豎直放置培養,培養條件:溫度(22±1)℃;光暗周期 16 h(光照)/8 h(黑暗)。
1.4 高鎂脅迫處理
將相關擬南芥種子分別種于0 mmol/L、0.01 mmol/L、0.1 mmol/L、3 mmol/L、6 mmol/L、8 mmol/L Mg2+濃度(用1 mol/L Mg2+母液加入相應體積的去離子水配制,1 mol/L Mg2+母液用12 g MgSO4加入100 mL去離子水配制)的1/6 MS固體培養基,用封口膜將培養基封口后放置在溫度為(22±1)℃、光暗周期為16 h光照/8 h黑暗的溫室中豎直培養。培養14 d后,對野生型植株、mgt6突變體及RNAi植株進行表型觀察,并統計植株根長和鮮重。文中將Mg2+濃度>3 mmol/L定義為高鎂條件。
1.5 Ca2+拮抗處理
配制不同 Mg2+濃度(0 mmol/L、0.01 mmol/L、0.1 mmol/L、3 mmol/L、6 mmol/L、8 mmol/L)的 1/6 MS固體培養基,并分別加入1 mmol/L或3 mmol/L Ca2+(用1 mol/L Ca2+母液加入相應體積的去離子水配制,1 mol/L Ca2+母液用 21.9 g CaCl2·6H2O 加入100 mL去離子水配制而成)。將擬南芥野生型種子、mgt6突變體和MGT6 RNAi植株(#8)的種子分別種在上述1/6 MS固體培養基上,用封口膜將培養基封口后放置在溫度為(22±1)℃、光暗周期為16 h光照/8 h黑暗的溫室中豎直培養7 d。觀察表型,并統計植株根長和鮮重。
1.6 統計學處理
所有數據用平均值±標準差()表示。多組間比較采用單因素方差分析,顯著性水平為P<0.05。采用Sigma Plot計算機軟件制圖。
2 結果
2.1 高鎂條件下MGT6突變體及其RNAi植株的表型觀察與分析
將野生型(WT)、mgt6突變體和MGT6 RNAi(#2、#8、#12)種子分別種于含不同 Mg2+濃度(0 mmol/L、0.01 mmol/L、0.1 mmol/L、3 mmol/L、6 mmol/L、8 mmol/L Mg2+)的1/6 MS固體平板上,豎直培養14 d后進行表型觀察。結果顯示:在3 mmol/L Mg2+濃度下,mgt6突變體植株和MGT6 RNAi植株的生長表型與野生型一致;但在高Mg2+濃度(6 mmol/L和8 mmol/L)下,mgt6突變體植株和MGT6 RNAi植株出現葉片變黃、地上部分變小的高鎂表型(圖1),同時,根長和鮮重均較野生型明顯降低(圖2)。上述結果表明,MGT6參與植物對高鎂脅迫的響應。

圖1 不同Mg2+濃度下的擬南芥表型分析(標尺:1 cm)Fig.1 Phenotype analysis of Arabidopsis thaliana under different Mg2+conditions(scale bar:1 cm)
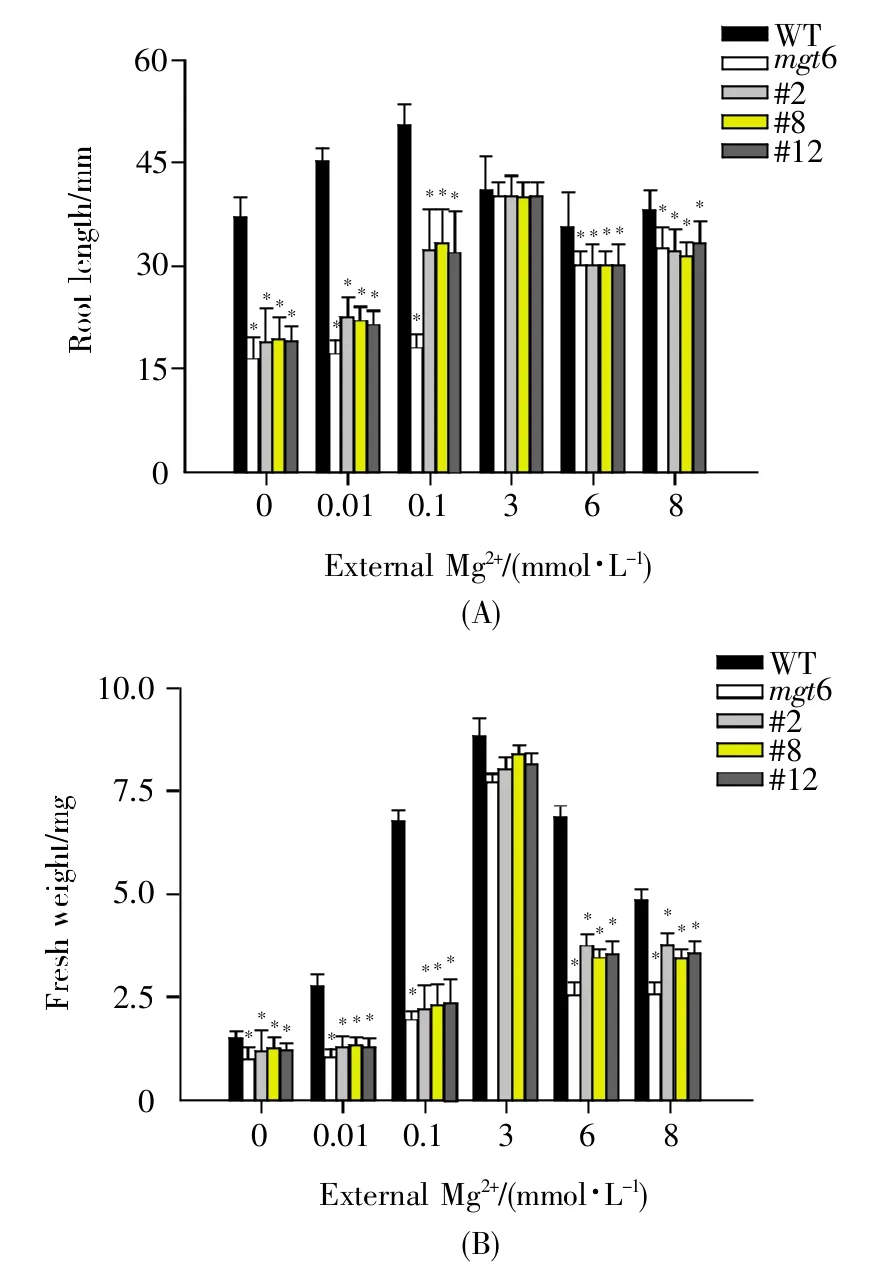
圖2 不同Mg2+濃度下的根長和鮮重分析(10個植株)(A)根長;(B)鮮重。*代表與WT相比有顯著性差異(P<0.01)。Fig.2 Root length and fresh weight analysis of Arabidopsis thaliana under different Mg2+conditions(10 plants)(A)Root length;(B)Fresh weight.Asterisks represent significant difference compared with WT(*P<0.01 by Student’s t test).
2.2 Ca2+拮抗MGT6突變體及其RNAi植株高鎂表型的分析
有研究表明Mg2+能和Ca2+互作來調控植物生長發育[14]。為了驗證高鎂條件下擬南芥MGT6是否能通過調節Ca2+/Mg2+動態平衡來維持植物正常生長發育,我們將擬南芥哥倫比亞野生型(WT)、mgt6突變體和隨機挑選的MGT6 RNAi(#8)種子分別種在含不同Mg2+濃度(0 mmol/L、0.01 mmol/L、0.1 mmol/L、3 mmol/L、6 mmol/L 和 8 mmol/L)的1/6 MS固體培養基上,且在各培養基中分別添加3 mmol/L Ca2+,豎直培養7 d后進行表型觀察。結果顯示:在低鎂條件(0 mmol/L、0.01 mmol/L、0.1 mmol/L Mg2+)下,加入3 mmol/L Ca2+后,mgt6突變體植株和MGT6 RNAi(#8)植株的低鎂表型不能得到恢復;但在高鎂條件(6 mmol/L、8 mmol/L Mg2+)下,加入3 mmol/L Ca2+后,mgt6突變體植株和MGT6 RNAi(#8)植株的高鎂表型得到緩解,葉片開始變綠,各植株長勢均恢復至野生型水平(圖3)。進一步的統計學分析顯示,mgt6突變體植株和MGT6 RNAi(#8)植株的根長和鮮重均恢復到野生型水平(圖4)。上述結果表明,Ca2+不能緩解mgt6突變體植株及MGT6 RNAi植株的低鎂表型,但能夠有效拮抗mgt6突變體及RNAi植株的高鎂敏感表型。

圖3 Ca2+拮抗mgt6突變體及RNAi植株的高鎂表型Fig.3 Phenotype analysis of mgt6 mutants and RNAi plants under high-Mg2+conditions after Ca2+supplement
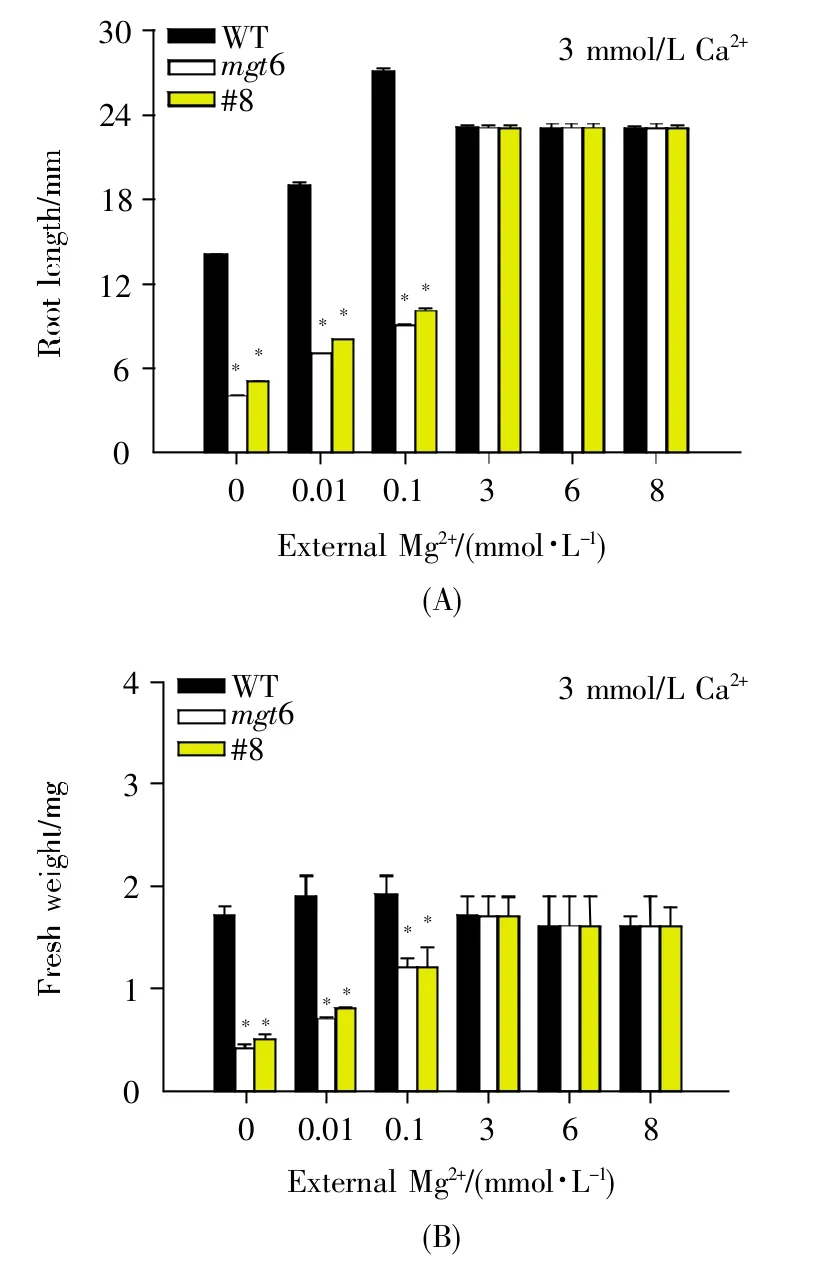
圖4 Ca2+拮抗mgt6突變體及RNAi植株高鎂表型的分析(5個植株)(A)根長;(B)鮮重。*代表與WT相比有顯著性差異(P<0.01)。Fig.4 Phenotype analysis of mgt6 mutants and RNAi plants under high-Mg2+conditions after Ca2+supplement(5 plants)(A)Root length;(B)Fresh weight.Asterisks represent significant difference compared with WT(*P<0.01 by Student’s t test).
2.3 Ca2+拮抗MGT6突變體及其RNAi植株高鎂表型的細胞學分析
為了探究Ca2+拮抗mgt6突變體及MGT6 RNAi植株高鎂表型的機制,我們進一步進行了mgt6突變體及RNAi植株在不同濃度Ca2+處理下的細胞學分析。結果顯示:高鎂脅迫(6 mmol/L Mg2+)處理下,mgt6突變體和MGT6 RNAi(#8)植株的葉肉細胞形態不規則、皺縮(圖5),且較野生型明顯縮小,僅為野生型葉肉細胞大小的49.1%(圖6);但當在高鎂培養基(6 mmol/L Mg2+)中加入1 mmol/L Ca2+后,mgt6突變體和MGT6 RNAi(#8)植株的葉肉細胞形態明顯變規則,皺縮現象減輕(圖5),同時,葉肉細胞大小也逐漸得到恢復,恢復至野生型葉肉細胞大小的57.2%(圖6);當在高鎂培養基(6 mmol/L Mg2+)中加入 3 mmol/L Ca2+后,mgt6 突變體和MGT6 RNAi(#8)植株的葉肉細胞形態、大小均逐漸恢復至野生型水平(圖5,圖6)。上述結果進一步說明,mgt6突變體和MGT6 RNAi(#8)植株的高鎂敏感表型能被一定濃度的Ca2+拮抗,MGT6能通過調控細胞鈣鎂離子動態平衡來響應高鎂脅迫。
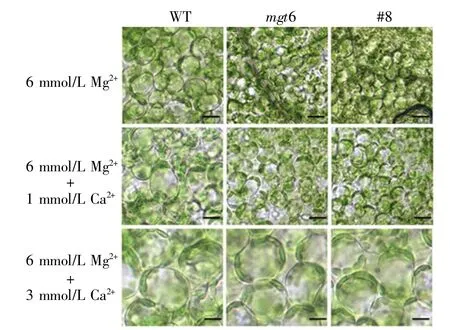
圖5 高鎂條件下不同濃度Ca2+對mgt6突變體及RNAi植株葉肉細胞形態的影響(標尺:10 μm)Fig.5 Influence of different Ca2+concentrations on the morphology of mesophyll cells of mgt6 mutants and RNAi plants treated with a high Mg2+concentration(scale bar:10 μm)

圖6 不同濃度Ca2+對高鎂(6 mmol/L Mg2+)處理下mgt6突變體及RNAi植株葉肉細胞大小的影響*代表與WT相比有顯著性差異(P<0.01)。Fig.6 Influence of different Ca2+concentrations on the size of mesophyll cells of the mgt6 mutants and RNAi plants treated with a high Mg2+concentration(6 mmol/L)Asterisks represent significant difference compared with WT(*P<0.01 by Student’s t test).
3 討論
鎂是植物生長發育所必需的營養元素之一,在植物的生長發育中起著非常重要的作用。但到目前為止,有關植物吸收和運輸Mg2+的分子機制報道較少。MGT6是植物中第一個被報道定位于細胞質膜,并在低鎂條件下介導植物根部吸收Mg2+的蛋白質。本研究發現,MGT6的突變體和RNAi植株對高鎂敏感,該高鎂表型能被一定濃度的Ca2+拮抗。這說明MGT6可能通過調控鈣鎂離子動態平衡來響應高鎂脅迫。
Ca2+和Mg2+是植物中十分豐富的二價陽離子,二者在植物細胞中可能具有拮抗作用,因此植物通過鈣鎂離子含量之間的平衡以獲得最適的生長和發育。維持這種平衡以應對土壤營養變化是植物礦物營養的重要特征。研究發現,植物能感知營養變化并經過信號轉導來調節轉運蛋白活性,從而適應環境營養變化[13~14]。目前,人們已鑒定出一些離子通道和轉運蛋白家族,它們幫助Ca2+、Mg2+跨質膜和細胞內膜轉運。在細胞和植株水平協調這些轉運系統的活性可控制Ca2+和Mg2+的營養水平[11]。植物中鈣鎂離子含量的平衡緊密相連,并且可能(至少部分)由共同的信號網絡調節。但到目前為止,研究人員對植物通過鈣鎂平衡來響應高鎂脅迫的分子機制還了解甚少。
本研究發現,擬南芥鎂轉運體MGT6的突變體mgt6和RNAi植株具有地上部分矮小、葉片發黃、葉鮮重顯著下降的高鎂敏感表型,這種高鎂表型能被外源施加的一定濃度的Ca2+互補。葉片細胞顯微結構的比較分析顯示:高鎂條件下,mgt6突變體及MGT6 RNAi植株的葉肉細胞減小,形態不規則;但隨著培養基中Ca2+濃度的增加,mgt6突變體及RNAi植株的葉肉細胞逐漸增大,細胞形態逐漸變得規則,當在培養基中加入3 mmol/L Ca2+時,突變體及RNAi植株的葉肉細胞大小和形態恢復到野生型水平。這進一步說明,mgt6突變體及MGT6 RNAi植株的高鎂敏感表型能被一定濃度的Ca2+拮抗。
綜上所述,MGT6在高鎂條件下能通過參與植物鈣鎂離子動態平衡來響應高鎂脅迫,維持植物在高鎂脅迫下的正常生長發育。但在高鎂條件下,MGT6誘導出植物細胞特異的Ca2+信號來維持植物Mg2+動態平衡進而響應高鎂脅迫的分子機制還有待進一步研究。另外,對于Ca2+/Mg2+平衡,我們應繼續完善調節和協調土壤營養狀態不斷變化下的Ca2+、Mg2+轉運過程的信號網絡。總的來講,揭示植物Ca2+和Mg2+平衡的分子機制將有助于改善作物的營養特性,從而提高作物產量,最終提高人類的健康水平。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
