谷氨酰胺代謝對人甲狀腺癌細胞系體外增殖能力影響的實驗研究
2022-09-14 00:47:00劉新艷王媛媛劉娜娜
陜西醫學雜志 2022年9期
劉新艷,王媛媛,劉娜娜,王 榮
(1.榆林市第一醫院內分泌科,陜西 榆林 719000;2.榆林市第一醫院全科醫學科,陜西 榆林 719000)
甲狀腺癌是內分泌系統常見的惡性腫瘤,2020年全球癌癥統計報告中指出甲狀腺癌在全球癌癥發病率中排名第9位,占全年新診斷惡性腫瘤的3.0%,女性發病率高于男性3倍,但病死率低,僅為0.5/10萬(女性)和0.3/10萬(男性)[1]。甲狀腺癌也是中國發病率增長速度最快的癌癥之一,2003—2011年,平均每年增加20%[2]。中國癌癥登記報告調查回顧性分析提示,2000—2015年中國女性常見的惡性腫瘤發病率排名依次是乳腺癌、肺癌、結直腸癌、甲狀腺癌、胃癌和宮頸癌[3]。2015年我國甲狀腺癌年齡標準化發病率為9.61/10萬,而年齡標準化病死率為0.35/10萬[4]。甲狀腺癌病理分型包括分化型甲狀腺癌(Differentiated thyroid carcinoma,DTC)、低分化甲狀腺癌(Poorly differentiated thyroid cancer,PDTC)、未分化甲狀腺癌(Anaplastic thyroid cancer,ATC)及甲狀腺髓樣癌(Medullary thyroid carcinoma,MTC)。95%以上甲狀腺癌病理類型是DTC,其臨床常用治療方法包括手術、放射性131I、TSH抑制等[5]。多數DTC患者臨床預后良好,但進展性局部晚期或轉移性碘難治性DTC、PDTC、MTC及ATC患者臨床預后差,通常采取手術、外照射及全身治療。ATC是甲狀腺癌中侵襲性最強、致死率最高的病理類型,患者平均生存期僅6個月,尚缺乏有效的治療手段[6-7]。
腫瘤代謝經典的Warburg 效應指腫瘤細胞即使在氧供充足的情況仍主要依賴糖酵解方式獲取能量,而非葡萄糖有氧氧化方式[8],已證實結直腸癌、乳腺癌、肺癌及甲狀腺癌中存在此現象,隨后葡萄糖代謝處于腫瘤代謝研究的核心地位。然而,并非所有的腫瘤均存在一致的Warburg 效應,Hans Krebs 于1935年首次報道了谷氨酰胺(Glutamine,Gln)的代謝研究,隨后Gln在腫瘤細胞生物學中的作用被逐步揭示[9-10]。腫瘤細胞除糖代謝改變以外,其氨基酸代謝及脂代謝等其他能量代謝方式也發生了變化。Gln作為半必需氨基酸,對于腫瘤的發生與發展至關重要,在多種惡性腫瘤中均發現腫瘤細胞對Gln具有一定的依賴性[11]。然而,Gln代謝在甲狀腺癌中的研究相對滯后,因此本研究通過一系列體外實驗探討Gln代謝對甲狀腺癌細胞體外增殖的影響,期望為甲狀腺癌的治療提供新的思路和策略。
1 材料與方法
1.1 細胞系 本研究所用甲狀腺癌細胞系包括4株ATC細胞系(C643、IHH4、8505C和8305C)和2株甲狀腺乳頭狀癌(PTC)細胞系(K1和TPC-1)。除C643細胞系由上海瑞金雷燁博士饋贈外,其余5株細胞系均由中國醫科大學附屬第一醫院關海霞教授饋贈。
1.2 主要試劑 胎牛血清(貨號:SH30088.03)購于HyClone公司;RPMI-1640 培養基(貨號:11875101)及無Gln RPMI-1640培養基(貨號:21870084)購于Gibco公司;0.25%胰蛋白酶(貨號:C0201-100ml)、ATP濃度檢測試劑盒(貨號:S0026)與凋亡檢測試劑盒(貨號:C1062L)購于碧云天生物技術有限公司;Gln(貨號:A100374)購于上海生工生物。
1.3 甲狀腺癌細胞培養 人甲狀腺癌細胞系C643、IHH4、8505C、8305C、K1和TPC-1常規在含10%胎牛血清的RPMI-1640 培養基中,于37 ℃、5% CO2的細胞培養箱中培養,隔天更換細胞培養液,傳代培養,選取處于對數生長期的甲狀腺癌細胞用于后續實驗。
1.4 噻唑藍(MTT)檢測甲狀腺癌細胞增殖活性 人甲狀腺癌細胞系置于37 ℃、5% CO2、飽和濕度的培養箱中培養至對數生長期,常規消化細胞。全自動細胞計數儀計數,按每孔細胞800個計算所需細胞總數。用排槍加至 96 孔板,每組設置5個復孔,置于培養箱37 ℃、5% CO2條件下培養,待細胞貼壁后用含不同濃度Gln(0、1、2 mmol/L)培養基繼續培養細胞,分別于0、1、2、3、4 d后各收1板,每孔加入MTT液20 μl,于培養箱孵育4 h后吸去上清液,每孔加入二甲基亞砜(DMSO)150 μl,在搖床上低速振蕩15 min以使結晶物充分溶解,在酶標儀570 nm波長下檢測各孔的光密度(OD)值,其中第3天時為最后2 d的96孔板重新補充不同濃度Gln培養基。
1.5 平板克隆形成實驗 取對數生長期甲狀腺癌細胞,常規消化,全自動細胞計數儀計數,按每孔1000個細胞計算所需細胞總數。將細胞種植于6孔板內,輕吹使細胞均勻分布,置于培養箱37 ℃、5% CO2條件下培養,待細胞貼壁后用不同濃度Gln(0、1、2 mmol/L)培養基處理細胞,每4 d更換1次培養基,10~14 d 后顯微鏡下觀察到克隆長成但尚未連續成片狀、大小至肉眼可見時,棄去培養基, PBS洗滌2次,每孔加入1.5 ml甲醇固定15 min。隨后結晶紫染色,用流水洗去殘留的結晶紫,通風干燥后拍照,統計克隆數。
1.6 甲狀腺癌細胞凋亡檢測 取對數生長期甲狀腺癌細胞,常規消化后分別予以不同濃度Gln(0、1、2 mmol/L)培養基繼續培養細胞48 h,隨后消化6孔板細胞,留存培養基(培養基中含凋亡細胞)連同消化后細胞一起離心重懸,控制細胞總數在1×106個左右,每種處理設置3個副孔。通過凋亡檢測試劑盒內的Annexin V-PI染料對目標細胞進行冰上避光染色15 min,同時設置未染組和Annexin V、 PI兩個單染組作為對照組,染色完成的細胞轉移到流式管內,加入0.5 ml經0.22 μm濾器過濾的PBS,上流式細胞儀分析,以488 nm波長激發,經515 nm濾光片過濾后檢測熒光素,用>600 nm濾光片檢測PI。應用Flowjo 7.6軟件進行數據分析。
1.7 甲狀腺癌細胞三磷酸腺苷(ATP)生產效能檢測 甲狀腺癌細胞予以不同濃度Gln(0、1、2 mmol/L)培養基處理24、48、72 h后常規消化6孔板細胞,利用ATP濃度檢測試劑盒中ATP檢測裂解液裂解細胞,催化螢光素產生螢光時需要ATP提供能量,從而用熒光分光光度計測定ATP濃度,用細胞總蛋白水平校正ATP濃度值。

2 結 果
2.1 不同濃度Gln對甲狀腺癌細胞增殖活性的影響 見圖1。與含有Gln的培養基相比,不含Gln培養基培養的6株甲狀腺癌細胞增殖活性明顯降低(均P<0.05)。

圖1 不同濃度Gln對甲狀腺癌細胞增殖活性的影響注:與Gln 0 mmol/L比較,*P<0.05,#P<0.01,▲P<0.001
2.2 不同濃度Gln對甲狀腺癌細胞平板克隆形成能力的影響 見圖2。與含有Gln的培養基相比,不含Gln培養基培養的6株甲狀腺癌細胞細胞平板克隆形成能力明顯降低(均P<0.05)。
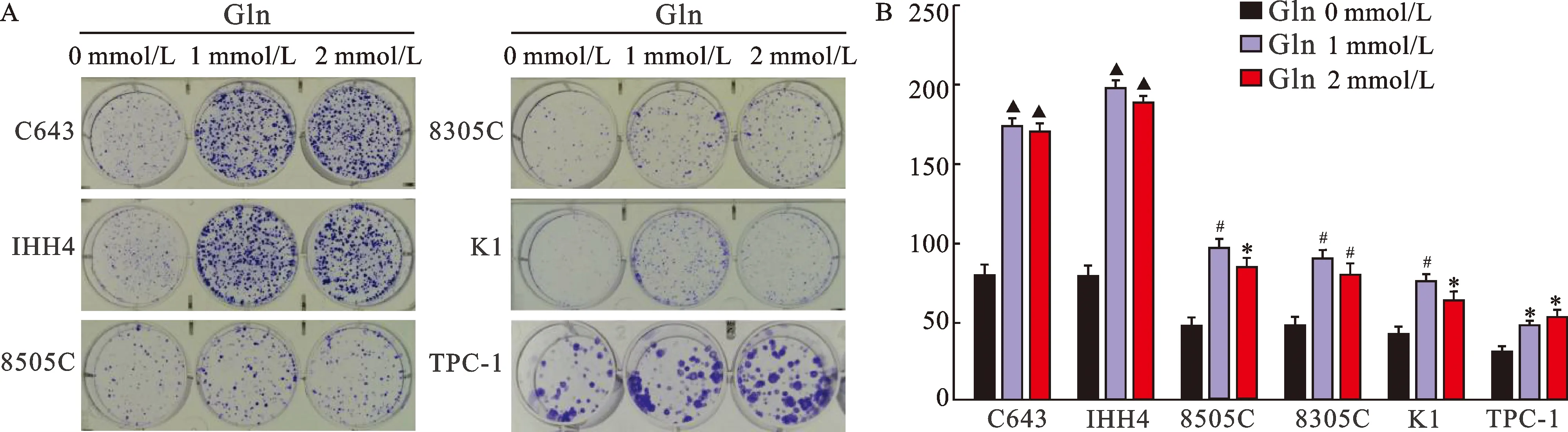
圖2 不同濃度Gln對甲狀腺癌細胞平板克隆形成能力的影響注:圖A為6株甲狀腺癌細胞平板克隆形成實驗(結晶紫染色);圖B中,與Gln 0 mmol/L比較,*P<0.05,#P<0.01,▲P<0.001
2.3 不同濃度Gln對甲狀腺癌細胞凋亡的影響 見圖3。與含有Gln的培養基相比,不含Gln培養基培養的6株甲狀腺癌細胞凋亡明顯增加(均P<0.05)。
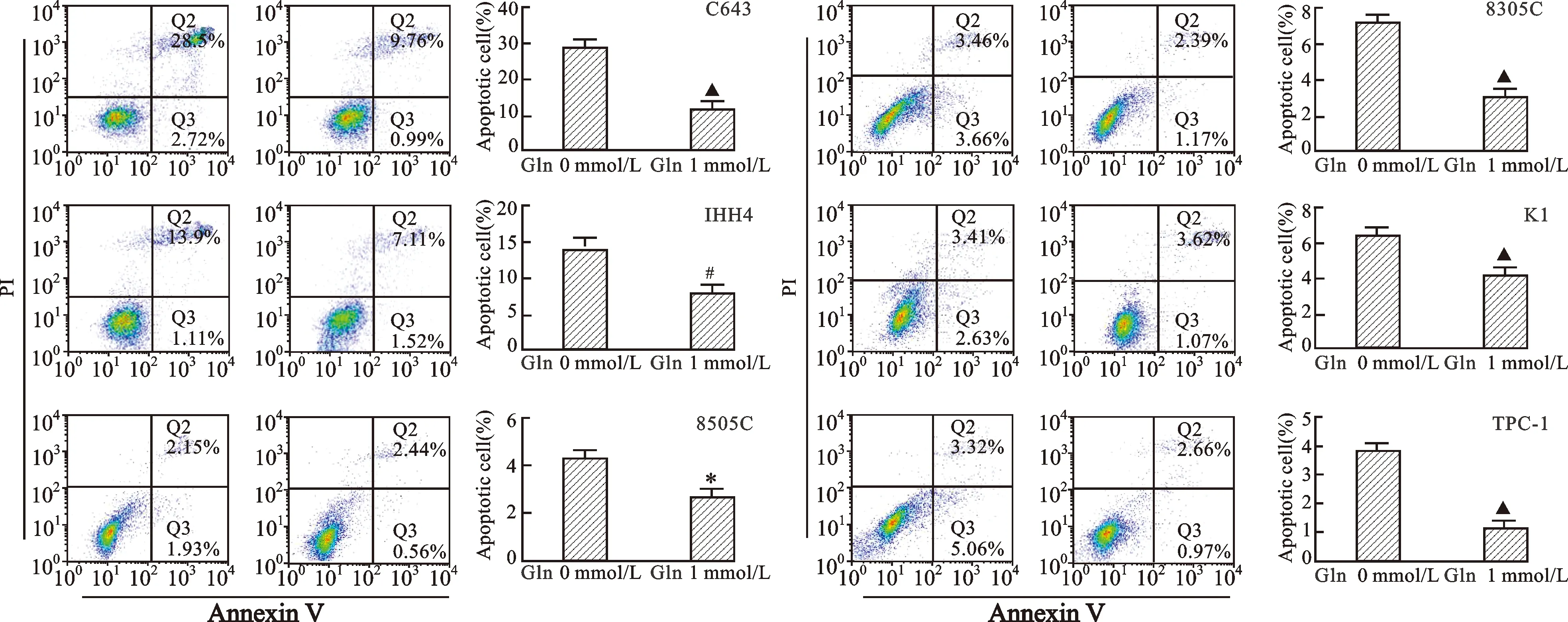
圖3 不同濃度Gln對甲狀腺癌細胞凋亡的影響(Annexin V-PI染色)注:與Gln 0 mmol/L比較,*P<0.05,#P<0.01,▲P<0.001
2.4 不同濃度Gln對甲狀腺癌細胞ATP生成能力的影響 見圖4。與含有Gln的培養基相比,不含Gln培養基培養的6株甲狀腺癌細胞ATP生成能力明顯降低(均P<0.05)。
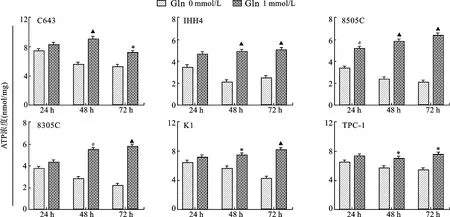
圖4 不同濃度Gln對甲狀腺癌細胞產生ATP能力的影響注:與Gln 0 mmol/L比較,*P<0.05,#P<0.01,▲P<0.001
3 討 論
甲狀腺癌是內分泌系統常見惡性腫瘤,也是中國發病率增長速度較快的腫瘤之一,目前已成為我國女性第四大惡性腫瘤。雖然臨床多數甲狀腺癌預后良好,但轉移性或碘難治性甲狀腺癌以及低分化或未分化甲狀腺癌尚缺乏有效的治療手段,進而導致患者臨床預后差,尤其是ATC患者1年生存率不足20%。甲狀腺癌致死率低,但發病率高,因此仍需探尋更為有效、安全的治療靶點和策略。
自腫瘤Warburg 效應被揭示以來,葡萄糖一直處于腫瘤代謝研究的中心地位。Hans Krebs于1935年研究了動物體內Gln代謝,進而掀起以Gln代謝為腫瘤治療靶點的研究熱潮。Gln屬于非必需氨基酸,也是人體最豐富的氨基酸,占血液中游離氨基酸庫的20%以上,占肌肉中游離氨基酸庫的40%以上。人體Gln始終保持恒定水平,Gln分解代謝參與維持機體酸堿平衡及氨基酸代謝平衡。雖然Gln為機體非必需氨基酸,但在腫瘤細胞中則處于必需氨基酸地位。腫瘤細胞快速增殖需要消耗大量能量,Gln通過多種跨膜轉運體進入腫瘤細胞,再經過線粒體Gln酶水解為谷氨酸和氨離子,隨后谷氨酸經過谷氨酸脫氫酶催化生成α-酮戊二酸(α-KG),后者則進入三羧酸(TCA)循環,最終產生ATP為腫瘤細胞的快速增殖提供能量。Gln代謝產生的碳用于氨基酸和脂肪酸的合成,而Gln代謝產生的氮直接用于嘌呤和嘧啶的從頭生物合成[12]。谷氨酸還與半胱氨酸、甘氨酸通過結合的方式生成谷胱甘肽(GSH)參與腫瘤細胞氧化還原反應,直接抑制活性氧(ROS)合成,抑制腫瘤細胞凋亡,GSH水平還與腫瘤的發生和耐藥性相關[13-14]。已有研究[15-16]證實,致癌基因突變上調腫瘤細胞Gln代謝,其中研究最多的是Myc致癌基因,Myc可以上調Gln轉運體(SLCIA5),誘導Gln酶(GLS)在mRNA和蛋白水平上的表達,并在缺氧條件下促進Gln回補TCA循環和誘導GSH的產生。研究[17]證實PIK3CA突變直腸癌細胞通過上調谷氨酸丙酮酸轉氨酶2(GPT2)來提高Gln代謝,使PIK3CA突變直腸癌細胞生長更加依賴Gln,提示靶向Gln代謝可能是治療PIK3CA突變型結直腸癌的一種有效方法。另外,谷氨酸還可通過多種途徑激活mTOR信號通路以促進腫瘤細胞生長[18]。
已知部分腫瘤的發生進展依賴于Gln,但Gln代謝是否參與了甲狀腺癌尤其是ATC的發生與發展仍不明確。研究[19]發現PTC細胞生長依賴于Gln,其次以Gln代謝為治療靶點的GLS抑制劑可以抑制PTC細胞的增殖、侵襲、遷移等惡性行為,提示Gln代謝可能是PTC的潛在治療靶點。沉默調節蛋白(SIRT)4是Sirtuin家族成員,具有去乙酰化酶活性并在能量代謝中發揮作用,已有研究[20]證實SIRT4通過抑制Gln代謝進而削弱甲狀腺癌細胞的增殖、遷移和侵襲能力。此次研究我們利用甲狀腺癌細胞體外實驗證實甲狀腺癌細胞的體外培養依賴Gln的供給,當剝奪腫瘤細胞培養基中Gln后會顯著抑制甲狀腺癌細胞增殖活性、平板克隆形成能力,并顯著誘導甲狀腺細胞發生凋亡。已知Gln為快速分裂的腫瘤提供能量、碳源及氮源,本研究將甲狀腺癌細胞置于不同濃度Gln培養基中培養24~72 h,結果顯示剝奪腫瘤細胞培養基中Gln后,ATP產生能力顯著降低,提示甲狀腺癌細胞的Gln依賴可能更多源于腫瘤所需能量不足。腫瘤細胞生長對于Gln的依賴使其成為極具潛力的抗腫瘤治療靶點,以Gln代謝為目標的化合物已被應用于臨床前期實驗并顯示出效果的有GLS抑制劑BPTES和CB-839,可分別抑制基因工程小鼠癌癥模型的生長和三陰性乳腺癌的進展[21-22]。腫瘤的異質性、不同腫瘤的基因類型和微環境對靶向治療提出了相當大的挑戰,研究顯示靶向Gln代謝還可與其他藥物組合誘導腫瘤合成致死,如抗凋亡蛋白Bcl-2特異性抑制與GLS抑制協同作用可誘導急性髓細胞白血病細胞發生凋亡及周期阻滯[23]。
綜上所述,甲狀腺癌細胞體外生長依賴Gln,剝奪Gln后甲狀腺癌細胞體外增殖、平板克隆形成能力、ATP生成能力受到抑制,細胞凋亡增加,靶向Gln代謝可能成為甲狀腺癌尤其是未分化甲狀腺癌的潛在治療靶點。
