弗氏鏈霉菌雙拷貝串聯(lián)基因tylF、tylE和tylCV工程菌的構(gòu)建
2022-09-15 03:08:28陳亞蘭
中國(guó)獸藥雜志 2022年8期
陳亞蘭,牛 春,張 萍
(寧夏泰瑞制藥股份有限公司,銀川 750101)
泰樂(lè)菌素(Tylosin)是一種由弗氏鏈霉菌(Streptomycesfradiae)產(chǎn)生的大環(huán)內(nèi)酯類獸用抗生素,外觀為淡黃色粉末,微溶于水,易溶于有機(jī)溶劑,呈弱堿性[1],由美國(guó)研究者Hamill等[2]于1959年發(fā)現(xiàn),是國(guó)際上目前使用最廣泛的獸用抗生素之一[3-4 ]。泰樂(lè)菌素對(duì)革蘭陽(yáng)性菌、某些革蘭陰性球菌、支原體、分枝桿菌、螺旋體及原蟲(chóng)等有抑制作用,常用于防治由支原體感染引起的豬氣喘病,也可以治療豬痢疾、萎縮性鼻炎、關(guān)節(jié)炎、豬丹毒、鉤端螺旋體病等[5]。此外泰樂(lè)菌素還可添加到飼料中作為畜禽的抗菌促生長(zhǎng)添加劑,促進(jìn)畜、禽的生長(zhǎng)以及對(duì)飼料的利用率,被廣泛地應(yīng)用于畜禽生產(chǎn)及獸醫(yī)治療,所以國(guó)際國(guó)內(nèi)市場(chǎng)需求量極大[6-8]。目前,在大規(guī)模工業(yè)生產(chǎn)中,弗氏鏈霉菌是泰樂(lè)菌素的主要生產(chǎn)菌[9-11]。泰樂(lè)菌素主要由泰樂(lè)菌素A、泰樂(lè)菌素B、泰樂(lè)菌素C和泰樂(lè)菌素D四種組分構(gòu)成,其中泰樂(lè)菌素A為主要組分,且生物活性最強(qiáng)。工業(yè)生產(chǎn)上要求A組分含量≥85%,四種組分的總含量≥95%。
弗氏鏈霉菌泰樂(lè)菌素生物合成基因簇(tylgene cluster)從抗性基因tylB延伸至tlrC,約85 kb,含43個(gè)ORFs提供泰樂(lè)菌素合成所需完整的結(jié)構(gòu)基因,其中tylF編碼的大菌素-O-甲基轉(zhuǎn)移酶(Macrocin-O-Methyltransferase)催化泰樂(lè)菌素 C3 位羥基的甲基化反應(yīng)[12],催化泰樂(lè)菌素C轉(zhuǎn)化為泰樂(lè)菌素A,泰樂(lè)菌素 C3 位羥基的甲基化反應(yīng)是泰樂(lè)菌素生物合成的最后一步,也是限速反應(yīng)[13];tylE編碼去甲基大菌素-O-甲基轉(zhuǎn)移酶(Demethylmacrocin-O-methyltransferase)催化去甲基大菌素轉(zhuǎn)化為大菌素(Macrocin, tylosin C);tylCV編碼碳霉糖轉(zhuǎn)移酶(Mycarosyl transferase),負(fù)責(zé)將碳霉糖(Mycarose)轉(zhuǎn)移到去甲基拉克亭霉素(Demethyllactinomycin),合成去甲基大菌素(Demethylmacrocin)。
外源基因?qū)腈溍咕某S梅椒ㄓ蠵EG介導(dǎo)的原生質(zhì)體法、電轉(zhuǎn)化法和接合轉(zhuǎn)移法[14-15],相較于其他兩種方法,接合轉(zhuǎn)移法除了操作簡(jiǎn)單外,還可以避開(kāi)包外核酸酶,一定程度上能克服鏈霉菌對(duì)外源基因的限制[16],在鏈霉菌遺傳操作中廣泛應(yīng)用。鏈霉菌工程菌構(gòu)建中常用的載體為pSET152整合型表達(dá)載體,含阿泊拉霉素抗性基因,能通過(guò)噬菌體 φC31 的attB位點(diǎn)特異性整合到鏈霉菌染色體上[17-18 ],是鏈霉菌常用的質(zhì)粒。紅霉素抗性基因啟動(dòng)子(permE)是已知的鏈霉菌組成型強(qiáng)啟動(dòng)子,它常用于在鏈霉菌中提高目的基因(簇)的表達(dá)水平,以方便目的基因(簇)的表達(dá)產(chǎn)物或所生物合成化合物的檢測(cè),以及提高目標(biāo)化合物的生物合成水平等[19-20]。
1 材料與方法
1.1 菌株和質(zhì)粒 弗氏鏈霉菌、質(zhì)粒pSET152(含有紅霉素抗性基因啟動(dòng)子permE)均由寧夏泰瑞制藥股份有限公司保藏。
1.2 載體、限制性內(nèi)切酶和試劑 質(zhì)粒提取試劑盒、DNA回收試劑盒、全基因組提取試劑盒和各種酶均購(gòu)自TakaRa公司。
1.3 培養(yǎng)基和抗生素 大腸桿菌的培養(yǎng)基為L(zhǎng)B培養(yǎng)基;弗氏鏈霉菌固體培養(yǎng)基為高氏一號(hào)培養(yǎng)基;大腸桿菌-鏈霉菌接合轉(zhuǎn)移培養(yǎng)基為MS培養(yǎng)基;孢子預(yù)萌發(fā)培養(yǎng)基為2×YT;發(fā)酵培養(yǎng)基采用公司內(nèi)部培養(yǎng)基[21]。
LB中氨芐青霉素(Ampicillin, Amp)使用量為25 μg/mL,安普霉素(Apramycin, Apr)儲(chǔ)存濃度為50 μg/mL, 卡那霉素(Kanamycin, Kan)的使用量為25 μg/mL,萘啶酮酸(Naphthylpyruvic acid, Nap)的使用量為50 μg/mL。
1.4 目的基因的獲取 從GenBank下載permE、tylF、tylE和tylCV的基因序列,根據(jù)序列使用引物設(shè)計(jì)軟件 Primer Premier 5分別設(shè)計(jì)permE、tylF、tylE和tylCV的重疊延伸PCR引物(表1),其中permE的上游和tylCV的下游分別引入EcoRV和XbaI酶切位點(diǎn)。以弗氏鏈霉菌全基因組為模板,PCR擴(kuò)增獲得所需要的目的基因片段,純化后置于-20 ℃保存,備用。
表1 目的基因擴(kuò)增引物及酶切位點(diǎn)Fig 1 Primers and restriction sites for target gene amplification
1.5 重組片段permE-tylF-tylE-tylCV的獲得 用重疊延伸PCR法連接permE、tylF、tylE和tylCV基因,實(shí)驗(yàn)方法為:以permE和tylF為模板,用引物P-f/F-r擴(kuò)增獲得permE-tylF基因片段,純化后-20 ℃保藏;以permE-tylF和tylE為模板,用引物P-f/E-r擴(kuò)增獲得permE-tylF-tylE基因片段,純化后-20 ℃保藏;以permE-tylF-tylE為模板,用引物P-f/CV-r擴(kuò)增獲得permE-tylF-tylE-tylCV基因片段,純化后-20 ℃保藏。
1.6 構(gòu)建重組載體pSET152-permE-tylF-tylE-tylCV使用EcoRV和XbaI酶37 ℃,過(guò)夜酶切pSET152質(zhì)粒和permE-tylF-tylE-tylCV基因片段,回收酶切產(chǎn)物。按照目的基因與載體的摩爾比為7的比例加入酶切后的pSET152質(zhì)粒和permE-tylF-tylE-tylCV基因片段,16 ℃的溫度下在T4 DNA連接酶的作用下過(guò)夜反應(yīng)。將連接產(chǎn)物轉(zhuǎn)化至DH5a大腸桿菌后再轉(zhuǎn)入ET12567大腸桿菌,篩選陽(yáng)性克隆子。
1.7 ET12567(pUZ8002, pSET152)與弗氏鏈霉菌的接合轉(zhuǎn)移 將轉(zhuǎn)化子的過(guò)夜培養(yǎng)物轉(zhuǎn)接在LB中,在50 μg/mL的安普霉素條件下,37 ℃培養(yǎng)轉(zhuǎn)化子,到合適濃度(OD600在0.4~0.6)收集菌體;用等體積新鮮的LB洗滌菌體兩次洗掉抗生素,0.1倍體積的LB懸浮備用。刮取適量孢子研磨,將研磨液于漏斗中過(guò)濾,收集濾液,50 ℃熱激10 min,冷卻到室溫后加入等體積的2×YT孢子預(yù)萌發(fā)培養(yǎng)基;37 ℃、180 r/min搖床培養(yǎng)2~4 h,3000 r/min離心收集孢子,并懸浮于1 mL的培養(yǎng)液中,打散孢子備用;按照供受比為10∶1(或1∶1)將處理過(guò)的轉(zhuǎn)化子培養(yǎng)物和孢子懸液混合,混勻后涂布平板[22-23]。培養(yǎng)21 h后覆蓋萘啶酮酸(50 μg/mL)和安普霉素(50 μg/mL)。
1.8tylF、tylE和tylCV基因雙拷貝菌株泰樂(lè)菌素效價(jià)檢測(cè) 將適量弗氏鏈霉菌菌絲體與接合子的菌絲體接種到發(fā)酵培養(yǎng)基中,28 ℃、230 r/min連續(xù)培養(yǎng)7 d后,取硫酸鋁酸化后的上清液檢測(cè)泰樂(lè)菌素效價(jià),檢測(cè)方法參見(jiàn)文獻(xiàn)[24]。
1.9 接合子的分子水平驗(yàn)證 將出發(fā)菌與接合子的菌絲接種到含有Apr的LB培養(yǎng)基中培養(yǎng)至對(duì)數(shù)生長(zhǎng)后期,提取全基因組。分別以引物P-f/CV-r進(jìn)行PCR擴(kuò)增。
2 結(jié)果與分析
2.1permE、tylF、tylE和tylCV基因的擴(kuò)增 以弗氏鏈霉菌全基因組為模板,分別用引物P-f/P-r、F-f/F-r、E-f/E-r、CV-f/CV-r擴(kuò)增強(qiáng)啟動(dòng)子permE、tylF、tylE和tylCV基因,基因片段大小為270 bp(圖1a)、735 bp(圖1b)、1188 bp(圖1c)和1386 bp(圖1 d),將PCR產(chǎn)物純化后保藏至-20 ℃?zhèn)溆谩?/p>
2.2 重疊延伸PCR法連接permE、tylF、tylE和tylCV基因 通過(guò)重疊延伸PCR法獲得重組基因片段permE-tylF-tylE-tylCV,大小約為3500 bp,電泳結(jié)果如圖2所示,在3500 bp處出現(xiàn)單一、清晰的條帶,說(shuō)明重組片段連接正確,可用于后續(xù)實(shí)驗(yàn)。將該片段膠回收后于-20 ℃保藏備用。

圖2 重組基因片段permE-tylF-tylE-tylCV電泳圖Fig 2 Electrophoretic diagram of recombinant gene fragment permE-tylF-tylE-tylCV
2.3 供體菌的構(gòu)建 篩選到一株陽(yáng)性克隆子,提取質(zhì)粒進(jìn)行PCR驗(yàn)證(圖3)和酶切驗(yàn)證(圖4),在約3500 bp處擴(kuò)增出了目的條帶,使用EcoRV和XbaI也切出了約3500 bp大小的片段,結(jié)合空載體的酶切結(jié)果(圖5),初步說(shuō)明pSET152-permE-tylF-tylE-tylCV重組載體構(gòu)建成功,測(cè)序結(jié)果表明tylF、tylE和tylCV基因均與GenBank上發(fā)表的序列相同。至此,pSET152-permE-tylF-tylE-tylCV重組載體構(gòu)建成功,可用于后續(xù)實(shí)驗(yàn)。
圖3 pSET152-permE-tylF-tylE-tylCV重組載體PCR驗(yàn)證結(jié)果Fig 3 PCR verification results of recombinant vector pSET152-permE-tylF-tylE-tylCV
圖4 pSET152-permE-tylF-tylE-tylCV重組載體酶切驗(yàn)證結(jié)果Fig 4 Results of enzyme digestion verification of recombinant vector pSET152-permE-tylF-tylE-tylCV
圖5 pSET152載體酶切結(jié)果Fig 5 pSET152 vector digestion result
2.4 重組菌株的搖瓶發(fā)酵驗(yàn)證 從接合轉(zhuǎn)移培養(yǎng)皿中共挑選了20個(gè)單菌落,并將其于試管斜面培養(yǎng),培養(yǎng)12 d后僅有13個(gè)單菌落斜面長(zhǎng)勢(shì)較好,斜面孢子層較厚,呈白色。刮取適量帶有少量培養(yǎng)基的斜面孢子接種于發(fā)酵培養(yǎng)基進(jìn)行搖瓶發(fā)酵驗(yàn)證,培養(yǎng)7 d后測(cè)定效價(jià)(連續(xù)培養(yǎng)3批),結(jié)果如表2所示。其中3、6、7、8、9、10和13號(hào)連續(xù)三批的效價(jià)趨于穩(wěn)定,且高于出發(fā)菌株,分別高出約12%、9%、9%、18%、23%、29%和21%。用液相色譜儀分析3、6、7、8、9、10和13號(hào)單菌落的發(fā)酵液,結(jié)果如表3所示。通過(guò)表2可得,7、8、9、10、13號(hào)單菌落發(fā)酵液中泰樂(lè)菌素A的含量顯著(P≤0.05)高于出發(fā)菌,其中10號(hào)單菌落泰樂(lè)菌素A在10.778 min出峰(圖6),相對(duì)峰面積為59.07%,高出出發(fā)菌11.66%。同樣,10號(hào)單菌落的泰樂(lè)菌素C也顯著(P≤0.05)高于其它4株菌和出發(fā)菌,而泰樂(lè)菌素B和泰樂(lè)菌素D的變化不顯著,說(shuō)明增加tylE和tylCV基因的拷貝數(shù)可有效提高泰樂(lè)菌素C的含量,增加tylF基因的拷貝數(shù)可積累泰樂(lè)菌素A。
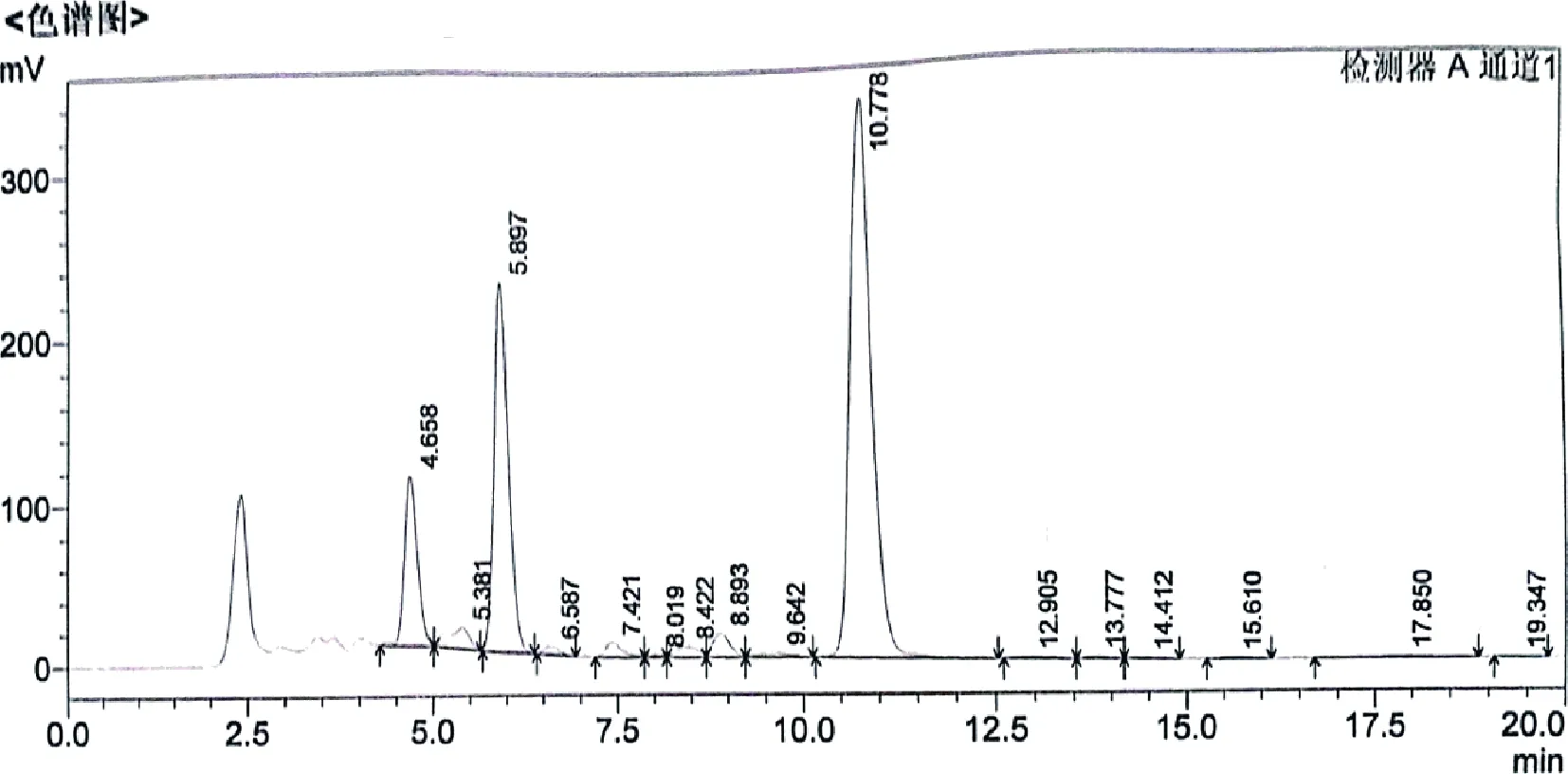
圖6 RTL21-10單菌落發(fā)酵液液相色譜峰圖Fig 6 Liquid chromatographic peak of fermentation broth of RTL21-10 single colony
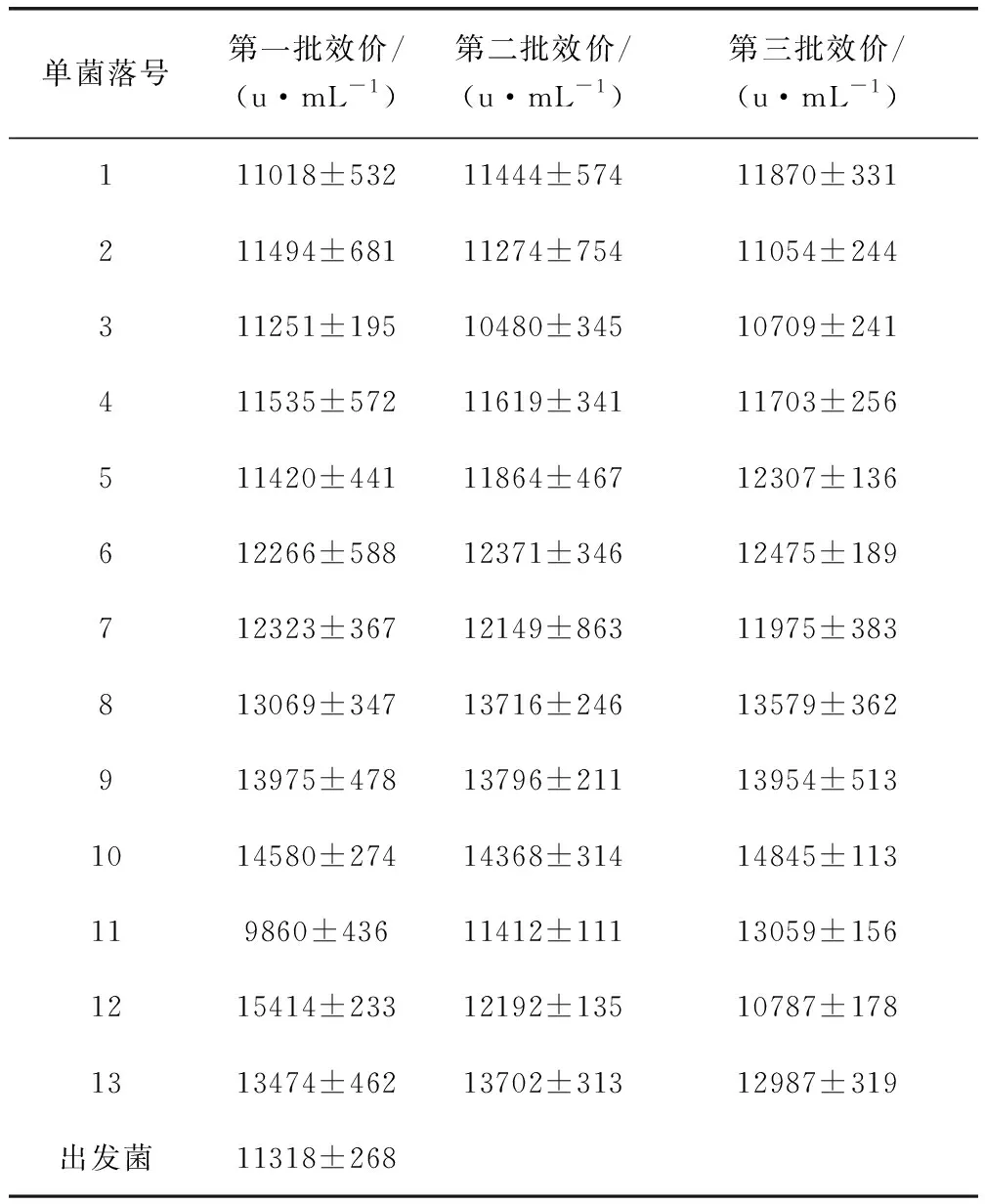
表2 連續(xù)3批發(fā)酵搖瓶效價(jià)測(cè)定情況Tab 2 Determination of fermentation titer for three consecutive batches of shaker
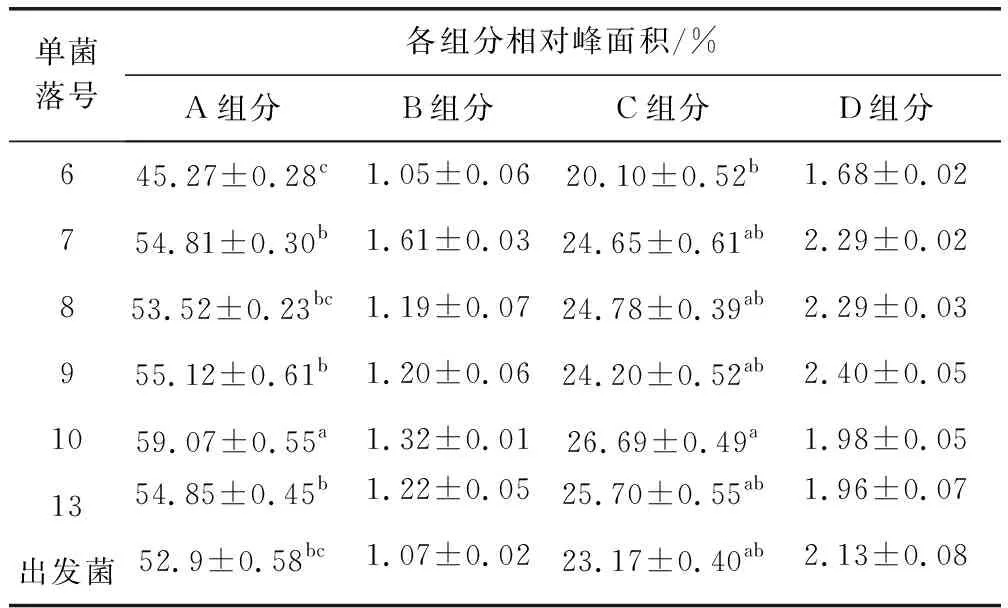
表3 各單菌落發(fā)酵液組分Tab 3 Components of fermentation broth of each single colony
2.5 工程菌株的PCR驗(yàn)證 提取 RTL21-10單菌落的全基因組,并以該全基因組為模板,用引物P-f/CV-r擴(kuò)增permE-tylF-tylE-tylCV片段,電泳結(jié)果如圖7所示,在3500 bp處有單一、清晰的條帶,說(shuō)明弗氏鏈霉菌tylF、tylE、tylCV雙拷貝工程菌構(gòu)建成功。
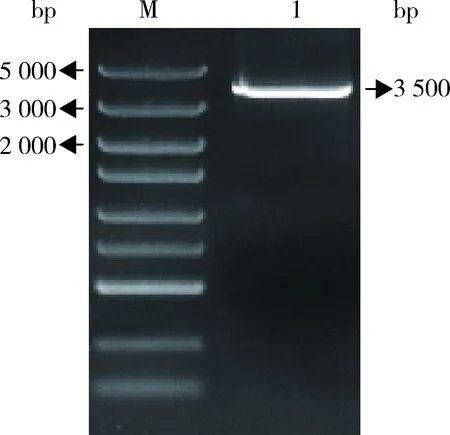
圖7 RTL21-10號(hào)單菌落PCR驗(yàn)證結(jié)果Fig 7 PCR verification results of RTL21-10 single colony
3 討 論
本研究成功構(gòu)建了一株雙拷貝tylE、tylF和tylCV基因的高產(chǎn)泰樂(lè)菌素工程菌株,以上三個(gè)基因由強(qiáng)啟動(dòng)子permE啟動(dòng)表達(dá),以確保發(fā)酵過(guò)程中工程菌的穩(wěn)定性,強(qiáng)化雙拷貝的tylF、tylE和tylCV基因的表達(dá)。采用以上策略,獲得一株弗氏鏈霉菌工程菌,單菌落編號(hào)為RTL21-10,效價(jià)比出發(fā)菌高出29%,利用液相色譜對(duì)RTL21-10單菌落的發(fā)酵液進(jìn)行分析,各組分均優(yōu)于對(duì)照菌株。由此可見(jiàn),增加泰樂(lè)菌素合成途徑中的相關(guān)基因的拷貝數(shù)可提高抗生素的產(chǎn)量,也可優(yōu)化組分。本研究成功構(gòu)建了高產(chǎn)泰樂(lè)菌素工程菌株,該方法為弗氏鏈霉菌的遺傳改造提供參考,具有重要的研究意義。
增加合成代謝途徑中關(guān)鍵酶基因的拷貝數(shù)是鏈霉菌代謝工程育種的有效手段[25],tylF基因編碼的大菌素-O-甲基轉(zhuǎn)移酶是限速酶,可催化泰樂(lè)菌素 C3 位羥基的甲基化反應(yīng)[26]。范亮等[27]構(gòu)建具有雙拷貝tylF基因的泰樂(lè)菌素基因工程菌,以解決泰樂(lè)菌素基因生物合成的限速環(huán)節(jié),重組菌株泰樂(lè)菌素的發(fā)酵單位較出發(fā)菌株提高32.7%。馬次郎[28]等人構(gòu)建了雙拷貝tylD、tylF、tylJ基因的泰樂(lè)菌素工程菌,泰樂(lè)菌素發(fā)酵搖瓶效價(jià)較出發(fā)菌株提高了28.1%,該菌株tylD、tylF、tylJ表達(dá)水平高于出發(fā)菌株,并且在進(jìn)入穩(wěn)定期后期表達(dá)量達(dá)到最大值。何翔宇[29]利用 Red/ET 同源重組技術(shù)構(gòu)建含有完整泰樂(lè)菌素生物合成基因簇的質(zhì)粒 pBAC-tyl,并將該質(zhì)粒在白色鏈霉菌 J1074(StreptomycesalbusJ1074)中進(jìn)行異源表達(dá),泰樂(lè)菌素的發(fā)酵產(chǎn)量達(dá)到 14.6 mg/L。此外,陳文青[30]利用紅霉素抗性基因啟動(dòng)子permE在弗氏鏈霉菌中異源表達(dá)vhb基因,在5L發(fā)酵罐條件下研究發(fā)現(xiàn),VHb蛋白的表達(dá)顯著促進(jìn)了泰樂(lè)菌素合成與菌體的生長(zhǎng),這種作用越是在限氧條件下,表現(xiàn)的越是明顯。但是,在工業(yè)生產(chǎn)過(guò)程中,隨著泰樂(lè)菌素產(chǎn)量的不斷提高,部分泰樂(lè)菌素被還原為雷諾霉素(Relomycin)—— 一種嚴(yán)重影響泰樂(lè)菌素產(chǎn)品質(zhì)量的副產(chǎn)物,Huang等[31]從弗氏鏈霉菌中純化了該酶,并對(duì)其性質(zhì)進(jìn)行了初步研究,確定了泰樂(lè)菌素還原酶隸屬于醛酮還原酶家族,朱慧等[32]的研究結(jié)果也證實(shí)了醛酮還原酶可催化還原泰樂(lè)菌素。
本研究成功構(gòu)建了高產(chǎn)泰樂(lè)菌素的工程菌株,但對(duì)tylE、tylF以及tylCV基因的表達(dá)情況還需進(jìn)一步研究,以更深入的了解弗氏鏈霉菌工程菌株的代謝途徑。同時(shí),在泰樂(lè)菌素的工業(yè)生產(chǎn)中,一方面可通過(guò)增加關(guān)鍵基因的拷貝數(shù)以提高泰樂(lè)菌素的產(chǎn)量,另一方面可通過(guò)抑制醛酮還原酶的活性以達(dá)到積累泰樂(lè)菌素的目的。
