凡納濱對蝦含酪氨酸酶結構域血藍蛋白的免疫功能
2022-09-28 01:40:16王子昂湯俊宇楊林蔚
廣東海洋大學學報 2022年5期
張 銳,王子昂,湯俊宇,楊林蔚
(1.廣東海洋大學食品科技學院/ 現代生化中心,廣東 湛江 524088;2.中山大學生命科學學院有害生物控制與資源利用國家重點實驗室,廣東 廣州 510275;3.南方海洋科學與工程廣東省實驗室(珠海),廣東 珠海 519000;4.廣東海洋大學深圳研究院,廣東 深圳 518108)
血藍蛋白是節肢動物和軟體動物血淋巴中的一種含銅呼吸蛋白,在氧運輸、能量貯存、滲透壓維持、蛻皮和表皮固化調節等方面發揮重要功能。研究發現,甲殼動物的血藍蛋白也有免疫活性,參與機體對多種病原的免疫防御[1-3]。在NCBI Genbank 數據庫中,每個甲殼類物種往往有多個序列有明顯差異的血藍蛋白編碼基因,表明其體內的血藍蛋白可能以多個基因表達產物混合物的形式出現,從而擁有結構和功能上的多樣性。因此,有必要從單一基因的角度去探討血藍蛋白的免疫功能。
凡納濱對蝦(Litopenaeus vannamei)是全球主要養殖蝦種之一,也是研究無脊椎動物免疫的重要對象。從NCBI 上公布的凡納濱對蝦基因組中共發現16個血藍蛋白基因,其蛋白結構域均擁有典型氨基端的Hemocyanin_N結構域和羧基端的Hemocyanin_C結構域,其中的一種血藍蛋白還含有酪氨酸酶結構域(Tyrosinase-domain),與其他血藍蛋白家族成員存在明顯差異,故命名為含酪氨酸酶結構域血藍蛋白(Tyrosinase-domain containing hemocyanin,TDCH)。本研究克隆該蛋白的基因tdch,分析其在對蝦抗副溶血弧菌免疫中的作用,為深入探索蝦類血藍蛋白功能以及病害防治提供基礎。
1 材料和方法
1.1 蝦和病原體
凡納濱對蝦(約10 g)購自深圳某對蝦養殖場,在約28 ℃富含氧氣的地下水(鹽度2.0) 中適應一周。正式實驗前,對5%的對蝦進行隨機抽樣,采用文獻[4-5]方法,通過PCR 檢測,確保沒有副溶血弧菌(Vibrio parahaemolyticus)和白斑綜合征病毒(White Spot Syndrome Virus,WSSV)污染。副溶血弧菌和白斑綜合征病毒按照文獻[6-7]方法制備。
1.2 凡納濱對蝦tdch基因片段的擴增及生物信息學
從凡納濱對蝦基因組和轉錄組數據庫獲得tdch基因序列[8],以凡納濱對蝦鰓組織cDNA 為模板,利用引物對TDCH-ORF-F/R(表1)擴增獲得tdch編碼區序列。PCR 反應體系(20 μL):Premix Taq 10 μL,ddH2O 7.5 μL,cDNA 模板0.5 μL,上下游引物各1 μL。PCR 產物經凝膠電泳檢測、目的條帶切膠回收,連接至pMD-19T,轉化至DH-5α 感受態細胞。挑取單菌落進行菌落PCR 檢測,陽性菌送至天一輝遠有限公司廣州測序部測序。獲得tdch序列后,運用SMART 網站(http://smart.embl-heidelberg.de/)分析tdch的結構域,運用AlphaFold 預測tdch的三維結 構[9]。用Clustal W 1.8 將tdch序列與Genbank 數據庫中凡納濱對蝦其他血藍蛋白序列進行比對。采用MEGA 5.0 軟件通過鄰接法(Neighbor-joining)進行進化樹分析,具體參數如文獻[10-11]。

表1 引物名稱及序列Table 1 Primers and their sequences
1.3 實時熒光定量PCR
取健康凡納濱對蝦,分別取肌肉、肝胰腺、鰓、胃、表皮、觸角、心臟、腸、幽門盲囊、眼柄和神經索,樣品剪碎至約2 mm,置于RNAlater(Invitrogen)中,血淋巴離心后用RNAlater 重懸。置4 ℃下12 h,放于-80 ℃冰箱保存,用于分析tdch組織分布。每組樣品中為15 尾蝦的組織混合物。每個組織至少取3組。
取凡納濱對蝦700 尾,隨機分為7 組,每尾蝦分別于第二腹肢基部位置用1 mL 的胰島注射器分別注射0.1 μg/μL 脂多糖(LPS)、0.1 μg/μL Poly (I:C)、2.0×103cfu/μL 金黃色葡萄球菌(Staphhylococcu saureus)、2.0×103cfu/μL 副溶血弧菌、2.0×103cfu/μL黑曲霉菌(Aspergillus niger)、2.0×104拷貝/μL WSSV 各50 μL,對照組注射同劑量磷酸鹽緩沖液(PBS,pH=7.2)[12]。在注射0、4、12、24、48、72、96 h時,分別隨機選取蝦6 尾,采集其鰓和血細胞,置于RNALater 存 儲。RNA 提 取、cDNA 合成和實時熒光定量PCR 安照文獻[13]的方法進行。根據tdch基因開放閱讀框的保守區,用Primer Premier 5.0 設計引物qRT-TDCH-F、qRT-TDCH-R,對照組EF-1α基因(Genbank 登錄號:GU136229)為管家基因,引物如表1。qRT-PCR 反應體系(10 μL):引物(10 μmol/L)各0.5 μL,Mix 5 μL,ddH2O 2.5 μL,cDNA 0.5 μL。反應條件:95 ℃4 min;95 ℃20 s、60 ℃30 s、72 ℃2 min,35 個循環;72 ℃10 min,95 ℃15 s。用2-ΔΔCt法計算相對表達量,用GraphPad Prism 7 軟件進行單因素方差分析。
1.4 RNA干擾
用T7 RiboMAX?Express RNAi System 試劑盒(Promega)依照文獻[14]及試劑盒說明書合成tdch基因特異的dsRNA,陰性對照為綠色熒光蛋白基因(GFP)的dsRNA,所用引物見表1。取體質健康、大小均一的凡納濱對蝦(約5 g)暫養后,將dsRNA用PBS 稀釋后,向對蝦的第二腹肢基部注射50 μL 1 μg/μL dsRNA。注射后48 h 隨機取蝦6 尾于RNAlater 中,-80 ℃保存,用于檢測dsRNA 對對蝦tdch的沉默效果。另外,在注射dsRNA 48 h 后,用1×106cfu/mL 的副溶血弧菌對凡納濱對蝦進行人工感染,每間隔2 h 統計對蝦死亡數量。
1.5 統計分析
采用SPSS 16.0 軟件進行統計學分析。計算3次檢測的平均值和標準差。采用t檢驗分析兩組數值間差異的顯著性。實驗組與對照組間存活率差異的顯著性則用Mantel-Cox log-rankχ2計算。
2 結果
2.1 凡納濱對蝦tdch結構
克隆得凡納濱對蝦tdch(GenBank 登錄號:ROT60816.1)的編碼區長1 656 bp,編碼551 個氨基酸,分子質量為63.37 ku,理論等電點為7.74。在1~17 氨基酸處含有一個信號肽,21~147 氨基酸處含有一個Hemocyanin_N 結構域,308~410 氨基酸處含有一個酪氨酸酶(Tyrosinase)結構域,413~522 氨基酸處含有一個Hemocyanin_C結構域(圖1、2)。

圖1 凡納濱對蝦tdch及其推導的氨基酸序列Fig.1 Tdch of Litopenaeus vannamei and its deduced amino acid sequence
2.2 凡納濱對蝦tdch與其他血藍蛋白的差異分析
將凡納濱對蝦tdch的mRNA 與其對應的DNA進行比對,發現tdch編碼區序列含有兩個外顯子及一個內含子。tdch氨基端含有一個Hemocyanin_N結構域,羧基端含有一個Hemocyanin_C 結構域,符合血藍蛋白家族的結構特征,與NCBI 數據庫中凡納濱對蝦其他血藍蛋白不同的是,其結構域中間含有典型的酪氨酸酶結構域(圖3)。構建的系統進化樹(1 000 倍bootstrap)表明,凡納濱對蝦tdch單獨聚為一支,在進化關系上遠離凡納濱對蝦其他血藍蛋白,提示tdch可能與以往研究的血藍蛋白不同,為新的血藍蛋白家族成員(圖4(A))。凡納濱對蝦tdch與其他物種血藍蛋白的系統進化樹顯示,凡納濱對蝦tdch與斑節對蝦(Penaeus monodon)、日本囊對蝦(Marsupenaeus japonicus)的血藍蛋白聚為一個分支,親緣關系較為接近,提示凡納濱對蝦tdch在進化較為保守(圖4(B))。

圖2 凡納濱對蝦tdch的結構Fig.2 Schematic diagram of tdch of Litopenaeus vannamei

圖3 凡納濱對蝦不同血藍蛋白的mRNA序列與對應的DNA序列比對結果及相應的結構Fig.3 Schematic diagram of different L.vannamei hemocyanins and the comparison results of their mRNA sequences and corresponding DNA sequences

圖4 凡納濱對蝦不同血藍蛋白(A)及甲殼動物血藍蛋白(B)的系統進化樹Fig.4 Phylogenetic trees of different hemocyanins in Litopenaeus vannamei(A)and TDCH in crustacean(B)
2.3 凡納濱對蝦TDCH的三維結構預測
經預測凡納濱對蝦TDCH 蛋白的三維結構簡單,主要以α螺旋結構為主,包含4處β折疊(圖5)。
2.4 凡納濱對蝦tdch基因組織表達分析
圖6 表明,tdch在凡納濱對蝦各組織中均有表達,在眼柄、肌肉、觸角和腸道中表達量較高,在肝胰腺、表皮、心臟和幽門盲囊中表達量較低。

圖6 凡納濱對蝦tdch的組織表達Fig.6 Expression of tdch in different tissues of Litopenaeus vannamei
2.5 凡納濱對蝦tdch時序表達
2.5.1 凡納濱對蝦免疫后tdch在血細胞中的表達變化 經6 種免疫原刺激后,凡納濱對蝦tdch在血細胞中表達量均顯著上升。其中,副溶血弧菌、金黃色葡萄球菌、白斑綜合征病毒和Poly (I:C) 刺激后,tdch在血細胞中4 h被極顯著激活(P<0.01),與對照組相比分別上調151.8、194.8、621.8 和1 254.9倍,并且在4~96 h 一直保持在高表達狀態。黑曲霉菌刺激后,tdch在血細胞中24 h 被顯著激活(P<0.01),與對照組相比上調2.8倍,并在48 h表達量驟升,與對照組相比上調322.6倍,并且在48~96 h一直保持在高表達狀態。LPS 刺激下,tdch在血細胞中24 及72 h 被顯著激活(P<0.01),與對照組相比分別上調3.5倍、1.8倍(圖7)。

圖7 不同免疫原感染后凡納濱對蝦tdch在血細胞中的表達Fig.7 Expression of tdch in hemocyte in Litopenaeus vannamei challenged by different immunogens
2.5.2 凡納濱對蝦免疫后tdch在鰓組織中的表達變化 圖8可見,經副溶血弧菌、金黃色葡萄球菌、黑曲霉菌、白斑綜合征病毒、LPS、Poly (I:C)或PBS 刺激后,凡納濱對蝦tdch在鰓組織中表達變化較小,在副溶血弧菌和黑曲霉菌刺激24 h時被顯著激活(P<0.01),與對照組相比分別上調2.1 倍和2 倍;在金黃色葡萄球菌刺激12 h時被顯著激活(P<0.01),與對照組相比上調1.7 倍,刺激4 h 及24~96 h 時表達量沒有影響;在白斑綜合征病毒刺激48 h 及96 h 時被顯著激活(P<0.01),與對照組相比分別上調2.4倍和1.9倍;在LPS刺激24 h及72 h時被顯著激活(P<0.01),與對照組相比分別上調1.9倍和1.9倍;Poly(I:C)刺激對tdch在鰓組織中的表達沒有影響。

圖8 不同免疫原感染后凡納濱對蝦tdch在鰓組織的表達Fig.8 Expression of tdch in gill of Litopenaeus vannamei challenged by different immunogens
2.6 凡納濱對蝦tdch在副溶血弧菌感染中的作用
圖9 表明,經RNAi 后,在血細胞中tdch被成功敲降,與對照組相比敲降效率達到70%。圖10 表明,注射dsRNA 48 h 后的對蝦經副溶血弧菌攻毒后,與GFP dsRNA 對照組相比,TDCH dsRNA 組的存活率顯著下降(Mantel-Cox log-rankχ2=8.159,P=0.043 <0.05),表明tdch的表達增強凡納濱對蝦對副溶血弧菌的抗性。
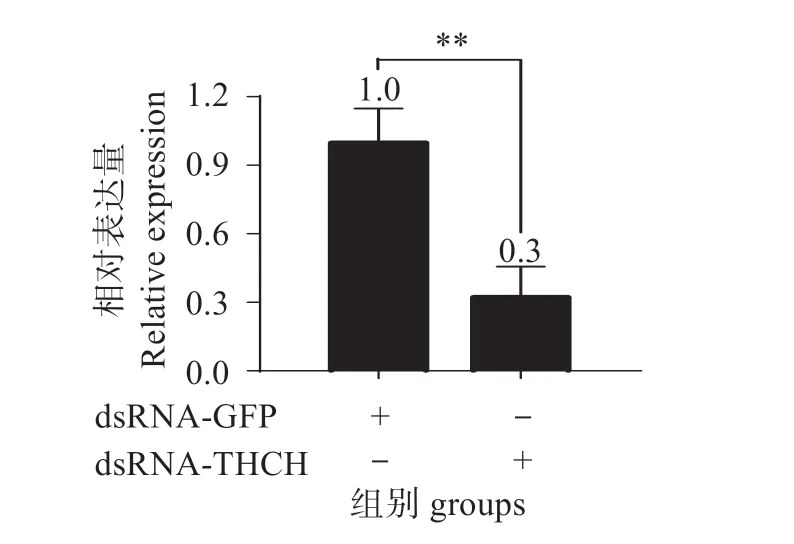
圖9 Tdch在血細胞中的敲降效率Fig.9 Knockdown efficiency of tdch in hemocyte

圖10 Tdch抑制副溶血弧菌感染Fig.10 Tdch inhibits Vibrio parahaemolyticus infection
3 討論
在凡納濱對蝦中,TDCH 蛋白氨基端的Hemocyanin_N 結構域和羧基端Hemocyanin_C 結構域與其他血藍蛋白類似[15],TDCH 還含有一個典型的酪氨酸酶結構域。酪氨酸酶是一種黑色素通路中的結合銅離子的酶,具有酚氧化酶活性,參與體液免疫反應、損傷修復等重要生理活動[16-17]。有研究表明,血藍蛋白可能是在大約55億年前由酚氧化物酶進化而來[18]。目前有研究證實血藍蛋白可能保留有酚氧化酶的活性,例如在軟體動物中,散蓋大蝸牛(Helix aspersa)的血藍蛋白在SDS 作用下能夠表現出酚氧化酶活性[19];也有報道稱凡納濱對蝦的血藍蛋白也具有酚氧化酶活性,并且還具有細胞凝集和細菌凝集活性[20-21],在外界病原菌刺激下能夠發揮類似酚氧化酶的活性參與免疫防御。血淋巴酚氧化酶活力包括血藍蛋白酚氧化酶活力和血細胞酚氧化酶原釋放的酚氧化酶活力,當機體受到外界病原刺激下,在一定時間內血細胞釋放的酚氧化酶不足以支撐機體的需要,此時血藍蛋白會表現出酚氧化酶活性參與免疫反應增強甲殼動物免疫防御能力[22]。而本研究中發現TDCH含有一個典型的酪氨酸酶結構域,提示TDCH 可能是對蝦血藍蛋白混合物酚氧化酶活性的一個來源,這還需要進一步實驗證實。
利用AlphaFold 預測凡納濱對蝦TDCH 的三維結構,結果顯示該蛋白主要以α螺旋結構為主,與已知節肢動物血藍蛋白的結構組成一致[23]。在對蝦中,眼柄與一系列生理功能高度相關,包括卵巢成熟、蛻皮、葡萄糖穩態,這些都與對蝦的生長高度相關[24]。Tdch在對蝦的眼柄中含量最豐富(圖4),提示tdch可能是內分泌調節相關基因之一。此外,血細胞[25]、鰓[26]和肝胰腺[27]是對蝦中重要的免疫相關組織。本研究中,這些組織中含有豐富的tdch,提示tdch可能與凡納濱對蝦的先天免疫有關。在副溶血弧菌、金黃色葡萄球菌、黑曲霉菌、白斑綜合征病毒、LPS 和病毒模擬物Poly(I:C)的刺激下,tdch在對蝦血細胞中的轉錄水平有顯著和持久的升高(圖5)。考慮到合成的dsRNA 類似物Poly(I:C)是病毒感染過程中發生的一種分子模式,tdch在Poly(I:C)和白斑綜合征病毒刺激過程中上調,表明tdch可能對包括DNA 病毒和RNA 病毒在內的病毒感染做出反應。值得注意的是,在副溶血弧菌、金黃色葡萄球菌和白斑綜合征病毒感染后4 h,tdch在血細胞中均顯著增加,提示tdch可能在感染早期就對細菌和病毒感染產生了反應。而黑曲霉菌刺激24 h 后tdch在血細胞中的表達才顯著增加,提示tdch可能在真菌感染的早期反應不明顯,主要在中晚期起作用。tdch在鰓組織中的反應弱于血細胞中,這可能是由于tdch需要在特定的組織環境中起作用。存活率實驗表明,敲降tdch能提高副溶血弧菌的敏感性,提示tdch在機體對抗副溶血弧菌感染中發揮重要作用。有研究表明,凡納濱對蝦血藍蛋白HMC可被副溶血弧菌、金黃色葡萄球菌及白斑綜合征病毒激活,促進p38及c-Jun磷酸化,調控下游抗菌肽的表達,參與抗菌與抗病毒感染[28]。然而凡納濱對蝦tdch是通過何種途徑發揮抗弧菌作用還有待進一步研究。
4 結論
本研究表明,tdch在各組織中均有表達,在眼柄中表達量最高;副溶血弧菌、金黃色葡萄球菌、黑曲霉菌、白斑綜合征病毒、LPS及Poly(I:C)刺激均可引起凡納濱對蝦tdch在血細胞中表達量上調,敲降tdch可提高副溶血弧菌的敏感性,表明tdch在機體對抗副溶血弧菌感染中發揮重要作用。本研究可為甲殼動物血藍蛋白類群的免疫功能研究提供新思路。
