鹽度馴化對虹鱒幼魚鱗片再生過程MMP2和MMP9基因表達的影響
2022-09-28 01:40:32王劉永周啟苓楊云生楊二軍李政鑫
廣東海洋大學學報 2022年5期
關鍵詞:水平
王劉永,周啟苓,馬 騫,2,楊云生,楊二軍,陳 剛,2,李政鑫
(1.廣東海洋大學水產學院,廣東 湛江 524025;2.南方海洋科學與工程廣東省實驗室(湛江),廣東 湛江 524025)
基質金屬酶蛋白(Matrix metalloproteinases,MMPs)屬于Metzincin 蛋白超家族[1],參與調控生物體多種信號通路,對骨組織細胞外基質重塑,器官發育,細胞遷移、增殖和凋亡,先天免疫和創傷修復等各項生命活動均有重要調節作用,在骨再生過程中發揮重要作用。MMP2(Gelatinase A)和MMP9(Gelatinase B)不僅參與調控骨基質的降解,還可介導成骨細胞的遷移及破骨細胞的活化、遷移和貼附等過程,是骨重建和骨吸收的關鍵調控因子[2-3]。MMP2 和MMP9 可促進骨髓間充質干細胞分化為成骨細胞[4];缺乏兩種酶則抑制由破骨細胞介導的骨吸收過程,從而影響骨重建進程[5]。MMP2 和MMP9 在金瑪麗魚(Poecilia latipinna)尾鰭再生初期表達水平增加,提示MMPs 在尾鰭再生前的炎癥調節及再生早期細胞外基質重塑中發揮作用[6]。Ando 等[7]研究發現,表達MMP9 的細胞在斑馬魚(Danio rerio)鰭條再生過程中可作為成骨祖細胞(Osteoblast progenitor cells,OPCs)進一步分化為成骨細胞,表明MMP9參與調節魚類骨骼再生過程。
鱗片是魚類外骨骼的重要組成部分,也是魚類特有的皮膚衍生物,其細胞組成和骨化機制與內骨骼十分相似,有保護身體、提供外部支持等功能[8-9]。硬骨魚類鱗片再生能力極強,一旦去除,可在幾周內生長到與正常鱗片相似大小和厚度[10-11]。此外,鱗片直接暴露于水環境且更易于采集,因此可作為骨骼再生研究的優良載體。已有研究表明,MMP2和MMP9在斑馬魚鱗片再生過程的早期階段大量表達,并在再生完成時恢復至與正常鱗片中相同的表達水平[11]。由此推測,MMP2 和MMP9 參與調節魚類鱗片再生過程,二基因表達水平可在一定程度上反映鱗片再生進程。
魚類的生存環境復雜多變,鹽度、溫度變化等均會對魚類各項生命活動產生影響。目前,關于鹽度變化對魚類骨骼再生過程的影響少有報道,僅見Guerreiro 等[12]研究的歐洲舌齒鱸(Dicentrarchus labrax)再生鱗生長參數及鈣化水平在鹽度馴化過程中的變化。虹鱒(Oncorhynchus mykiss)屬鮭形目(Salmoniformes)鮭科(Salmonidae)太平洋鮭屬(大馬哈魚屬)(Oncorhynchus),是目前世界上養殖最為廣泛的冷水性經濟魚類之一[13],也是我國養殖產量最高的鮭鱒魚類[14]。虹鱒對鹽度的適應性較強,經鹽度馴化后可在海水中正常生長[15],但入海后的存活率、生長速率等與鹽度馴化方式密切相關[16]。本研究檢測MMP2 和MMP9 基因在虹鱒幼魚鱗片再生過程中表達水平的變化,探討MMPs 基因在鱗片再生過程中的作用,進一步分析鹽度馴化對鱗片再生過程的影響,為揭示魚類骨骼再生的分子調節機制提供基礎資料,為鮭鱒魚類的養殖生產實踐提供參考。
1 材料方法
1.1 樣品采集
虹鱒幼魚體長(16.33±1.30)cm,體質量(52.21± 7.24)g,采集于山東省濰坊市臨朐縣淡水虹鱒養殖基地。實驗前將虹鱒置于直徑1.6 m,高0.6 m 的PE 桶中,水深約0.3 m,流水暫養3 d(鹽度3,水溫14~16 ℃),光周期條件為12 h:12 h,每天8:00、16:00投喂配合飼料(總投喂量約為魚體質量的3%)。
設置鹽度馴化組(Salinity acclimation,SA)和對照組(Control group,CG),每組設置3個平行。實驗前刮除虹鱒幼魚左側(背鰭后基部至尾柄處,長約2 cm,寬約1 cm)鱗片。實驗用海水晶調節鹽度,用鹽度計測定鹽度。鹽度馴化組以4/d 的速率將鹽度由3 升至27,對照組鹽度為3。分別在馴化1、3、7、14、21 d 時隨機取幼魚3 尾,解剖,分別采集魚體左側帶皮再生鱗(Regenerated scale,RS)及右側對應位置的帶皮正常鱗片(Ontogenic scale,OS),立即置于液氮中速凍,轉移至-80 ℃保存備用。
1.2 MMP2和MMP9基因的系統進化分析
基于基因MMP2(GenBank 登錄號:ON249089)和MMP9(GenBank 登錄號:ON249090)的序列信息,利用EditSeq 軟件預測虹鱒MMP2 和MMP9 基因的開放閱讀框,并將其翻譯成對應的氨基酸序列,用DNAMAN軟件與NCBI其他魚類及高等脊椎動物的MMP2 和MMP9 氨基酸序列進行多重序列比對,利用MEGA 6.0 軟件以鄰接法(Neighbour-Joining,NJ)構建系統進化樹,針對進化樹各分支結點進行1000次重復抽樣檢驗。
1.3 引物設計
基于虹鱒MMP2 和MMP9 的cDNA 序列,利用Primer Premier 5.0 軟件設計基因特異性引物MMP2-F/R、MMP9-F/R,內參引物18S-F/R(表1),用于實時熒光定量PCR(qRT-PCR)檢測。
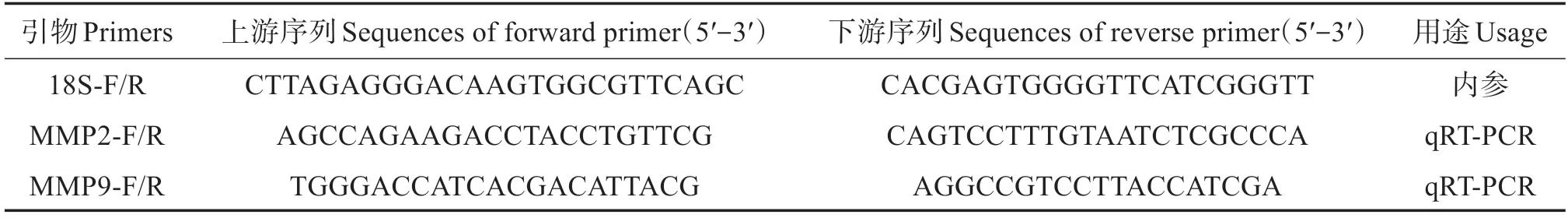
表1 所用引物序列Table 1 Primer sequences
1.4 總RNA提取及cDNA第一鏈合成
按照Trizol(Invitrogen)法提取虹鱒幼魚鱗片總RNA。通過瓊脂糖凝膠電泳和Nanodrop 2 000核酸蛋白測定儀(Thermo scientific)分別檢測總RNA 的完整性及濃度,根據EasyScript?Fist-Strand cDNA Synthesis SuperMix(TransGen)說明書,將1 μg 總RNA 反轉錄合成第1 鏈cDNA,保存于-20 ℃冰箱待用。
1.5 實時熒光定量PCR檢測
分別用引物MMP2-F/R 及MMP9-F/R,以18S rRNA 基因為內參,檢測鹽度馴化組及對照組正常鱗和再生鱗中MMP2 和MMP9 基因的相對表達量。qRT-PCR 根據TransStart Top Green qPCR SuperMix試劑盒(TransGen)說明操作,在LightCycler 96 熒光定量PCR 儀(Roche)上進行,各樣品設置3 個重復。所有引物均經過擴增效率檢測(E>90%;R2>0.990),qRT-PCR產物經測序驗證。
根據qRT-PCR 測得的Ct值,采用2-△△Ct法分別計算兩個實驗組各時間點再生鱗及正常鱗中MMP2和MMP9 基因的相對表達量,以再生鱗與正常鱗中MMPs 基因相對表達量比值表示再生鱗MMPs 基因表達水平。所得數據均以平均值±標準差(n=3)表示,使用SPSS 19.0 中的單因素方差分析(one-way ANOVA)法及Duncan's 多重比較,分析各基因表達水平在鱗片再生過程中不同時間點的差異水平,P<0.05時顯著差異,若P<0.01則差異極顯著。
2 結果與分析
2.1 MMP2和MMP9基因的系統進化分析
虹鱒MMP2和MMP9基因的開放閱讀框分別為1 968、2 028 bp,分別編碼655和676個氨基酸(圖1)。虹鱒MMP2 和MMP9 與已報道的其他硬骨魚類及高等脊椎動物相應MMPs氨基酸序列的系統進化樹中(圖2),虹鱒MMP2 和MMP9 均首先與鮭科的大西洋鮭(Salmo salar)聚為一支,再與鮭形目白斑狗魚(Esox lucius)的聚為一支;鮭形目MMP2 和MMP9分支均首先與鱸形目、鳉形目、鰈形目及頜針魚目種類聚為一支,再與鯉形目、鲇形目及電鰻目種類聚類;在硬骨魚類MMP2 和MMP9 分支中,骨舌魚目的美麗硬仆舌骨魚(Scleropages formosus)MMPs均單獨形成一分支。

圖1 虹鱒MMP2和MMP9基因編碼的氨基酸序列Fig.1 Deduced amino acid sequences of MMP2 and MMP9 in Oncorhynchus mykiss
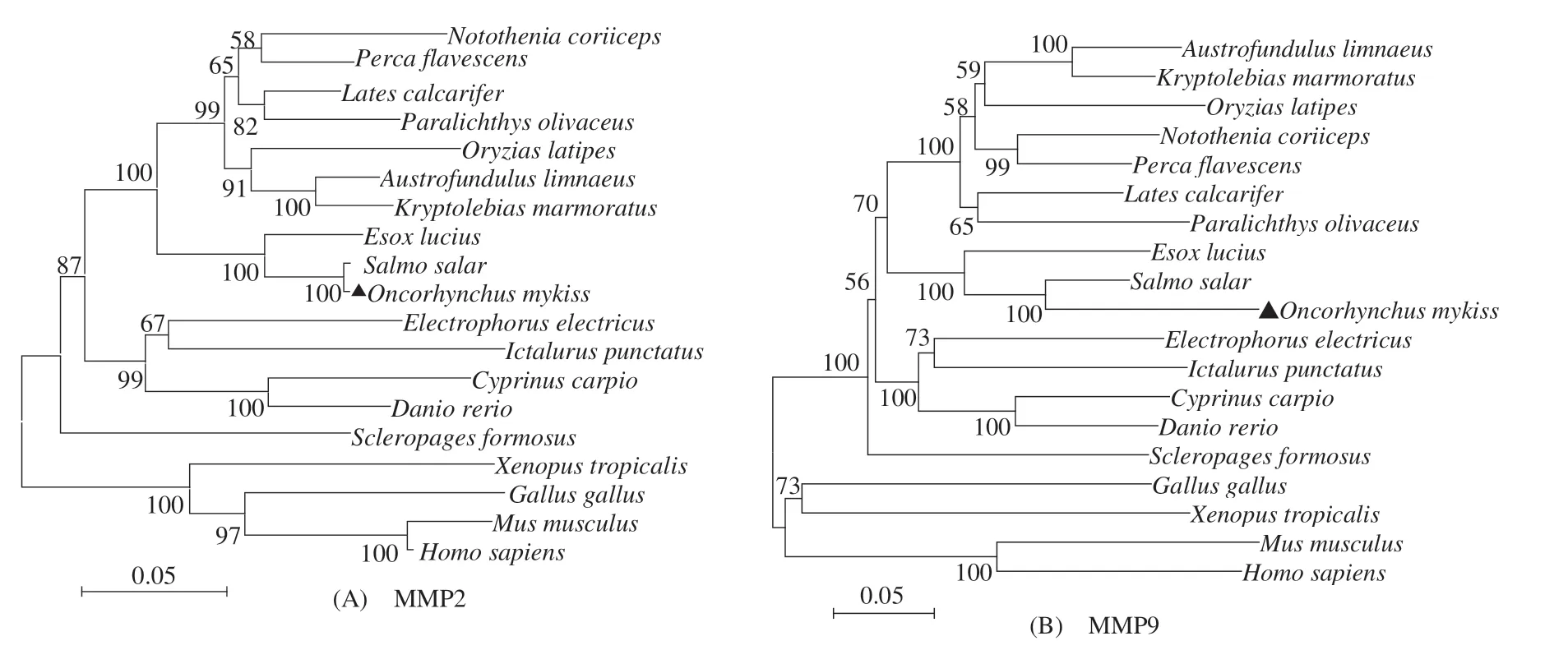
圖2 基于MMP2和MMP9氨基酸序列構建的NJ系統進化樹Fig.2 Phylogenetic trees of MMP2 and MMP9 amino acid sequences based on Neighbor-Joining method
2.2 MMP2 和MMP9 基因在鱗片再生過程中的表達水平
如圖3所示,在虹鱒幼魚鱗片再生過程中,對照組MMP2 表達水平呈先升后降趨勢:7 d 時最高(約為1 d 的8 倍),21 d 時依然顯著高于1 d;鹽度馴化組中再生鱗片MMP2 基因表達水平變化趨勢與對照組相似,總體呈先升后降趨勢:3 d 時最高(約為1 d 的2.2 倍),7 d 時顯著低于1 d,14 d 時最低(為1 d的17%)。對照組MMP9 表達水平在3 d 時顯著降低后,在7 d 時升至最大值,14 d 后始終維持在較低水平;在鹽度馴化組中,MMP9表達水平總體呈降低趨勢,1、3 d時較高,7、14 d時極顯著降至不足1 d的1%。鹽度馴化組及對照組再生鱗片的MMP9 基因在21 d時表達水平均有少許回升。
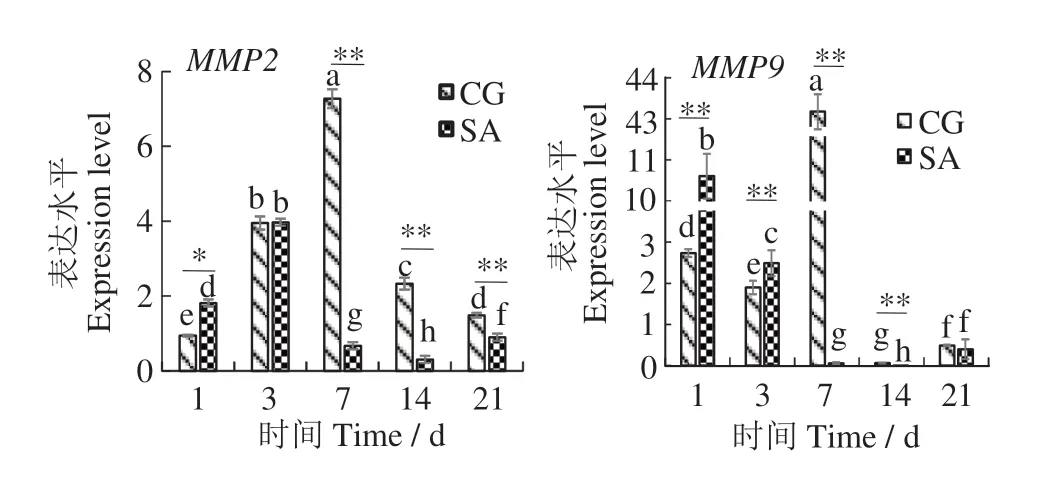
圖3 虹鱒幼魚鱗片再生過程中基因MMP2、MMP9的表達水平Fig.3 expression levels of MMP2 and MMP9 mRNA during scale regeneration in young Oncorhynchus mykiss
3 討論
基于19 個脊椎動物MMP2 和MMP9 氨基酸序列構建的系統進化樹顯示,MMP2 和MMP9 的系統進化樹結構較為相似;由于魚類MMPs 與哺乳類、鳥類及兩棲類的氨基酸同源性較低,導致進化樹在結構上主要分為硬骨魚類和高等脊椎動物兩大分支;硬骨魚類分支中,虹鱒MMP2 和MMP9 均首先與大西洋鮭聚成一支,與傳統形態分類結果一致。
BMP4可誘導軟骨細胞、成骨細胞的分化,在骨骼形成過程中有重要作用[17]。在鱖(Siniperca chuatsi)頜骨重塑初期,由間充質細胞分化的成骨細胞數量顯著增加,后期骨化趨于完善時成骨細胞數量減少;在這一過程中BMP4 表達水平相應地呈現先升后降趨勢[18]。由此可見,與成骨細胞分化、增殖等過程密切相關基因的表達水平可間接反映骨組織中成骨細胞的分化程度。本研究中,MMP2 和MMP9的表達水平在對照組虹鱒幼魚鱗片再生過程中均呈先升后降趨勢,表明兩個基因在鱗片再生過程中可能有調節作用。
Filanti等[2]發現,MMP2是成骨細胞分泌的MMPs中數量最高的酶,MMP2 在大鼠(Rattus norvegicus)成熟成骨細胞中顯著表達。MMP2在正常骨組織中表達量通常較低;但在骨修復早期階段會高表達,而后隨著骨修復完成表達量逐漸降低[19]。由此推測,MMP2 表達水平變化趨勢可作為鱗片再生過程中成骨細胞分化、增殖的標志特征之一,反映魚類骨骼再生的進程。MMP9 是破骨細胞分泌的主要蛋白之一,可能在激活破骨細胞從而啟動骨吸收的過程中發揮作用[20]。在斑馬魚鱗片再生的早期階段,破骨細胞與成骨細胞同時被活化,而在骨基質重塑后期二者細胞活性受到抑制[11];MMP9 表達水平變化可能與骨骼前體細胞的成熟和分化,以及成骨、破骨細胞活性等有關[21]。本研究中,MMP9在虹鱒幼魚鱗片再生過程早期表達水平較高,驗證了MMP9 對骨骼再生的調節作用,在鱗片再生過程中MMP9表達水平變化趨勢可能與破骨細胞的分化增殖及細胞活性的發揮有關。
本研究表明,在鹽度馴化初期(1~3 d),再生鱗MMP2 表達水平顯著高于或接近于對照組,隨著鹽度馴化的進行,馴化組MMP2 基因表達水平極顯著降低。此外,鹽度馴化組MMP9 表達水平呈顯著下降趨勢,表明鹽度馴化可能抑制了破骨細胞活性。已有研究表明,鹽度馴化對歐洲舌齒鱸再生鱗中成骨細胞標志性酶——堿性磷酸酶(alkaline phosphatase,ALP)的活性具有抑制作用,由此推測高鹽水體可能抑制成骨細胞分化,減緩骨礦化速度,從而抑制鱗片再生[12]。由此可見,鹽度馴化對虹鱒幼魚鱗片再生過程可能產生一定抑制作用。然而,鹽度馴化組MMP2和MMP9基因的表達水平在21 d均顯著回升至與對照組相近的水平,表明在鹽度馴化后期魚體已逐漸適應鹽度變化,再生過程不再受到抑制。
4 結論
1)MMP2和MMP9的表達水平在對照組虹鱒幼魚鱗片再生過程中均呈現先升高后降低的趨勢,兩個基因表達水平的變化可分別作為鱗片再生過程中成骨細胞與破骨細胞分化增殖的標志特征之一。
2)鹽度馴化抑制了鱗片再生過程中MMP2 和MMP9 表達水平,魚體骨代謝水平在鹽度適應過程中會發生變化,但在后期MMPs 基因表達水平恢復至與對照組相近水平,表明本研究采用4/d 的鹽度馴化模式較為合理,可為鮭鱒魚類養殖生產實踐提供重要參考。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
