墨吉明對蝦高血糖激素基因FmCHH-I的分子鑒定和表達特征
2022-09-28 01:40:20雷易果陳兆明
廣東海洋大學學報 2022年5期
關鍵詞:物種
雷易果,陳兆明,王 偉
(廣東海洋大學水產學院,廣東 湛江 524088)
在甲殼動物中,高血糖激素(Crustacean Hyperglycemic Hormone,CHH)是一類由眼柄中的X 器官-竇腺-復合體合成并分泌的神經肽類激素[1]。CHH 家族神經肽可分為I 型和II 型兩大類[2],CHH-I型神經肽有二硫鍵結構,信號肽與成熟肽之間由一段前導肽(CHH precursor-related peptide,CPRP)連接,而CHH-II 型神經肽結構中的信號肽直接與成熟肽相連。CHH-I 型神經肽僅包括經典的CHH 類激素,主要參與調節血淋巴中的葡萄糖水平;CHHII 型神經肽種類多樣,主要包括蛻皮抑制激素(Molt-inhibiting hormone,MIH)、大顎器官抑制激素(Mandibular organ-inhibiting hormone,MOIH)以及性腺抑制激素(Gonad-inhibiting hormone,GIH)[3]。CHH 家族神經肽在甲殼動物的脂質代謝、蛻皮、應激反應、性腺發育等多個生理過程中均發揮重要調節作用[4-5]。
我國是對蝦養殖大國,凡納濱對蝦(Litopenaeus vannamei)是人工養殖的主要蝦種,但其種質資源和親蝦嚴重依賴于進口,當前的對蝦行業還存在養殖環境惡化、種質退化、病害頻發等問題[6],這些都制約著對蝦養殖產業的可持續發展。因此,開發凡納濱對蝦的替代或補充物種十分必要。墨吉明對蝦(Fenneropenaeus merguiensis)是淺海的暖水性蝦類,在我國東南沿海均有分布,若其人工養殖技術得以突破,可在一定程度上緩解目前存在的對蝦養殖問題[7]。在墨吉明對蝦基礎生物學方面已有一系列研究,史黎黎等[8]研究了墨吉明對蝦C 型溶菌酶基因的克隆及表達,周婷婷等[9]研究了胰島素樣促雄性腺激素基因(FmIAG)的分子特征,楊世平等[10]研究了丁香酚對墨吉明對蝦的麻醉效果,為其人工繁殖方法的開發提供了參考。但是,目前墨吉明對蝦的大規模人工養殖技術還有待開發和完善,其基礎生物學知識還比較匱乏。有關對蝦CHH 家族神經肽的種類和功能,研究者已進行了一些探索,但目前對于墨吉明對蝦的CHH 家族基因信息了解的還比較有限,眼柄中的X器官對CHH家族基因的調控作用以及CHH 家族神經肽在墨吉明對蝦性腺發育中的功能演化仍有待于闡明。本研究利用分子生物學技術和生物信息學分析,獲得了墨吉明對蝦CHH-I基因(FmCHH-I)cDNA 序列,探究FmCHH-I的組織表達情況以及卵巢發育不同時期FmCHH-I的表達規律,分析生產上常用的單側眼柄切除法對FmCHH-I表達的影響,為探明FmCHH-I的功能奠定基礎,并為甲殼動物CHH神經肽的相關研究提供參考。
1 材料與方法
1.1 材料
實驗用墨吉明對蝦購自廣東省湛江市霞山區水產品批發市場,在室內循環水養殖系統中進行暫養,系統由供氣系統、給回水系統、生物濾池等組成,調節室溫為26~28 ℃,每天定時定點定量投喂3次飼料,對水體進行連續充氣并定期換水,維持良好水質,觀測對蝦的活動狀態是否異常。挑選活力較好的對蝦[體質量(30.5±0.8)g,體長(13.5±1.2)cm]進行后續實驗。
1.2 實驗試劑
TransZol Up Plus RNA Kit(RNA 提取試劑盒)、TransScript One-Step gDNA Removal 反轉錄試劑盒購自TransGenBiotech 公司,Primer ScriptTM1st Strand cDNA Synthesis Kit 逆轉錄試劑盒購自日本TaKaRa公司,SYBR PrimeScript RT-PCR Kit購自SangonBiotech公司,SMARTerTMRACE cDNA Amplification Kit 購自美國Clontech 公司,M5 HiPer Ptopo-TAVector、DH5α 大腸桿菌感受態細胞菌種購自全式金生物試劑公司。
1.3 總RNA 提取與FmCHH-I 基因cDNA 序列全長獲取
使用RNA 提取試劑盒,用Trizol 方法提取墨吉明對蝦總RNA。后通過反轉錄試劑盒將其反轉錄為cDNA。
根據本課題組前期獲取的墨吉明對蝦轉錄組序列信息(GenBank 序列號ON646230)設計特異性引物(表1),以墨吉明對蝦總RNA逆轉錄產物cDNA為模板,克隆出FmCHH-I基因的中間序列。采用Primer 5.0 軟件設計3′RACE、5′RACE 特異性引物(表1),利用RACE 技術擴增FmCHH-I基因的5′-UTR 和3′-UTR,從而得到FmCHH-I的cDNA 全長。PCR 產物通過瓊脂糖凝膠電泳檢測,對目的片段進行純化回收。通過連接、轉化后,按照實驗操作,提取出菌液后,用此菌液作為模板,選用通用引物M13(-47)和M13(-48),進行菌液PCR 擴增,通過瓊脂糖凝膠電泳檢測反應產物。分裝200 μL 菌液,送廣州艾基生物技術有限公司進行測序。
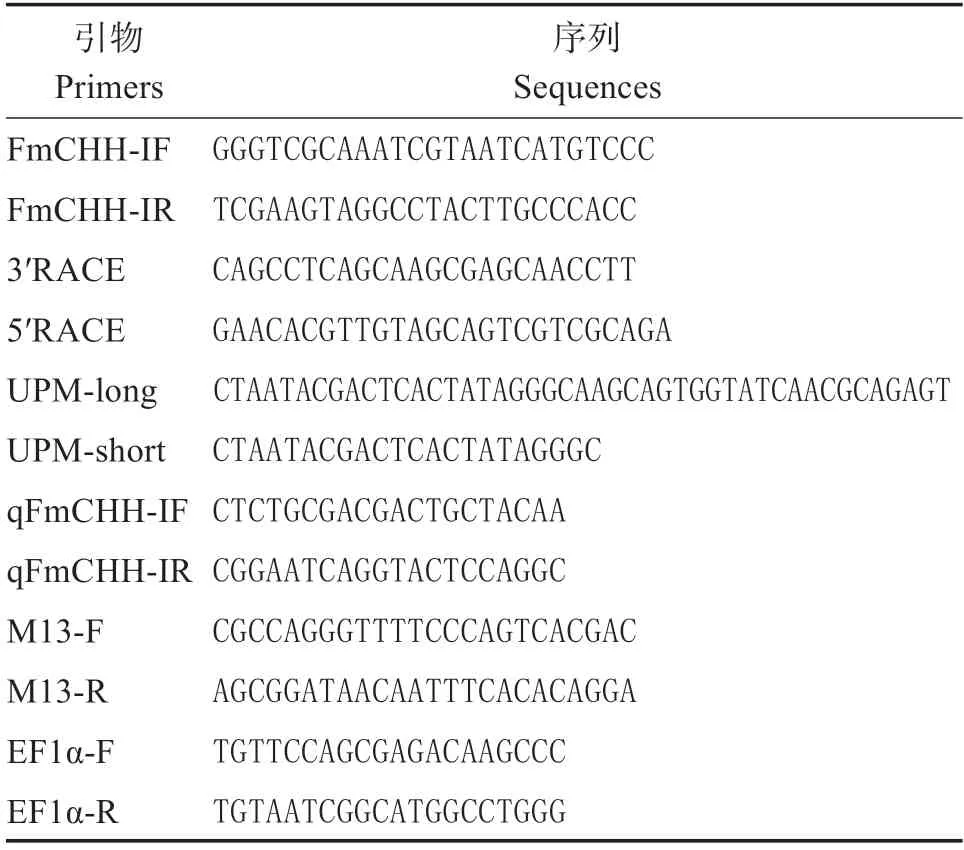
表1 PCR引物Table 1 PCR primers
1.4 生物信息學分析
通過NCBI 數據庫中BIASTX 在線程序(http://blast.Ncbi.nlm.nih.gov/Blast.cgi) 對墨吉明對蝦FmCHH-I基因進行同源性分析,利用SignalP 4.1 Server(www.cbs.dtu.dk/services/SignalP/)預測該氨基酸序列的信號肽以及具體位置;利用在線軟件Expasy(http://web.expasy.org/compute_pi/) 預 測FmCHH-I 蛋白分子質量和等電點;使用Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)比對墨吉明對蝦與其他物種CHH基因的氨基酸序列;用MEGA 7.0鄰接(NJ)法構建系統進化樹。
1.5 FmCHH-I基因的組織表達
取健康墨吉明對蝦的腦、眼柄、鰓、肝胰腺、腸、肌肉、卵巢等7種組織,進行RNA提取以及cDNA轉錄(方法見1.3)。用半定量RT-PCR方法檢測FmCHH-I在墨吉明對蝦不同組織中的轉錄情況。以EF1α 為內參引物,RT-PCR 擴增條件為95 ℃3 min;95 ℃30 s,58 ℃30 s,72 ℃30 s,35個循環;72 ℃10 min。
1.6 不同性腺發育時期FmCHH-I的表達
參考文獻[14]的方法,根據卵巢的外觀形態和性腺發育指數(GSI)區分墨吉明對蝦卵巢的不同發育階段。選擇處于卵巢成熟期與未發育的雌性墨吉明對蝦各3 尾,取其眼柄組織,進行RNA 提取并反轉錄為cDNA。以反轉錄的cDNA 為模板,設計定量PCR 引物qFm-CHH-IF/ qFm-CHH-IR,進行實時熒光定量PCR 檢測,EF1α 為內參引物(表1),目的基因和內參基因各做3個平行擴增。反應條件為95 ℃5 min;95 ℃15 s,55 ℃60 s,75 ℃5 s,40個循環。用2-△△CT法比較FmCHH-I基因在不同發育時期的眼柄組織中的表達量。利用SPSS 軟件對數據進行單因素方差分析,用Duncan 方法進行多重比較,顯著性水平設為0.05。
1.7 單側眼柄切除對FmCHH-I基因表達的影響
隨機取處于性腺成熟期的雌性墨吉明對蝦3尾,用鑷燙法切除單側眼柄。眼柄組織RNA提取和基因表達量檢測方法見1.6。比較切除及未切除眼柄的雌性墨吉明對蝦眼柄組織的CHH-I表達量。
2 結果與分析
2.1 FmCHH-I基因cDNA序列及其結構
克隆得FmCHH-I基因的cDNA 全長599 bp,其中5′非翻譯區(UTR)長度為37 bp,3′-UTR 長度為193 bp,開放閱讀框(ORF)長為369 bp,編碼123 個氨基酸(圖1),其中在第1~20 位包含一個信號肽(圖2),隨后有一段由26個氨基酸組成的前導肽,末端的成熟肽具有74個氨基酸,無跨膜區。

圖1 墨吉明對蝦FmCHH-I基因的cDNA和氨基酸序列Fig.1 Amino acid and cDNA sequence of gene FmCHH-I in Fenneropenaeus merguiensis

圖2 墨吉明對蝦FmCHH-I蛋白信號肽預測Fig.2 Signal peptide prediction of FmCHH-I in Fenneropenaeus merguiensis
2.2 FmCHH-I基因的理化性質分析
經預測,FmCHH-I基因編碼的蛋白分子質量為13.5 ku,理論等電點為7.56。脂溶指數為93.44,不穩定系數為64.58,被劃分為不穩定蛋白。總平均親水性系數為0.157,最高親水位在第15 位為3.622,最高疏水位在第112 位為-1.789,即此蛋白為親水性蛋白(圖3)。
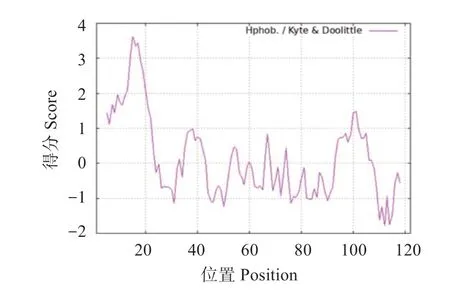
圖3 FmCHH-I蛋白疏水性Fig.3 Hydrophobicity for the FmCHH-I protein
2.3 FmCHH-I基因的同源性
FmCHH-I與GenBank 上部分物種的CHH-I同源序列比對結果見圖4。CHH-I 蛋白序列在不同甲殼動物之間較為保守,FmCHH-I與對蝦科物種相似性較高,與斑節對蝦(Penaeus monodon)的同源性最高,與熱液口蝦(Rimicaris kairei)的同源性最低,序列中6個半胱氨酸殘基在不同物種間高度保守(圖4),系統發育分析(圖5)表明,FmCHH-I 與斑節對蝦聚在一起,與對蝦科物種聚為一支,與斑節對蝦、凡納濱對蝦(Litopenaeus vannamei)、日本囊對蝦(Marsupenaeus japonicus)和刀額新對蝦(Metapenaeus ensis)等物種的CHH基因相似性較高。這些同源基因在所選物種氨基酸序列的后半部分均有相同的功能結構域(圖5)。

圖4 墨吉明對蝦FmCHH-I蛋白的多序列比對Fig.4 Multiple sequence alignment of the FmCHH-I protein of Fenneropenaeus merguiensis
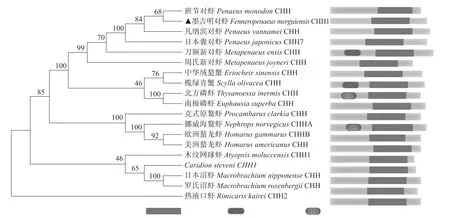
圖5 墨吉明對蝦FmCHH-I基因的系統進化分析和蛋白結構域預測Fig.5 Phylogenetic analysis of FmCHH-I gene and protein structural domain prediction
2.4 FmCHH-I基因的表達特征
圖6 表明,FmCHH-I基因在墨吉明對蝦眼柄中表達量最高,在肝胰腺、腸道中表達量次之,在其他組織中無明顯表達或表達量極低。
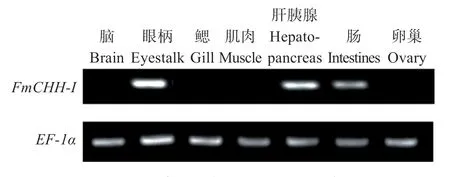
圖6 墨吉明對蝦CHH-I的組織表達Fig.6 Tissue distribution of FmCHH-I
圖7(a)顯示,FmCHH-I基因在雌性對蝦卵巢未發育期的表達量與性腺成熟期的表達量無顯著差異。處于性腺成熟期的墨吉明對蝦在切除眼柄后,FmCHH-I基因的表達量遠遠高于未切除眼柄對蝦的表達量(P<0.01)(圖7(b))。

圖7 性腺發育及切除眼柄對FmCHH-I表達量的影響Fig.7 Effect of gonad development and eyestalk abloation on FmCHH-I expression
3 討論
甲殼類高血糖激素(CHH)家族神經肽參與調控甲殼動物多種生理過程。許多學者研究了該超基因家族在甲殼動物中的功能。在日本沼蝦(Macrobrachium nipponense)[11]、斑節對蝦(Penaeus monodon)[12]、羅氏沼蝦(Macrobrachium rosenbergii)[13]等蝦蟹類中克隆出了CHH家族基因的序列,通過對比發現,不同物種間該基因結構存在差異,但總體保持著CHH家族基因功能結構域的保守性,如均有信號肽、成熟肽中均有6個半胱氨酸殘基等,這些特征也被證明是CHH基因超家族的典型特征[14]。
本研究首次在墨吉明對蝦中克隆到CHH 家族神經肽成員FmCHH-I基因的cDNA全長,為599 bp,其中5′-UTR 37 bp,3′-UTR 193 bp,ORF為369 bp,編碼123 個氨基酸,在第1~20 位包含一個信號肽,中間有一個由26個氨基酸組成的前導肽,隨后是一段74個氨基酸組成的成熟肽,無跨膜區,含有6個半胱氨酸殘基,成熟肽中第12位甘氨酸殘基缺失,且有二鹽基切割位點KR,為I型CHH神經肽的典型特征。
FmCHH-I 與CHH-II 蛋白氨基酸序列比對可見,CHH 神經肽中半胱氨酸的含量高,且序列高度保守。系統進化分析發現,CHH-I基因在對蝦科中極為保守。FmCHH-I與斑節對蝦、凡納濱對蝦、日本囊對蝦和刀額新對蝦的CHH 同源基因均有很高的相似性,且序列中的半胱氨酸高度保守,6 個半胱氨酸殘基形成3 個二硫鍵結構,起穩定肽鏈空間結構的作用。由此推斷,半胱氨酸在CHH神經肽的生物學功能保守性方面扮演著重要角色。
本研究的半定量RT-PCR 檢測結果顯示,FmCHH-I基因在墨吉明對蝦眼柄中表達量最高,在肝胰腺、腸道中也有表達,這與羅氏沼蝦[13]的結果一致。FmCHH-I在眼柄中高表達可能是因為,該基因主要由眼柄中的X 器官竇腺復合體合成和釋放,再通過血淋巴傳遞到其他組織而發揮功能[17]。但Liu 等在凡納濱對蝦的組織中發現,CHH基因在其心臟中的表達量最高,推測可能由物種間的差異造成[15]。Sedl-Meier等[16]研究發現,CHH-I基因能調節甲基法尼脂的分泌,并影響肝胰腺分泌消化酶,調控中腸釋放淀粉酶,由此解釋了在肝胰腺、腸道中有CHH基因表達的現象。
研究發現,在一些物種中CHH-I在雄性精巢中的表達量遠高于卵巢,由此推測CHH-I基因可能與雄性的發育密切相關[18]。對蝦養殖人員在生產上常常采取切除單側眼柄的方法促進卵巢成熟。本研究中,雌性對蝦性腺成熟期FmCHH-I基因的表達量與未發育期無顯著差異;處于性腺成熟期的墨吉明對蝦在切除眼柄后,FmCHH-I基因表達量遠高于未切除眼柄對蝦中的表達量,說明CHH-I基因不僅與對蝦體內血糖調節有關,還可能參與調控卵巢成熟過程。推測主要有兩方面原因。首先,在許多物種中CHH-I基因都有多種形式的可變剪切體或亞型,如美洲龍蝦(Homarus americanus)CHH-I基因就存在著CHH-A和CHH-B兩種形式,只有CHH-B對卵巢發育有促進作用[19]。Kleijn 等[20]在雌性美洲龍蝦生殖周期的研究中發現,X 器官/竇腺復合體中的CHH-A只在卵黃發生前期表達量較高,CHH-B則在卵黃發生前期與成熟期表達量較高。而本研究的FmCHH-I基因可能為CHH-I型基因中的一個亞型,因此在雌性對蝦性腺成熟期與未發育時期的表達量無顯著差異。
此外,對蝦性腺成熟期需要積累大量營養物質,這一時期對蝦所需要的營養物質遠高于生長期[21]。切除單側眼柄的目的是為了刺激對蝦性腺發育和繁殖,而移除儲存和分泌CHH 神經肽的X 器官/竇腺復合體后,對蝦體內的營養物質不能及時吸收和轉移,當體內能量不足時血糖含量會下降,此時需要大量的CHH-I 蛋白發揮作用,刺激機體產生更多的葡萄糖,從而滿足自身能量需求。
綜上,本研究首次在墨吉明對蝦中克隆得到FmCHH-I基因的cDNA 全長,多序列比對、系統進化和基因表達分析表明,FmCHH-I基因的功能較為保守,在對蝦的發育過程中可能起調節血糖以及參與性腺發育調控的作用,但此作用過程還需進一步的研究驗證。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
