一株富含多不飽和脂肪酸杜氏藻的鑒定和優化培養研究?
2022-09-30 03:19:56陳仁霞朱葆華曹子豪靳桂勇潘克厚
中國海洋大學學報(自然科學版) 2022年10期
陳仁霞,朱葆華,曹子豪,李 赟,靳桂勇,潘克厚,2??
(1.海水養殖教育部重點實驗室(中國海洋大學),山東 青島 266003; 2.青島海洋科學與技術試點國家實驗室 海洋漁業科學與食物產出過程功能實驗室,山東 青島 266237)
微藻作為水體環境中的初級生產者,可高效利用太陽能將CO2和H2O轉化為淀粉、油脂和多糖等有機物,目前已被廣泛開發應用于生物柴油和生物餌料領域[1]。然而,微藻在生產過程中存在生物量低和生產成本高的問題,且在不同環境條件下微藻的蛋白質、類胡蘿卜素和多不飽和脂肪酸等活性物質的含量不同,這些問題阻礙了微藻的規模化生產。當前,主要的解決途徑是篩選活性物質含量高的藻種并通過優化其培養條件獲得較大的生物量[2-3]。
多不飽和脂肪酸對機體有非常重要的生理調節功能[4]。例如二十碳五烯酸(EPA)能降低血液中甘油三酯和膽固醇的含量,從而加快體內飽和脂肪酸的代謝速率[5];二十二碳六烯酸(DHA)是促進大腦發育的重要物質,被認為具有誘導神經再生的功能[6];EPA與DHA是神經細胞膜的重要組成成分,主要從深海冷水魚魚油中獲取,資源稀缺。有研究表明α-亞麻酸可以在人體內通過去飽和酶和延伸酶的催化,轉化為EPA和DHA[7]。此外,α-亞麻酸還有增長智力、保護視力和延緩衰老等功效,具有重要的藥用及經濟價值;多不飽和脂肪酸中的亞油酸有降低血液膽固醇、降血壓和防治動脈粥樣硬化等功效,被稱為“血管清道夫”[8]。亞麻酸和亞油酸都是人體必需的脂肪酸,需從外界獲取,因此亟需尋找富含亞麻酸及亞油酸的原料。微藻具有富含多不飽和脂肪酸、生長快和可直接食用等優點,成為生產亞麻酸、亞油酸和EPA等多不飽和脂肪酸的重要原料。
杜氏藻(Dunaliella)屬于綠藻門(Chlorophyta)綠藻綱(Chlorophyceae)團藻目(Volvocales)杜氏藻科(Dunaliellaceae)。杜氏藻對鹽度有廣泛的適應性,在鹽度2~350的水域均能存活[9]。另外杜氏藻富含多糖、類胡蘿卜素和多不飽和脂肪酸等活性物質,具有無細胞壁、生長速度相對較快和抗逆性強等優勢被廣泛應用于規模化生產。因為杜氏藻的生物量易受溫度、鹽度和光照等培養條件的影響[10-12],所以可以通過優化培養條件獲得更高的生物量和更豐富的活性物質[13]。
本研究對一株分離自內蒙古哈馬太湖的藻株進行了形態學和分子生物學鑒定,確定該藻株為Dunaliellasp.,將其命名為HK128。并在此基礎上探討不同培養條件對HK128生長的影響,初步評估HK128生產多不飽和脂肪酸的潛力,進而為其開發利用提供理論依據。
1 材料與方法
1.1 藻種、培養條件及培養基
HK128是從內蒙古哈馬太湖(39°06′N,108°02′E)中分離的一株綠藻。哈馬太湖常年平均水深1.5 m,四季溫度12.5~26.0 ℃,pH在8.5以上。基于哈馬太湖水域微堿的特點,將分離的HK128接入Zarrouk培養基中進行為期7 d的預培養,而后收獲、轉接,進入正式培養。預培養在光暗比12 h∶12 h,溫度25 ℃,光照強度75 μmol/(m2·s)和轉速160 r/min的光照震蕩培養箱中進行。
1.2 HK128的鑒定
本實驗在Zeiss imager Z2微分干涉顯微鏡下對HK128進行形態學觀測,包括藻細胞形態和大小。隨后取指數生長期的藻液,用HP Plant DNA Kit(50)試劑盒法(OMEGA)提取HK128微藻基因組DNA,并設計2對引物18SF: 5’GCTGAAACTTAAAGGAATTGACGG3’, 18SR: 5’GAGAGCCAAGATATCCGTTGTTG3’;5.8SF: 5’TGCTGAAACTTAAAGGAATTGACG3’,5.8SR: 5’ GAGAGCCAAGATATCCGTTGTTG3’分別擴增18S rDNA和5.8S rDNA部分序列[14]。PCR產物送至上海生工生物工程有限公司進行測序。獲得的擴增序列經與GenBank中相關序列信息進行BLAST相似性檢索,選取相似性大于97%的序列,以光滑念珠菌(Candidaglabrata)、衣藻(Chlamydomonas)和小球藻(Chlorella)等作外類群,用Mega5.1鄰接法(Neighbor-joining)建立系統進化樹,重復1 000次計算bootstrap值,用于分離藻株的分子鑒定。
1.3 HK128培養條件優化
為優化HK128的培養條件,本研究分別比較了在不同的培養基、初始pH和鹽度等條件下(見表1)藻株HK128的生長情況。實驗流程為:先篩選出適宜HK128生長的培養基,然后進行溫度、初始pH、鹽度和光強的優化。

表1 實驗變量及變化范圍
1.4 分析方法
1.4.1 藻細胞密度及生物量測定 利用藻細胞的光密度(OD)和單位體積藻液中的藻細胞干質量(DCW)表征藻細胞的生長情況。藻細胞光密度(OD)采用分光光度計測定750 nm波長下的吸光值。用血球計數板法統計細胞密度,建立細胞密度與吸光度之間的標準曲線:
Y=1374.5X-159.76,R2=0.99。
藻細胞光密度OD750與細胞密度的相關性系數為0.99,顯示OD750與細胞數之間具有較好的相關性。
藻細胞干質量的測定方法如Lamers等[15]所述。將GF/C玻璃纖維素膜(直徑47 mm,孔徑0.45 μm)烘至恒重m0。取5 mL藻液,抽濾藻液至生物膜上,將生物膜放入65 ℃的烘箱,烘干至恒重m1。利用公式:藻細胞干質量(g/L)=(m1-m0)×200得出微藻的生物量。
1.4.2 藻細胞的葉綠素a、b及總類胡蘿卜素含量的測定 取第7天指數生長期的藻液5 mL測定葉綠素a、b以及總類胡蘿卜素,測定方法采用95%乙醇提取法[16]。按以下公式計算藻細胞的葉綠素含量:
CChlorophyll a(mg/L)=13.95×OD665-6.88×OD649;
CChlorophyll b(mg/L)=24.96×OD649-7.32×OD665;
2.05×Chlorophylla-114.8×Chlorophyllb)。
式中:C表示葉綠素含量;T表示總類胡蘿卜素含量;OD665、OD649和OD470分別表示藻液在665、649和470 nm下的光密度值。Chlorophylla表示葉綠素a,Chlorophyllb表示葉綠素b, Total carotenoids表示總類胡蘿卜素。
1.4.3 藻粉主要生化成分的測定 初始接種的OD750為0.2左右,收集第7天的藻液,離心(4 500 r/min,4 ℃,10 min),蒸餾水洗滌2次,再次離心后倒掉上清,收集藻泥并于真空冷凍干燥機中冷凍干燥24 h,冷凍干燥的藻粉放置于玻璃干燥皿中避光暫存。
1.4.3.1 可溶性總蛋白含量測定 采用BCA蛋白測定試劑盒法測定可溶性總蛋白[17]。稱取0.03 g冷凍干燥的藻粉,加入3 mL磷酸鹽緩沖液(0.1 mol/L,pH=7.4)于10 mL離心管中,冰浴條件下進行超聲細胞破碎。取離心后上清20和200 μL試劑混合(1 mL Cu試劑和50 mL BCA試劑混合),加入酶標板中,放恒溫培養箱于37 ℃孵育25 min。最后,用酶標儀562 nm波長測吸光值,通過牛血清蛋白標準曲線Y=0.847 7X+0.080 9,R2=0.99換算得到藻粉中可溶性蛋白的含量。
1.4.3.2 多糖含量測定 采用硫酸蒽酮法測定微藻中多糖含量[18]。稱取0.01 g冷凍干燥后的藻粉,加入6 mL磷酸緩沖溶液于10 mL離心管中,振蕩混勻。超聲破碎20 min,沸水浴30 min。取4 mL溶液,加0.2 mL硫酸鋅,沸水浴5 min;加0.4 mL亞鐵氰化鉀,振蕩混勻后離心。取1 mL上清于玻璃管中,加入硫酸蒽酮溶液4 mL,沸水浴10 min,冷水迅速冷卻至室溫15 min。測量620 nm處的吸光值,并通過制作的葡萄糖標準曲線Y=0.006 3X+0.030 1,R2=0.99,換算得到藻粉中多糖的含量。
1.4.3.3 總脂含量測定 采用質量量法[19],稱取0.04 g(m0)冷凍干燥的藻粉,加2 mL氯仿和1 mL甲醇于10 mL離心管中,渦旋振蕩混勻12 h。將離心管放置于有冰水混合物的玻璃燒杯中,超聲破碎20 min。離心(4 500 r/min)10 min,將上清液移至新的離心管中。重復3次上述過程,該過程合并所有上清至新的離心管中。上清液中加入1.2 mL 0.9%NaCl,渦旋震蕩后,靜置分層。吸取下層液體,轉移至恒重的玻璃管中(m1)用水浴將液體完全揮發,放置于65 ℃烘箱中烘干,冷卻至恒重稱重m2。總脂含量:
TTotallipidcontent=((m2-m1)/m0)×100%。
1.4.3.4 藻粉脂肪酸組成分析 采用Lamers等[15]氣相色譜分析方法,稱取0.01 g藻粉,加入1 mL(2 mol/L)KOH-CH3OH溶液,混勻,將玻璃管放入75 ℃水浴鍋中皂化,甲酯化30 min,冷卻至室溫。加入1.5 mL HCl-CH3OH(3 mol/L)溶液,混勻。75 ℃水浴鍋中酸化30 min,冷卻至室溫。加入2 mL蒸餾水,0.5 mL正己烷,混合均勻。3 500g離心10 min。吸取上層正己烷注入棕色進樣瓶中,冷凍避光保存,供色譜分析。將氣相色譜儀的條件設置如下:進樣口溫度250 ℃;內部程序升溫至150 ℃維持1 min,再以15 ℃/min升高到250 ℃保留10 min;載氣為N2和H2,流速分別為40和25 mL/min。采用面積歸一法,將樣品所有組分都顯示出峰值,實驗結果用被測組分的面積占總面積的百分比表示。
1.5 數據分析
計算3個重復實驗的平均值及標準差(n=3),進行統計分析。采用SPSS分析軟件進行單因素方差分析,p<0.01表示差異極顯著,p<0.05表示差異顯著,用Origin Pro 9.1進行作圖分析。
2 結果與討論
2.1 HK128的形態學觀察及分子生物學鑒定
用玻璃微吸管法將分離的藻株在Zarrouk培養基中培養,培養7 d后鏡檢。在48孔板中將目標藻株進行多次稀釋,并在固體Zarrouk培養基中涂布獲得單藻落。挑取單藻落轉入液體培養基,在光暗比12 h∶12 h、溫度25 ℃、光照強度75 μmol/(m2·s)條件下培養。在液體培養基中HK128藻細胞呈綠色的紡錘形(見圖1),長8~10 μm。有2條等長的頂生鞭毛,游動迅速。細胞側緣有一個紅色的眼點,中部有一個杯狀色素體。藻細胞沒有堅硬的細胞壁,周邊只有一層薄薄的生物膜包裹。根據藻細胞形態初步確定HK128為一株杜氏藻。
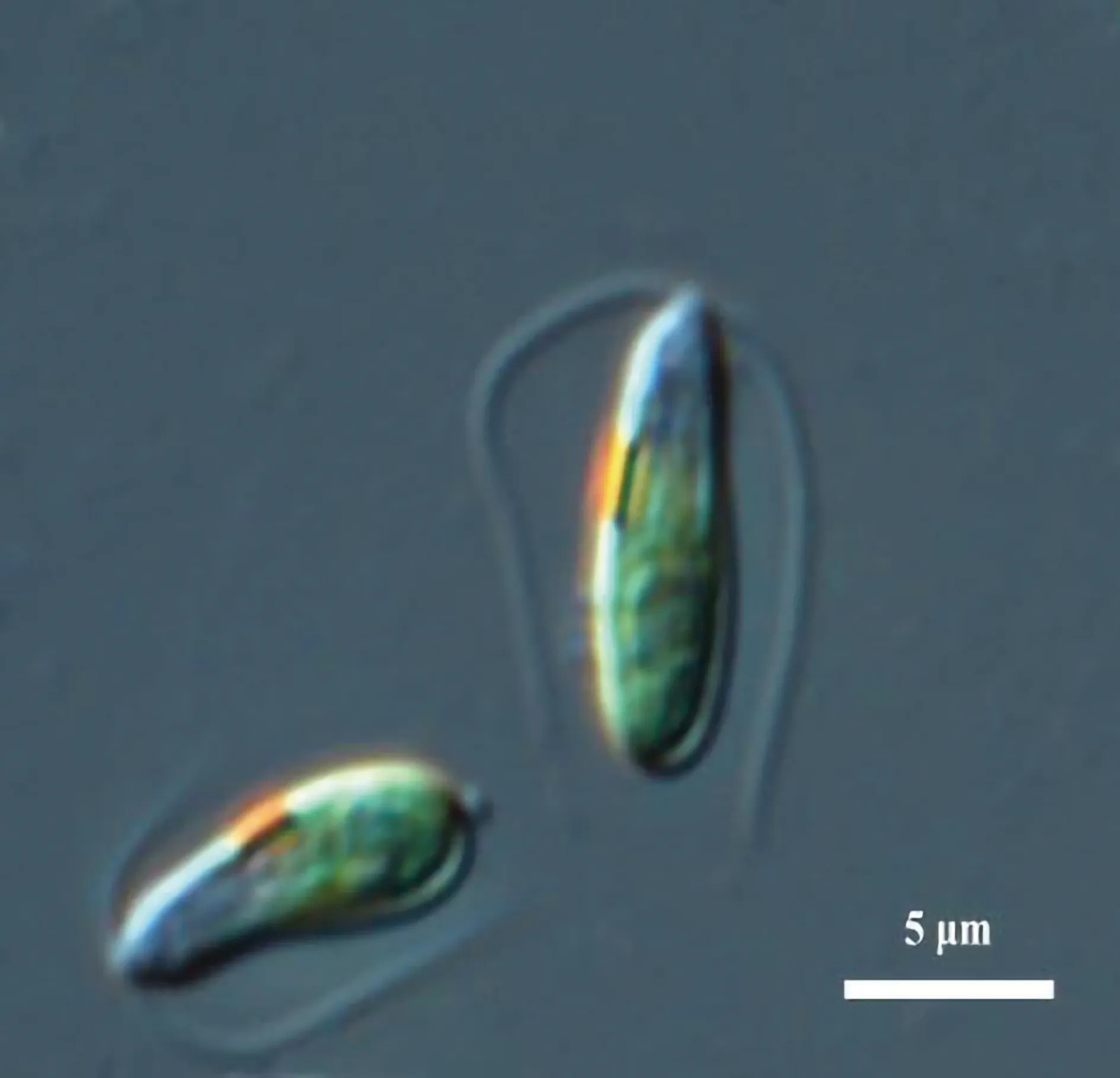
圖1 HK128藻細胞形態
本研究擴增得到的HK128 18S rDNA和5.8S rDNA序列長度分別為865和915 bp。結果顯示,獲得的與HK128序列相似性大于97%的序列共12條,均為杜氏藻屬的微藻。使用Mega5.1的鄰接法(Neighbor-joining),利用擴增序列及相似性較高的序列構建18S rDNA和5.8S rDNA序列系統進化樹(見圖2)。結果顯示,HK128的18S rDNA與杜氏藻屬中Dunaliellasalinastrain(KMMCC1064)在NJ樹上聚為一支,相似距離為95%。HK128的5.8S rDNA與杜氏藻屬中特氏杜氏藻Dunaliellatertiolectastrain(CCAP19/23)在NJ樹上聚為一支,相似距離達100%。核糖體5.8S rDNA具有很低的遺傳保守性,是進行生物種內及種間的理想分子標記[20]。但由于杜氏藻屬微藻在分類鑒定中仍存在著許多爭議,且分類體系尚未完全確立,所以本研究結果只能表明HK128藻株是Dunaliellasp.。
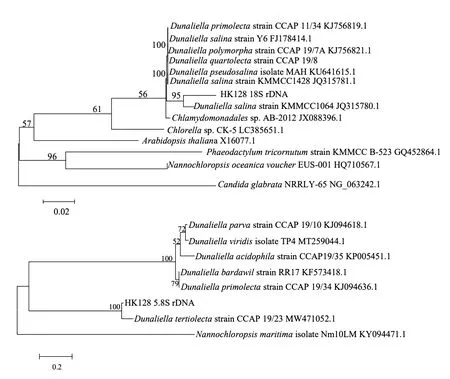
圖2 基于18S rDNA和5.8S rDNA序列信息構建的系統進化樹(Neighbor-joining法)
2.2 HK128的適宜培養基
HK128在3種培養基中培養7 d,結果顯示(見圖3),HK128在Zarrouk培養基(鹽度20,pH 9.5)中生長最優。在0~7 d保持持續增長的趨勢,培養基顏色逐漸加深至墨綠色。培養至第7天,HK128在Zarrouk培養基中的OD750是在f/2培養基(鹽度30,pH 8.11)中OD750的2.02倍,是在BG11培養基(鹽度5,pH 7.23)中OD750的5.43倍。說明HK128藻株更適合在Zarrouk培養基中進行培養。

((a)HK128在三種培養基中OD750的變化;(b)第7天HK128藻液顏色。(a)Changes of OD750 of HK128 in three media;(b)Day 7 HK128 algal liquid color.)
2.3 HK128培養條件優化
Santhanam等[21]和Dinesh Kumar等[22]的研究結果表明溫度、鹽度和光強等培養條件對微藻細胞的生長有顯著影響。有報道表明D.salina在30 ℃、D.viridis在37 ℃、D.tertiolecta在26 ℃生長最佳[23]。本研究中,HK128在20、25和30 ℃具有相同的生長趨勢(見圖4(a)),在第7天25 ℃時HK128藻細胞的OD750(1.5)顯著高于20 ℃(1.4)和30 ℃(1.1)時的OD750(p<0.01),說明HK128合適的培養溫度為25 ℃,與已報道的研究結果基本一致[22]。在戶外實際生產過程中,溫度波動大,HK128藻株有較寬的溫度適應范圍,這對于實際生產有重要意義。
pH從改變環境酸堿度和碳酸鹽平衡系統(CO2+H2OH2CO3H++HCO3-2H++CO32-)兩方面對微藻的生長產生影響[24-25]。本實驗利用NaHCO3和Na2CO3不同的添加比例來調節Zarrouk培養基的初始pH(9.3~11.5)。如圖4(b)所示,隨著培養液初始pH增大,藻細胞生長速度逐漸降低。在第7天,高pH組(pH=11.0和pH=11.5)的細胞密度顯著低于低pH組(pH=9.3和pH=9.5)的細胞密度(p<0.05),這表明HK128藻株適宜初始pH為9.3~9.5。Mclachlan等[26]發現杜氏藻能在pH為6.0~10.5的環境中正常生長,最適值為8.0~9.0,這與本研究的結果基本一致,說明HK128藻株對堿性環境具有較強的耐受性。
在溫度25 ℃、光暗比12 h∶12 h、初始pH=9.3,光強75 μmol/(m2·s)培養條件下,通過向Zarrouk培養基添加0~1.0 mol/L NaCl將培養液鹽度調至20~55,比較HK128的生長情況。結果顯示(見圖4(c)),HK128藻株在鹽度20~41的培養基中1~7 d持續增長,較低鹽度組(20和27)的細胞密度顯著高于較高鹽度組的細胞密度(34和41),而在高鹽度組(48和55)HK128藻細胞不能正常生長,可見HK128是一株適宜低鹽度培養的杜氏藻,與Cifuentes等[27]發現D.tertiolecta適宜在低鹽度生長的結果一致。HK128藻株適應低鹽度的原因可能是長期培養于低鹽度的培養基中。
光強對HK128生長的影響如圖4(d)所示,在光強為75~200 μmol/(m2·s)范圍內,HK128藻株的OD750隨著光照強度的增加相應提高,但當光強超過200 μmol/(m2·s),再提高光強并未明顯促進藻細胞的生長,但也未出現明顯的光抑制現象,說明HK128對高光強具有很強的適應性。

圖4 HK128的OD750在不同溫度(a)、初始pH(b)、鹽度(c)和光強(d)下的變化
在上述獲得的優化條件下,即溫度25 ℃,初始pH=9.3,鹽度20,光強200 μmol/(m2·s),在第7天HK128的生物量可達3.6 g/L,是Morowvat等[28]優化的D.salina生物量(1.015 g/L)的3.54倍。此外,隨著培養時間的延長,培養基的pH從初始的9.3增至第7天的10.23(見圖5),說明HK128具有較強的pH耐受特性。

圖5 在優化條件下HK128生物量(a)及pH(b)的變化
收集培養至第7天的藻粉,分析HK128的主要生化成分,結果如表2所示。其總脂含量為細胞干質量的24.83%、可溶性蛋白為細胞干質量的17.70%、多糖為細胞干質量的11.99%、葉綠素含量為26.00 mg/L、類胡蘿卜素含量為5.74 mg/L。Fazeli等[29]研究結果表明,D.tertiolectaDCCBC26在含有0.3 mol/L NaCl的培養基中葉綠素a含量為9.84 mg/L、類胡蘿卜素含量為3.75 mg/L。姜建國等[30]分析了5種杜氏藻的生化組成發現Dunaliellabardawil847、Dunaliellabioculata199、Dunaliellaprimolecta818、Dunaliellasalina435和Dunaliella558的總脂含量在11.7%~20.8%,相比上述已報道的杜氏藻藻株,本實驗培養的HK128的總脂含量(24.83%)具有顯著優勢。

表2 在優化條件下第7天HK128的主要生化成分含量
進一步分析HK128的脂肪酸組成(見表3),發現該藻株脂肪酸碳鏈長度主要集中在C4~C24之間。HK128的脂肪酸圖譜基本與Chen等[31]研究的結果一致。其中棕櫚酸(C16∶0)和α-亞麻酸(C18∶3n-3)是HK128的主要脂肪酸,相對含量分別為33.84%和32.48%,其次是相對含量為10.40%的順-11-二十碳烯酸(C20∶n-9)和相對含量為8.05%的亞油酸(C18∶2)。一般來說,棕櫚酸是最豐富的脂肪酸,在所有的微藻群中普遍存在。有研究表明,Dunaliellaprimolecta[32]和Phaeodactylumtricornutum[1]中棕櫚酸含量分別為21.8%和21.63%。

表3 在優化條件下第7天HK128的脂肪酸組成(占總脂肪酸的百分含量)
杜氏藻中富含亞麻酸和亞油酸,Xie等[33]在150 μmol/(m2·s)光強下培養的Rhodomonassalina含有12.90%的α-亞麻酸和16.47%的亞油酸。Evans等[34]發現在10 d的連續光照條件下,D.tertiolecta中α-亞麻酸占總脂肪酸含量的34.7%。本研究中,HK128中α-亞麻酸占總脂肪酸含量的32.48%,但其生物量(3.6 g/L)顯著高于已報道的Dunaliellatertio-lecta(0.46 g/L)和Dunaliellasp.(0.78 g/L)的生物量[35-36]。說明該藻株具有用于生產亞麻酸的優勢。α-亞麻酸是維系人腦進化的核心物質,是保持人體健康的必需脂肪酸,具有增長智力、保護視力和抑制癌癥轉移等功效[7]。目前亞麻酸主要存在于深海魚油和亞麻籽中,但是由于漁業資源日漸枯竭,亞麻籽生長環境苛刻、產量低,導致亞麻酸資源缺乏,亟需尋找新的亞麻酸生產原料。
3 結語
本研究分離的藻株HK128是一株喜低鹽度和高光強、耐高溫及高pH的Dunaliellasp.。在溫度為25 ℃,光暗周期為12 h∶12 h,初始pH=9.3,光強為200 μmol/(m2·s)條件下,在鹽度20的Zarrouk培養基上進行優化培養,在培養的第7天HK128的生物量為3.6 g/L,總脂含量可為細胞干質量的24.83%,其中多不飽和脂肪酸α-亞麻酸占總脂肪酸含量的32.48%。由以上結果可以看出,HK128富含α-亞麻酸、生長速度快、生物量高,可作為生產α-亞麻酸的理想候選藻株,具有很大的開發潛力。
