人類胎盤的葡萄糖轉運機制及GDM 對胎盤葡萄糖轉運的影響
2022-10-08 13:47:42何銀東柳汝明
昆明醫科大學學報 2022年9期
何銀東,張 峻,黃 樺,柳汝明,王 茜
(昆明醫科大學第一附屬醫院臨床藥學科,云南 昆明 650032)
葡萄糖是胎兒的主要能量來源,由于胎兒缺乏合成自身所需葡萄糖的能力,胎兒需要的所有葡萄糖均通過胎盤從母體獲取,因此胎盤在葡萄糖轉運至胎兒的過程中行駛非常重要的作用。葡萄糖從母體側到胎兒側的轉移主要通過胎盤上的葡萄糖轉運體(glucose transporters,GLUTs)完成。此外,胎盤自身對葡萄糖的代謝是調控葡萄糖從胎盤向胎兒轉運的重要因素。
1 正常胎盤上的葡萄糖轉運
1.1 正常胎盤上GLUTs 的分布及其對葡萄糖轉運的影響
胎盤上葡萄糖的轉運,首先由母體面的合胞體滋養層細胞,將母體側的葡萄糖轉運至胎盤中,合胞體滋養層細胞可分為面向母體的微絨毛膜(microvillous membranes,MVM)及面向子體側的基底膜(basal membrane,BM)。再由胎盤子體面的血管內皮細胞,將胎盤中的葡萄糖轉移到子體側。具體來說,葡萄糖可通過濃度擴散,或者分布在上述不同細胞中的各種葡萄糖轉運體(glucose transporters,GLUTs)完成葡萄糖轉運[1],見圖1(據Placenta,引自Day P E,2013)。其中,GLUTs在胎盤上葡萄糖轉運過程中行駛著非常重要的作用。
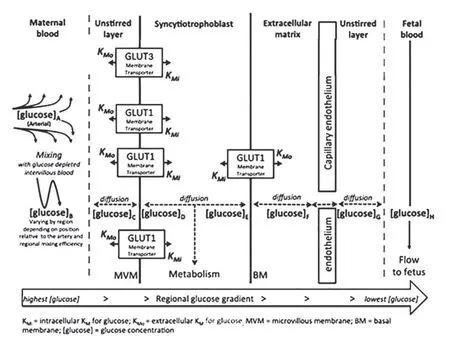
圖1 胎盤上的葡萄糖轉運簡圖[1]Fig.1 Transplacental glucose transport
人類胎盤共表達有GLUT1、3、4、8、9、10和12 七種亞型。目前的研究結果表明不同的GLUTs 在胎盤上的表達水平和分布位置不同,同時,一些GLUTs 的表達隨著妊娠期的變化而改變,提示不同GLUTs 在胎盤葡萄糖轉運過程中行使不同的功能。
GLUT1 在合胞體滋養層上的表達水平顯著高于胎盤其他部位,是胎盤上主要負責葡萄糖轉運的轉運體,GLUT1 也表達在胎盤內皮細胞。隨著妊娠的推移,GLUT1 在滋養層細胞MVM 上的表達逐漸增加,孕晚期達到最高水平[2-3]。GLUT 1在MVM 的表達量約是BM 的3~4 倍[4],折疊的MVM 展開后的膜面積是BM 的7~9 倍,這使得MVM 上的物質交換效率遠高于BM。因此有假設提出,BM 上的葡萄糖轉運是葡萄糖經合胞體滋養層轉運的限速步驟[5]。Vardhana 等[6]發現,隨著BeWo 細胞BM 上GLUTs 表達的減少,葡萄糖轉運也減少。
GLUT3 主要表達于合胞體滋養層的MVM 以及胎盤血管內皮細胞。隨著妊娠的推移,合胞體滋養層細胞上的GLUT3 的表達下降[7]。因此推測GLUT3 主要在孕早期發揮作用。GLUT3 與葡萄糖有非常高的親和力,這保證了細胞在能量物質缺乏時能快速有效獲得葡萄糖。在孕早期,子宮-胎盤血流系統不完全成熟,在低能量物質(如葡萄糖)的環境中,GLUT3 能促使大量葡萄糖快速從母體側轉運到胎兒側,滿足胎兒對葡萄糖的需求[8]。
GLUT4 表達在合胞體滋養層細胞的BM 上,MVM 上幾乎不表達,且孕晚期胎盤上的合胞體滋養層BM 上GLUT4 的表達顯著升高[9]。GLUT4 是胰島素依賴型葡萄糖轉運體。在孕早期,胰島素刺激GLUT4 的表達及其介導的葡萄糖攝取。James-Allan[9]及Ericsson 等[10]發現,胰島素能增加孕早期合胞體滋養層細胞上GLUT4 的表達和葡萄糖攝取。Hiden 等[11]研究發現孕早期胰島素受體(insulin receptor,IR)主要表達在合胞體滋養層細胞的MVM。Desoye G 等[12]的研究結果中,免疫組化結果提示:孕早期,IR 主要表達在合胞體滋養層細胞的MVM,而在孕晚期,主要表達在胎盤上的內皮細胞。最近,James-Allan 等[9]發現在MVM 和BM 上都有IR 表達,但在MVM 上的表達是BM 上的10 倍,隨著妊娠推進,MVM 和BM 上的IR 表達都增加。因此,有假設提出:胰島素可能通過MVM 上的IR 調控BM 上GLUT4 的表達及其介導的葡萄糖轉運。而且,在孕晚期,胰島素也可能影響經胎盤的葡萄糖攝取。
GLUT8 表達于足月胎盤上的合胞體滋養層和內皮細胞中。但GLUT8 對經胎盤的葡萄糖轉運的影響尚不清楚。
胎盤上的GLUT9 存在GLUT9α 及GLUT9β兩種亞型,前者主要表達在合胞體滋養層的BM以及內皮細胞中,后者主要表達在合胞體滋養層的MVM 上[13]。
胎盤組織中發現GLUT10 存在,但其在胎盤上的位置和功能都不明確。
GLUT12 主要表達于孕早期合胞體滋養層細胞的胞質中。在足月胎盤中,主要表達于絨毛血管平滑肌和絨毛間質細胞。隨著妊娠的推進,合胞體滋養層細胞上GLUT12 的表達降低[14]。在妊娠不同時期,GLUT12 的分布變化可能與胰島素受體的變化有關,可能參與孕早期合胞體滋養層細胞上的胰島素依賴的葡萄糖攝取。
1.2 胎盤上葡萄糖代謝對葡萄糖轉運的影響
除了GLUTs 的分布和活性會影響胎盤上的葡萄糖轉運外,胎盤上葡萄糖代謝、子宮-胎盤血流量也會影響胎盤上的葡萄糖轉運。Trond 等[15]發現約30%被胎盤自身消耗。而且,胎盤的葡萄糖消耗與胎盤的葡萄糖攝取呈正相關,與胎兒的葡萄糖消耗負相關。這表明胎盤自身對葡萄糖的消耗也是調節葡萄糖母親-胎兒轉移的重要手段之一,但目前對此方面的研究仍然較少。
2 妊娠期糖尿 病(gestational diabetes milletus,GDM)患者胎盤上葡萄糖轉運的改變
GDM 是指妊娠期間首次發生或發現的不同程度的葡萄糖耐受不良。全球發病率高達14.0%[16],我國GDM 發病率高達8.4%~18.9%,且呈逐年上升的趨勢[17]。GDM 會對子體產生不良影響。短期內,新生兒則發生巨大兒、低血糖和高膽紅素血癥風險增加[18];且在后期發生肥胖及2 型糖尿病等慢性代謝疾病的風險增加[19]。
研究發現,與正常孕婦胎盤相比,GDM 患者的胎盤重量、直徑、厚度等有所增加[20-21],胎盤上的纖維素樣壞死、脈絡膜血管病變、缺血性疾病和絨毛不成熟均增加[22]。因此GDM 患者的胎盤結構發生不同程度病變,可能會引起胎盤葡萄糖轉運的異常等功能改變,從而可能增加胎兒發生不良結局的風險。
2.1 GDM 患者胎盤上GLUTs 的變化
GDM 患者常表現出高血糖和高胰島素血癥,早期體外實驗探討了血糖和胰島素對胎盤合胞體滋養層細胞GLUT1 的表達水平的影響。例如,Hahn、Illsley 等[23-24]發現,當血糖值為0 mmol/L時,合胞體滋養層細胞上的GLUT1 的表達增加,葡萄糖攝取增加。而當血糖值為20~25 mmol/L時,GLUT1 的表達降低后,葡萄糖攝取減少。而當血糖濃度為1~12 mmol/L 時,GLUT1 的表達并未發生改變[24]。但該研究尚未探討血糖對GLUT1活性的影響。Basak 等[25]發現胰島素可促進孕早期BeWo 細胞株的GLUT1 mRNA 的表達和葡萄糖攝取。
一些研究報道了GDM 對胎盤上GLUT1 功能的影響,但結果不完全一致。Jansson 等[26]發現正常產婦與血糖控制良好GDM 患者的MVM 和BM 上GLUT1 的表達和葡萄糖攝取無顯著差異。然而,Gaither[27]和Yao 等[28-30]的研究發現GDM患者合胞體滋養層上GLUT1 的表達增加,且與子代出生體重呈正相關。GLUT1 表達的增加可能是GDM 患者中葡萄糖轉運過多和胎兒生長加速的原因之一。
Sciullo E 等[31]發現與健康產婦相比,GDM患者胎盤上GLUT3 的表達降低。但Kainulainen等[32]的免疫組化結果顯示,GDM 患者胎盤上GLUT3的表達無顯著變化。因此,目前關于GDM 患者胎盤上GLUT3 表達的影響尚無統一定論。
研究GDM 胎盤上GLUT4 表達的研究報道較少,結果也不盡一致。Stanirowski 等[2]發現,胰島素治療的GDM 胎盤滋養層細胞上的GLUT4 表達增加,但是Colomiere 等[28]發現GDM 胎盤上GLUT4 的表達水平降低,且胰島素治療會進一步降低GLUT4 的表達水平。GDM 胎盤上GLUT4 的表達水平的改變對葡萄糖轉運的影響尚無報道。
Stanirowski 等[2]的研究發現,胰島素治療的GDM 胎盤上GLUT9 的表達水平顯著高于正常胎盤。另外還有研究報道,GDM 患者胎盤BM 上的GLUT9α 表達水平明顯高于正常胎盤。
暫無關于GDM 影響胎盤上GLUT8 及GLUT12的表達水平的報道。
由目前的研究結果可知,GDM 可能會通過影響胎盤上GLUTs 的表達,從而改變胎盤上的葡萄糖轉運。但GDM 對不同GLUTs 的表達影響可能不完全相同,并且,飲食控制及胰島素治療的GDM 對GLUTs 的影響也有所差異。另一方面,大部分的研究結果均研究胎盤上GLUTs 的表達水平,針對其轉運活性的影響研究較少。
2.2 GDM 患者胎盤上葡萄糖代謝的變化及其對葡萄糖轉運的影響
一些研究討論了GDM 患者胎盤上葡萄糖代謝的改變對胎盤上葡萄糖轉運的影響。最近的一項研究發現,GDM 胎盤上的細胞滋養層細胞的糖原合成減少[33],但是Michelsen 等[15]的研究表明GDM 患者胎盤上的糖原合成與正常產婦無差別。
3 胎盤上調節葡萄糖轉運的分子機制及GDM 患者胎盤上分子機制的改變
已有一些研究探討了胎盤上AMP 激活的蛋白激酶(AMP kinase,AMPK)、哺乳動物雷帕霉素靶標(mammalian target of rapamycin,mTOR)信號通路和胰島素/胰島素樣生長因子1(insulin-like growth factor 1,IGF1)信號通路對胎盤上葡萄糖轉運影響。
AMPK 是人體重要的能量傳感因子,當細胞內低能量狀態時(AMP/ADP:ATP 的比例增加),AMPK 信號通路被激活,從而促進葡萄糖及脂肪酸等物質的產能代謝。AMPK 可以調節骨骼肌、肝臟等組織中GLUTs 轉運功能。一些研究也報道了AMPK 信號通路調節胎盤上葡萄糖轉運,以及GDM 患者胎盤上AMPK 活性的改變。Carey,E.A等[34]發現在胎盤滋養層細胞中,AMPK 敲除可抑制GLUT3 蛋白的表達及活性。肥胖的GDM 患者胎盤上的AMPK 的mRNA 表達水平下降[35-36]。Guo Yao 等[28]研究結果表明,GDM 胎盤上GLUT1表達增加,AMPK 及其下游ACC 的磷酸化水平降低,提示AMPK 信號通路被抑制。GDM 合并巨大兒患者胎盤上AMPK 及ACC 磷酸化水平與新生兒的體重呈負相關[37]。
mTOR 是絲氨酸/蘇氨酸蛋白激酶,其與不同的蛋白成分組合形成mTOR 復合物1(mammalian target of rapamycin complex,mTORC1)和mTORC2,mTORC1 是胎盤上感應葡萄糖轉運和代謝的重要靶點之一[38]。胎盤合胞體滋養層細胞上的mTOR信號通路受葡萄糖調節[39],葡萄糖可通過生成ATP 直接與mTOR 結合激活mTOR 信號通路[40]。mTOR 信號通路可能會影響胎盤上GLUTs 的表達水平。在JEG-3 滋養層細胞中抑制mTOR 的表達可降低GLUT3 的表達水平[41]。GDM 胎盤上mTOR信號通路被激活,mTORC1 的下游關鍵蛋白磷酸化水平顯著上調,且磷酸化水平變化與子代出生體重呈正相關[42]。
胰島素/ IGF1 是調節胎盤生長和胎兒發育的關鍵信號通路。IGF1 與IGFR1 受體或者IR 結合后,導致胰島素受體底物(insulin receptor substrate,IRS)以及磷脂酰肌醇3 激酶(phosphatidylinositol-3 kinase,PI3K)的激活,激活的PI3K,最終激活下游的AKT 和mTOR 信號通路,增加糖原合成和儲存。研究發現IGF1 可促進合胞體滋養層細胞上的葡萄糖攝取和GLUT1 的表達[43]。
4 小結
胎兒從母體有效攝取葡萄糖有賴于胎盤的正常結構和功能,胎盤主要通過多種GLUTs 將母體側葡萄糖轉運至子體側,此外胎盤上葡萄糖代謝、子宮-胎盤血流量也會對葡萄糖轉運產生影響。GDM 患者胎盤上的葡萄糖轉運與正常孕婦胎盤上的葡萄糖轉運存在差異。目前研究認為胎盤合胞體滋養層細胞上的GLUTs 是導致葡萄糖轉移發生改變的原因之一,不同的GLUTs 表達及活性的改變,如何對GDM 患者胎盤上葡萄糖轉運產生影響,有待進一步確認。此外GDM 胎盤細胞滋養層細胞的葡萄糖代謝活性改變也可能影響葡萄糖的轉運。影響胎盤上葡萄糖轉運的具體機制包括AMPK、mTOR 及胰島素/IGF1 信號通路。但具體分子機制的改變對胎盤上葡萄糖轉運以及對胎兒不良結局的影響,仍然有待進一步研究。
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中國生殖健康(2019年2期)2019-08-23 08:12:10
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51