基于線粒體COII基因序列的長江下游翹嘴鲌群體遺傳多樣性分析鲌
2022-10-11 12:52:30張桂寧方弟安薛向平毛成誠彭云鑫
南方農業學報 2022年7期
關鍵詞:研究
張桂寧,方弟安,2*,薛向平,毛成誠,彭云鑫
(1中國水產科學研究院淡水漁業研究中心/農業農村部淡水漁業和種質資源利用重點實驗室,江蘇無錫 214081;2上海海洋大學/水產科學國家級實驗教學示范中心,上海 201306;3南京農業大學,南京 210095)
0 引言
【研究意義】翹嘴鲌()俗稱大白魚、翹嘴白魚等,隸屬于鯉科(Cyprinidae)鲌亞科(Cultrinae),是東亞特有鯉科魚類中重要的一個類群(倪勇和伍漢霖,2006)。翹嘴鲌生活于水體上層,是一種廣溫性魚類,其肉質鮮嫩,生長迅速,營養價值高,是目前人工養殖的重要水生經濟魚類之一(何舜平等,2004)。近年來,由于水域環境遭到破壞,加之過度捕撈,致使我國野生的翹嘴鲌資源面臨巨大挑戰,呈現出群體小型化和低齡化的趨勢(賈永義,2019)。開展生物群體遺傳多樣性分析是采取有效保護措施的基礎(代應貴和肖海,2011),即明確翹嘴鲌遺傳多樣性,不僅有利于評估其種質資源狀況,還能為保護翹嘴鲌種質資源提供參考依據。【前人研究進展】線粒體DNA(mtDNA)具有結構簡單、進化速率適宜及母系遺傳的特點,適用于物種進化和分子多態性研究(陳四海等,2011;鄒輝等,2020)。魚類mtDNA主要包括37個編碼基因(13個疏水蛋白基因、2個rRNA基因和22個tRNA編碼基因)、1個負責復制和轉錄起始的控制區(D-loop)和1個輕鏈復制起始區(郭新紅等,2004),其中細胞色素氧化酶(COI、COII和COIII)是由疏水蛋白基因編碼。與其他線粒體蛋白編碼基因相比,基因的進化速率較快(彭居俐等,2009),常被用于研究親緣關系較近的種、種下分類單元及地理群體間的關系(王偉,2007;黃小彧,2012;楊子拓,2016)。近年來,國內外針對翹嘴鲌的研究主要集中在營養與飼料(王桂芹等,2008)、解剖學與生物學特性(Cao et al.,2011)、遺傳多樣性(黃小彧,2012;王丹等,2015)及人工繁殖和胚胎發育(周彥鋒等,2018;賈永義,2019)等方面,而基于mtDNA分析翹嘴鲌遺傳多樣性的研究主要集中在基因、D-Loop控制區和基因序列。王偉等(2008)研究證實了基因序列在翹嘴鲌群體遺傳學研究中的有效性,并分析黑龍江興凱湖至江蘇太湖6個水域的翹嘴鲌群體遺傳多樣性,認為通過基因可將我國翹嘴鲌進行區域劃分,但尚不能作為群體鑒別的分子標記;黃小彧(2012)基于線粒體D-Loop控制區分析長江水系14個翹嘴鲌群體的遺傳多樣性,結果發現翹嘴鲌群體具有高單倍型多樣性和低核苷酸多樣性共存現象;王丹等(2015)利用線粒體基因初步分析鲌屬魚類的遺傳多樣性,結果表明鲌屬魚類具有高的遺傳多樣性。【本研究切入點】受環境污染、棲息地被破壞及水利修建等因素的影響,翹嘴鲌野生資源瀕臨枯竭,且種群小型化、低齡化現象嚴重,但目前針對翹嘴鲌遺傳背景的系統研究較少,難以為其種質資源的保護工作提供可靠依據,尤其鮮見基于基因序列開展地理位置較近翹嘴鲌群體遺傳多樣性的研究報道。【擬解決的關鍵問題】在長江下游水域(淀山湖、高郵湖、太湖、長蕩湖及長江江蘇段)設點捕獲收集翹嘴鲌野生樣本,基于基因序列分析翹嘴鲌遺傳多樣性,明確其遺傳多樣性的豐富程度和進化歷史,為翹嘴鲌育種及遺傳改良打下基礎。
1 材料與方法
1.1 試驗材料
2019年5—9月,使用絲網分別在淀山湖(DS)、高郵湖(GY)、太湖(TH)、長蕩湖(CD)和長江(CJ)江蘇段設5個采樣水域(圖1)捕獲收集翹嘴鲌樣本,剪下尾鰭浸泡于95%酒精中,帶回實驗室備用。
1.2 形態學數據測量
使用精度為0.01 mm的游標卡尺測量所有翹嘴鲌的全長和體長,以精度為0.01 g的電子秤稱量魚體質量。當魚體長超過500.00 mm時,則使用卷尺和臺秤進行測量。
1.3 DNA提取
剪取備用鰭條組織樣品(<30.0 mg)置于離心管中,采用海洋動物組織基因組DNA提取試劑盒[天根生化科技(北京)有限公司]提取DNA,以1.0%瓊脂糖凝膠電泳檢測DNA質量。DNA樣品-20℃冰箱保存備用。
1.4 COII基因PCR擴增及測序
依據NCBI數據庫中的翹嘴鲌基因序列(AY704458)設計PCR擴增引物:-3(5'-TGAT CGCTCTGCCCCCCCT-3')、-5(5'-AGGATCAG CTCGAAGTGC-3')(王偉等,2008)。PCR反應體系25.0μL:dNTP Mixture 12.0μL,上、下游引 物(5μmol/L)各0.5μL,DNA模板2.0μL,ddHO 10.0μL。擴增程序:94℃預變性5 min;94℃10 s,59℃10 s,72℃30 s,進行35個循環;72℃延伸5 min;4℃結束。2.0%瓊脂糖凝膠電泳檢測PCR擴增產物,將條帶清晰的有效擴增產物送至亦欣生物科技(上海)有限公司進行雙向測序,測序引物與擴增引物一致。
1.5 數據分析
測量所有翹嘴鲌的體長和體質量,通過SPSS 19.0進行均值比較分析。采用ClustalX 1.83(Petren and Case,1997)和Seaview(Gouy et al.,2012)進行基因核苷酸組成及序列分析;使用DNASPv5(Librado and Rozas,2009)計算群體單倍型,并進行Tajima’s D和Fu’s Fs中性檢驗,以推測群體是否發生群體擴張;運用Arlequin 3.5(Excoffier and Lischer,2010)進行遺傳多樣性分析、群體分子方差分析(Analysis of molecular variance,AMOVA)及計算遺傳距離和基因流();在MEGA 6.0中以Kimura雙參數法(Kimura-2-parameter)為替代模型(Tamura et al.,2013),采用鄰接法(Neighbour-joining,NJ)構建翹嘴鲌群體單倍型系統發育進化樹,分析各群體間的親緣關系。
2 結果與分析
2.1 形態學數據統計分析結果
翹嘴鲌體長與體質量數據均為每個采樣點的平均數值,以太湖翹嘴鲌群體最大,其次是長江群體,而后依次是高郵湖群體、長蕩湖群體和淀山湖群體。共測序得到197個基因序列(表1)。

圖1 翹嘴鲌采樣水域分布情況Fig.1 Distribution of C.alburnus sampling waters

表1 5個翹嘴鲌野生群體的生物信息Table 1 Biological information of 5 wild C.alburnus populations

圖2 部分翹嘴鲌樣本的PCR擴增電泳結果Fig.2 PCR amplification electrophoresis of some C.alburnus samples
2.2 翹嘴鲌COII基因序列變異及單倍型分析結果
以提取的DNA為模板進行PCR擴增,運用2.0%瓊脂糖凝膠電泳檢測PCR擴增產物,部分電泳結果如圖2所示。5個翹嘴鲌野生群體197個個體的基因序列長度經比對后獲得的有效長度為425 bp,有31個變異位點(表2),變異率為7.29%。翹嘴鲌基因測序結果(圖3)表明,其堿基含量排序為T(29.61%)>C(28.03%)>A(24.19%)>G(18.17%)。AT含量為53.8%,CG含量為46.2%。
31個變異位點在197個個體中共定義出13種單倍型(表3)。其中,9個單倍型為各群體特有的單倍型,占69.23%,即hap6和hap7為高郵湖特有,hap4為淀山湖特有,hap8和hap9為長蕩湖所特有,長江群體特有4個單倍型(hap10、hap11、hap12和hap13)。此外,有4個單倍型(hap1、hap2、hap3和hap5)在翹嘴鲌群體中共享,占30.77%,有2個單倍型(hap1和hap2)在5個群體中均有分布。其中,又以hap1最常見,存在于138個個體中,是優勢單倍型;其次是hap2。太湖群體與淀山湖群體共享hap3。基于Kimura-2-parameter構建的翹嘴鲌群體單倍型系統發育進化樹(圖4)顯示,5個翹嘴鲌野生群體所特有的單倍型并未各自聚類形成獨立分支,而是相互交錯形成復雜的簇群,說明不同翹嘴鲌群體間的分化并不明顯。

表2 翹嘴鲌COII基因的變異位點Table 2 Variation sites of mtDNA COII gene of C.alburnus

圖3 部分翹嘴鲌COII基因的測序圖譜Fig.3 Sequencing map of COII gene in some C.alburnus

表3 COII基因在5個翹嘴鲌野生群體中的單倍型分布情況Table 3 Haplotype distribution of the mtDNA COII gene in the 5 populations of C.alburnus
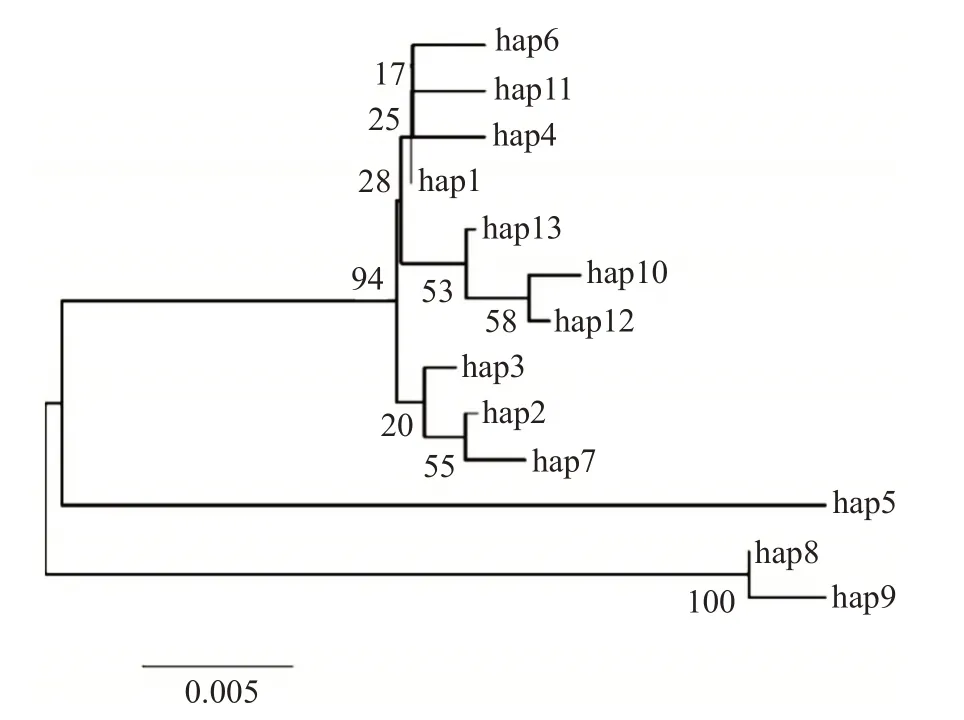
圖4 基于Kimura-2-parameter構建的翹嘴鲌群體單倍型系統發育進化樹Fig.4 Phylogenetic tree of haplotype of C.alburnus populations based on Kimura-2-parameter
2.3 遺傳多樣性分析結果
從5個翹嘴鲌野生群體197個個體中共檢測出13個單倍型,單倍型多樣性指數()為0.273~0.603,以長江群體和長蕩湖群體的最高,太湖群體的最低;核苷酸多樣性指數()為0.001~0.017,以長蕩湖群體的最高,淀山湖群體的最低(表4)。5個翹嘴鲌野生群體的為0.491,為0.007,說明其遺傳多樣性豐富。翹嘴鲌群體內的遺傳距離為0.001~0.018(表5),其中淀山湖群體內的遺傳距離最小,長蕩湖群體內的遺傳距離最大;不同群體間遺傳距離為0.002~0.015,即群體間無顯著差異。5個翹嘴鲌野生群體間的為2.443~54.325(表5),以太湖群體與淀山湖群體間的最大(54.325),長蕩湖群體與淀山湖群體間的最小(2.443)。分子方差分析(AMOVA)結果(表6)顯示,不同群體間的遺傳變異為10.47%,而群體內的遺傳變異為89.53%。
2.4 群體歷史動態分析結果
中性檢驗結果顯示:除了長蕩湖群體的Tajima’s D為正值外,其余群體的Tajima’s D均為負值(表4),且太湖群體表現為極顯著偏離中性理論下的Wright-Fisher模型(<0.01),長江群體表現為顯著偏離中性理論下的Wright-Fisher模型(<0.05)。Fu’s Fs檢驗結果表明,淀山湖和長江的Fu’s Fs為負值(表4),且長江群體具有統計學意義。5個翹嘴鲌野生群體的Tajima’s D和Fu’s Fs均為負值(-1.479和-0.547),但不具統計學意義。
3 討論
遺傳分化指數()是群體間遺傳分化的一個重要指標,常用于衡量2個群體間的遺傳分化程度:當<0.05時表示群體間幾乎無遺傳分化,當0.05<<0.15時表示群體間存在低度分化,當0.15<<0.25時表示群體間存在中度分化,當>0.25時表示群體間存在高度分化(黃小彧,2012)。本研究中,5個翹嘴鲌野生群體間為0.105,表明不同翹嘴鲌野生群體間存在低度分化。Hebert等(2003)對動物界11個門13320個物種的研究結果顯示,種內遺傳距離大多數在1%以下,很少超過2%。本研究結果表明,淀山湖、高郵湖、太湖、長蕩湖和長江群體的群體內遺傳距離分別為0.001、0.009、0.003、0.018和0.004,均符合Hebert等(2003)提出的觀點。將5個翹嘴鲌野生群體視為一個整體進行分析發現,翹嘴鲌群體的為0.491,為0.007,表現出低和高,說明具有較豐富的遺傳多樣性,與黃小彧(2012)基于線粒體D-Loop控制區研究長江水系14個翹嘴鲌群體遺傳多樣性得出的結論(為0.866,為0.0033)相反,究其原因可能是黃小彧(2012)研究利用的D-Loop控制區標記與本研究采用的基因不同,也有可能是禁捕政策的實施促使近年來長江水域翹嘴鲌的資源量有所增加所致。顏岳輝等(2019)曾研究證實波氏吻蝦虎()表現出低和高,與本研究結果相似,可能是歷史群體曾發生瓶頸效應,而后伴隨出現群體快速增長及積累新突變。
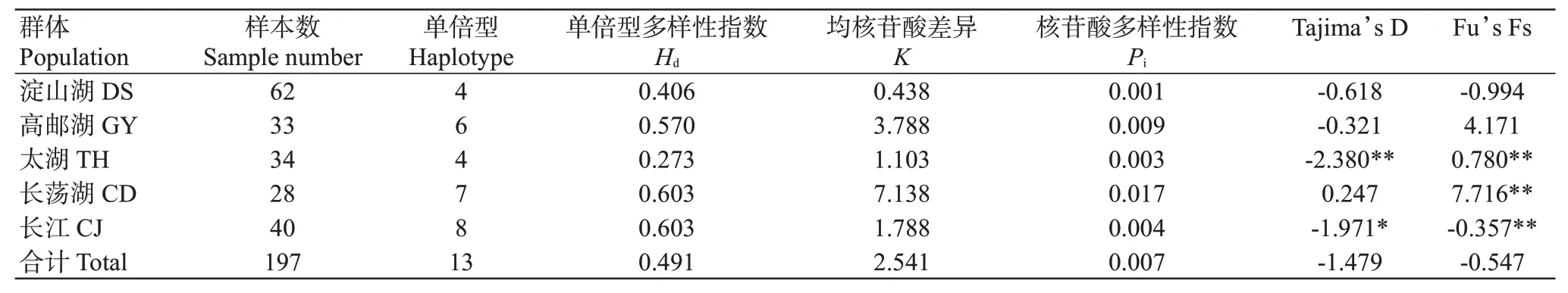
表4 5個翹嘴鲌野生群體的遺傳多樣性參數Table 4 Genetic diversity parameters of 5 C.alburnus populations

表5 不同翹嘴鲌群體的基因流和遺傳距離Table 5 Gene flow and genetic distance of different C.alburnus populations
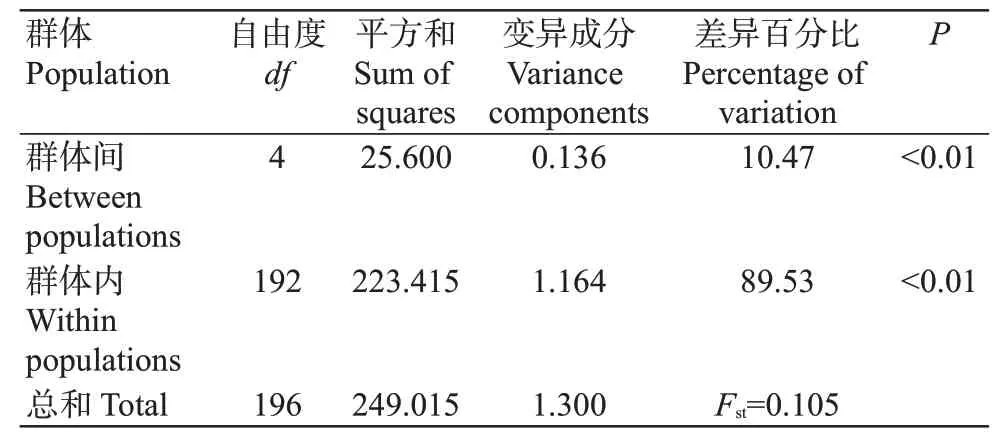
表6 不同翹嘴鲌野生群體間及群體內的分子方差分析結果Table 6 Analysis of molecular variance between and within the C.alburnus populations
人類活動在很大程度上會影響群體間的基因交流,如人工引種和增殖放流等。基因交流是指生物個體從其發生地分散出去而導致不同群體間基因雜交的過程,其基本作用是削弱生物群體間的遺傳差異(Grant and Bowen,1998)。如大于1.000,表明在群體中會有較高水平的基因交流,尤其是>4.000表明群體間存在頻繁的基因交流(Mcmillen-Jackson and Bert,2004)。本研究中,5個翹嘴鲌野生群體的在2.443~54.325,且長蕩湖群體與其他水域群體間的均較小(2.443~6.179)。從本研究采樣區域的地理位置來看,長蕩湖與其他水系不通,由于地理隔離造成該水域翹嘴鲌群體與其他水域翹嘴鲌群體間存在基因交流障礙;而太湖的地理條件優越、物產豐富,先后建立了銀魚()、秀麗白蝦()、翹嘴鲌和青蝦()等國家級水產種質資源保護區(谷孝鴻等,2009),為不同地理群體的基因交流創造了條件。太湖漁業資源調查發現,隨著人類捕撈活動的加劇及棲息環境被破壞,翹嘴鲌多樣性逐漸降低,魚種趨于小型化和單一化(賈永義,2019),也可能是其他因素導致太湖翹嘴鲌群體遺傳多樣性降低,具體原因有待進一步探究。翹嘴鲌性情活潑好動,在無地理隔離條件下群體間的基因交流可能是導致不同群體形態相近的主要原因之一(王偉,2007)。淀山湖群體與太湖群體間的為54.325,表明這2個水域的翹嘴鲌存在一定的基因交流,是由于淀山湖與太湖距離相對較近,且歷史上有河流連通;高郵湖群體與長蕩湖群體間的遺傳距離最大(0.015),則因地理隔離形成了穩定的地理群體;由于地理隔離的原因,長蕩湖與其他幾個水域間無河流連通,因此長蕩湖翹嘴鲌群體與其他湖泊的翹嘴鲌群體間幾乎無基因交流。
本研究對不同翹嘴鲌野生群體及整體分別進行Tajima’s D檢驗和Fu’Fs檢驗,以推測翹嘴鲌群體是否經歷過群體擴張事件。當Tajima’s D和Fu’Fs均為負值且具有顯著意義時,則認為該群體曾發生過群體擴張現象(Fu,1997;劉偉等,2016)。在5個翹嘴鲌野生群體中,僅長江群體的Tajima’s D和Fu’Fs均為負值且具有顯著意義,故推測該群體曾發生過群體擴張現象。
4 結論
長江下游翹嘴鲌群體表現出低單倍型多樣性和高核苷酸多樣性,遺傳多樣性較豐富,群體間存在遺傳分化但分化程度差異不顯著。因此,后續研究應增加野生翹嘴鲌群體的樣品數量和調查水域,全面而系統地評估長江下游各水域翹嘴鲌的種質資源狀況,為建立自然保護區及人工增殖放流提供科學依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
