斑馬魚zgc113227多克隆抗體及卵形鯧鲹EVM0008813多肽抗體的制備
2022-10-11 12:52:30張永德宋漫玲羅輝余燕玲林勇陳曉漢馮鵬霏羅洪林
南方農(nóng)業(yè)學(xué)報 2022年7期
張永德,宋漫玲,羅輝,余燕玲,林勇,陳曉漢,馮鵬霏,羅洪林*
(1廣西水產(chǎn)科學(xué)研究院/廣西水產(chǎn)遺傳育種與健康養(yǎng)殖重點實驗室,廣西南寧 530021;2西南大學(xué)水產(chǎn)學(xué)院,重慶 402460)
0 引言
【研究意義】斑馬魚()原產(chǎn)于印度、巴基斯坦等熱帶地區(qū),除了具有較高的經(jīng)濟價值外,其生理組織結(jié)構(gòu)及分子調(diào)控等與人類相似,與人類基因的同源性高達87%,且飼養(yǎng)成本低、繁殖周期短、產(chǎn)量高,已廣泛應(yīng)用于脊椎動物發(fā)育機制、免疫學(xué)、藥物研發(fā)、毒理學(xué)評估及人類疾病模型研究等領(lǐng)域(Cross et al.,2003;吳唯維等,2014;段蕊和吳昊澤,2017;程祖春等,2019;錢星宇,2019)。卵形鯧鲹()俗名金鯧,已被列為名貴食用魚類,主要生活在熱帶、亞熱帶海域;隨著國內(nèi)外消費市場需求的不斷上升,卵形鯧鲹將發(fā)展成為最具潛力的海水經(jīng)濟養(yǎng)殖魚類之一(林川等,2018;張永德等,2020;段家文等,2021)。抗體是脊椎動物在應(yīng)對內(nèi)外抗原刺激的免疫應(yīng)答中,由漿細胞(效應(yīng)B細胞)分泌的一類能與相應(yīng)抗原特異性結(jié)合、具有免疫功能的球蛋白。除了本身作為生物學(xué)功能的概念,抗體還可作為一種研究工具應(yīng)用于科研、醫(yī)學(xué)、農(nóng)業(yè)、環(huán)境保護和食品檢測等領(lǐng)域,按其來源可分為天然抗體、單克隆抗體、多克隆抗體及基因工程抗體四大類(賈慧娜和羅海玲,2012;趙久剛等,2018),其中,多克隆抗體反應(yīng)強度大,且制備技術(shù)簡單,省時省力,價格低廉,因此在科研領(lǐng)域已得到廣泛應(yīng)用。【前人研究進展】目前,有關(guān)多克隆抗體制備的工藝已較成熟(張佳鳳等,2015;何麗番和高海,2016;李歌等,2019;馮鵬霏等,2021),在水生生物研究領(lǐng)域也取得長足進展。王雪妹(2015)制備獲得的抗鰻弧菌多克隆抗體為實現(xiàn)快速、靈敏檢測鰻弧菌提供了可靠工具。張永德等(2018)為研究羅非魚淋巴細胞特異性蛋白酪氨酸激酶(Lck)蛋白功能及其作用機制,制備了尼羅羅非魚Lck多克隆抗體并進行鑒定。何明旺等(2019)通過克隆卵形鯧鲹抗菌肽基因,構(gòu)建原核表達載體并體外誘導(dǎo)表達重組融合蛋白,成功制備獲得高效的hepcidin-5蛋白多克隆抗體,為深入探究卵形鯧鲹hepcidin-5的生物學(xué)功能奠定了基礎(chǔ)。潘傳燕等(2019)通過構(gòu)建原核表達載體、大腸桿菌融合表達及免疫日本大耳兔,成功制備獲得尼羅羅非魚過氧化物酶體增殖物激活受體δ(PPARδ)多克隆抗體。丁寧(2020)通過制備大黃魚鼠抗CD3ε多克隆抗體,探究了細胞毒性因子CD3分子在硬骨魚T細胞活化及免疫應(yīng)答中的功能作用。劉倩等(2020)制備獲得抗鼠源類固醇激素合成急性調(diào)節(jié)蛋白(Steroidogenic acute regulatory protein,StAR)多克隆抗體并應(yīng)用于中華鱉卵巢組織中StAR蛋白表達和定位分析,結(jié)果顯示StAR蛋白在卵巢間質(zhì)細胞和精巢的間質(zhì)細胞、精原細胞及胞質(zhì)中均有表達。王鷺等(2020)構(gòu)建了卵形鯧鲹腫瘤壞死因子超家族成員6(Tumor necrosis factor superfamily 6,TNFSF6)原核表達系統(tǒng)并成功制備獲得TNFSF6多克隆抗體。Nguyen-Phuoc等(2021)成功制備獲得蝦急性肝胰壞死病(AHPND)ToxA特異性多克隆抗體,為下一步研發(fā)基于抗原—抗體相互作用的AHPND檢測試劑盒奠定了基礎(chǔ)。【本研究切入點】本課題組前期研究卵形鯧鲹轉(zhuǎn)錄組時通過加權(quán)基因共表達網(wǎng)絡(luò)分析(Weighted correlation network analysis,WGCNA)發(fā)現(xiàn)1個與卵形鯧鲹胚胎發(fā)育相關(guān)基因(EVM0008813),同時經(jīng)同源序列查找獲得斑馬魚同源基因zgc113227(NP_001014341),但至今未見有關(guān)斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因功能特性的研究報道。【擬解決的關(guān)鍵問題】為深入研究斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因的生物學(xué)功能,根據(jù)其基因序列人工合成斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因,構(gòu)建對應(yīng)的原核表達載體,經(jīng)大腸桿菌融合表達后免疫日本大耳兔以制備多克隆抗體,為后續(xù)深入研究這2個基因的功能特性提供重要工具。
1 材料與方法
1.1 試驗材料
TRIzol試劑購自Invitrogen公司,ET-B2m表達載體和大腸桿菌B21感受態(tài)細胞購自武漢金開瑞生物工程有限公司,質(zhì)粒提取試劑盒(DP116)和膠回收試劑盒(DP209)購自天根生化科技(北京)有限公司,親和層析柱料購自美國GE Healthcare公司,T4 DNA連接酶、DNA聚合酶、蛋白分子量標(biāo)準(zhǔn)(Marker)及限制性內(nèi)切酶(H I和I)購自美國Fermentas公司,羊抗兔-HRP抗體購自美國Jackson公司,弗式完全佐劑和弗氏不完全佐劑購自美國Sigma公司。
1.2 試驗方法
1.2.1 目的基因合成從卵形鯧鲹轉(zhuǎn)錄組中篩選出1個與胚胎發(fā)育相關(guān)的基因(EVM0008813),以EVM0008813基因為查詢序列,對斑馬魚全基因組序列進行BLASTp搜索(E<e-10),獲得斑馬魚同源基因zgc113227(NP_001014341)。利用Primer Premier 5.0設(shè)計特異性引物(表1)用于檢測表達載體中的斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因,委托生工生物工程(上海)股份有限公司合成基因序列及PCR擴增引物。

表1 斑馬魚zgc113227和卵形鯧鲹EVM0008813基因的PCR擴增引物Table 1 PCR amplification primers of zebrafish zgc113227 and T.ovatus EVM0008813 genes
1.2.2 蛋白結(jié)構(gòu)分析及抗原表位預(yù)測采用ExPASy(https://web.expasy.org/protparam/)預(yù)測蛋白親/疏水性等理化性質(zhì),運用TMHMM 2.0(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)進行蛋白跨膜結(jié)構(gòu)和信號肽預(yù)測,利用DNASTAR中的Protean模塊對蛋白抗原指數(shù)、柔性區(qū)域、表面可及性等參數(shù)進行預(yù)測,使用SMART(http://smart.embl.de/)和MEME(https://meme-suite.org/meme/)預(yù)測分析蛋白結(jié)構(gòu)域,并以Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)進行序列同源比對分析。
1.2.3 原核表達載體構(gòu)建及鑒定以同源重組方式構(gòu)建原核表達載體pET-B2m-zgc113227和pETB2m-EVM0008813,然后轉(zhuǎn)化B21感受態(tài)細胞;篩選陽性克隆并提取質(zhì)粒進行PCR擴增,具體PCR反應(yīng)體系及擴增程序參照潘傳燕等(2019)的研究方法。PCR擴增產(chǎn)物經(jīng)2.0%瓊脂糖凝膠電泳檢測后送至生工生物工程(上海)股份有限公司測序。
1.2.4 融合蛋白表達與鑒定以原核表達載體pET-B2m-zgc113227和pET-B2m-EVM0008813分別轉(zhuǎn)化B21感受態(tài)細胞,挑選陽性菌體接種至含抗生素的LB液體培養(yǎng)基中,37℃恒溫培養(yǎng)至A≈0.6;取部分菌液為對照,剩余菌液加入IPTG(終濃度0.5 mmol/L),繼續(xù)誘導(dǎo)培養(yǎng)3 h;兩組菌液各取0.15 mL,12000×g離心2 min,菌體沉淀以40μL 1×Loading Buffer重懸裂解,12000×g離心10 min后收集上清液進行SDS-PAGE檢測。
1.2.5 zgc113227蛋白和EVM0008813多肽大量表達與純化陽性菌體經(jīng)IPTG誘導(dǎo)大量表達后,收集發(fā)酵液,8000×g離心5 min并收集菌體。將菌體懸浮于預(yù)冷的NTA-0緩沖液中冰浴30 min,超聲波破碎菌體,12000×g離心10 min,收集上清液及菌體沉淀。菌體沉淀根據(jù)Ni-NTA操作說明進行蛋白純化,并以SDS-PAGE檢測斑馬魚zgc113227蛋白和卵形鯧鲹EVM0008813多肽的純化情況。
1.2.6 zgc113227蛋白和EVM0008813多肽抗體制備選用健康級成年日本大耳白兔為試驗動物,將純化的zgc113227蛋白及EVM0008813多肽與牛血清白蛋白(BSA)偶聯(lián)物分別與等容積弗氏完全佐劑充分乳化后,于日本大耳白兔皮下進行多點注射,各免疫2只,免疫劑量為500μg/只。每只日本大耳白兔免疫5~6次,每次免疫間隔14 d,待抗體表達恒定后采血分離血清,分裝,-80℃保存?zhèn)溆谩?/p>
1.2.7 免疫效價測定采用間接ELISA測定抗血清效價。以純化zgc113227蛋白為包被抗原(2μg/mL),吸取樣品按100μL/孔的劑量包被96孔酶聯(lián)板,4℃過夜;棄孔內(nèi)液體,洗滌3次,加入待測血清,37℃溫育1 h;洗滌,再加入1∶5000稀釋的酶標(biāo)二抗(100μL/孔),37℃溫育1 h,洗滌后每孔加入100μL顯色液(TMB),37℃溫育20 min,加入50μL終止液終止反應(yīng),采用全波長酶標(biāo)儀測定吸光值(OD)。陽性反應(yīng)的最大稀釋度即為待測抗體效價。抗EVM0008813多肽血清效價測定方法同上。
1.2.8 斑馬魚zgc113227多克隆抗體鑒定及純化采用Western blotting鑒定zgc113227蛋白:分別取10和25 ng純化的融合蛋白進行SDS-PAGE電泳,并轉(zhuǎn)印至PVDF膜,封閉液中37℃封閉2 h后加入稀釋1000倍的兔抗血清,37℃搖床孵育1 h,洗膜,然后加入羊抗兔-HRP二抗(1∶10000倍稀釋),37℃孵育1 h,采用ECL化學(xué)發(fā)光試劑盒發(fā)光顯影,暗室反應(yīng)2 min,取出膠片立即浸入顯影液中顯色1 min,再浸入定影液中定影1 min,晾干,拍照分析。采用Protein G親和層析柱對抗體進行純化,將收集的抗zgc113227血清與4倍體積的1% NaAc混合后上樣,1% NaAc洗雜,以3.5%冰乙酸沖洗至無蛋白流出檢出,收集洗脫產(chǎn)物,測定OD并估算抗體濃度;采用飽和碳酸鈉調(diào)節(jié)洗脫產(chǎn)物pH至7.0,使用10 kD超濾管進行純化濃縮,經(jīng)SDS-PAGE檢測后-20℃保存?zhèn)溆谩?/p>
1.2.9 卵形鯧鲹EVM0008813多肽抗體鑒定及純化采用高效液相色譜—質(zhì)譜聯(lián)用(HPLC-MS)純化鑒定卵形鯧鲹EVM0008813多肽抗體。通過査閱相關(guān)文獻,確定選用乙腈三氟乙酸水溶液系統(tǒng),色譜流動相為0.1%三氟乙酸水溶液—乙腈系統(tǒng),梯度洗脫,參照孟迂(2010)以HPLC分離純化蜂毒多肽的方式進行具體操作。
2 結(jié)果與分析
2.1 斑馬魚zgc113227和卵形鯧鲹EVM0008813跨膜結(jié)構(gòu)、信號肽及理化性質(zhì)預(yù)測結(jié)果
斑馬魚zgc113227基因編碼415個氨基酸殘基,編碼蛋白分子量為48091.58 Da,理論等電點(pI)為9.02,不穩(wěn)定性指數(shù)為55.00,脂肪指數(shù)為92.22。卵形鯧鲹EVM0008813基因編碼395個氨基酸殘基,編碼蛋白分子量為45397.07 Da,pI為9.05,不穩(wěn)定性指數(shù)為46.33,脂肪指數(shù)為87.11。跨膜結(jié)構(gòu)及信號肽預(yù)測結(jié)果顯示,斑馬魚zgc113227和卵形鯧鲹EVM0008813均無跨膜結(jié)構(gòu)及信號肽存在。
2.2 斑馬魚zgc113227和卵形鯧鲹EVM0008813抗原表位參數(shù)預(yù)測結(jié)果
2.2.1 斑馬魚zgc113227親/疏水性、抗原指數(shù)、柔性區(qū)域和表面可及性DNASTAR預(yù)測結(jié)果(圖1)顯示,斑馬魚zgc113227蛋白平均親水性為-0.49,其中親水性氨基酸占比較高,主要包括第30~45、55~72、78~112、129~138、168~188、205~220、244~253、264~273、287~290、292~296、303~321、323~345和360~400位氨基酸。抗原指數(shù)高低決定了蛋白抗原表位位點存在的可能性。斑馬魚zgc113227氨基酸序列潛在多個且均勻分布的抗原表位區(qū)域,且這些位點的抗原指數(shù)較高,分別是第36~48、53~58、62~69、75~82、87~111、115~121、129~136、170~187、205~220、229~232、241~255、261~277、297~331、336~350、360~401和406~411位氨基酸。此外,斑馬魚zgc113227蛋白柔性較強、表面可及性高,具有制備蛋白抗體的優(yōu)勢。
2.2.2 卵形鯧鲹EVM0008813親/疏水性、抗原指數(shù)、柔性區(qū)域和表面可及性DNASTAR預(yù)測結(jié)果(圖2)顯示,卵形鯧鲹EVM0008813蛋白氨基酸序列中含有較多的親水性氨基酸,平均親水性為-0.49,親水性區(qū)段較高的主要有第22~48、52~58、63~72、77~95、101~112、117~121、130~136、170~175、182~190、206~220、244~274、277~308、313~333、346~364、370~381和389~395位 氨 基 酸。卵 形 鯧 鲹EVM0008813氨基酸序列也潛在多個抗原表位位點且分布均勻,其中第30~47、52~58、63~69、75~80、87~94、103~111、117~121、130~136、170~177、183~189、196~199、205~220、228~233、242~252、261~308、311~337、346~363、372~385、391~395位氨基酸的抗原指數(shù)較高,同時具備較強的親水區(qū)域,制備抗體的可行性較高。
2.3 斑馬魚zgc113227和卵形鯧鲹EVM0008813蛋白結(jié)構(gòu)域預(yù)測結(jié)果
Clustal Omega預(yù)測得知斑馬魚zgc113227和卵形鯧鲹EVM0008813的氨基酸序列相似性為58.12%。利用SMART預(yù)測蛋白結(jié)構(gòu)域,結(jié)果發(fā)現(xiàn)斑馬魚zgc113227氨基酸序列C端第195~358位氨基酸包含1個DDE_Tnp_4結(jié)構(gòu)域,卵形鯧鲹EVM0008813氨基 酸 序 列C端 第195~344位 氨 基 酸 也 含 有1個DDE_Tnp_4結(jié)構(gòu)域,故推測斑馬魚zgc113227和卵形鯧鲹EVM0008813均屬于DDE_Tnp_4家族。采用MEME對斑馬魚zgc113227和卵形鯧鲹EVM000881進行保守結(jié)構(gòu)域分析,結(jié)果(圖3)均獲得5個保守基序(Motif 1~Motif 5),且這5個基序是氨基酸序列中最保守的區(qū)域。
2.4 斑馬魚zgc113227和卵形鯧鲹EVM0008813表達載體鑒定結(jié)果
合成斑馬魚zgc113227和卵形鯧鲹EVM0008813基因序列,通過同源重組分別構(gòu)建原核表達載體pET-B2m-zgc113227和pET-B2m-EVM0008813。然后以構(gòu)建的2個表達載體為模板,PCR擴增zgc113227和EVM0008813基因,經(jīng)2.0%瓊脂糖凝膠電泳檢測,分別獲得約1500和1000 bp的擴增片段(圖4),片段大小與預(yù)期結(jié)果一致。經(jīng)測序與序列比對分析,證實斑馬魚zgc113227和卵形鯧鲹EVM0008813表達載體成功構(gòu)建,目的基因片段插入位置正確。
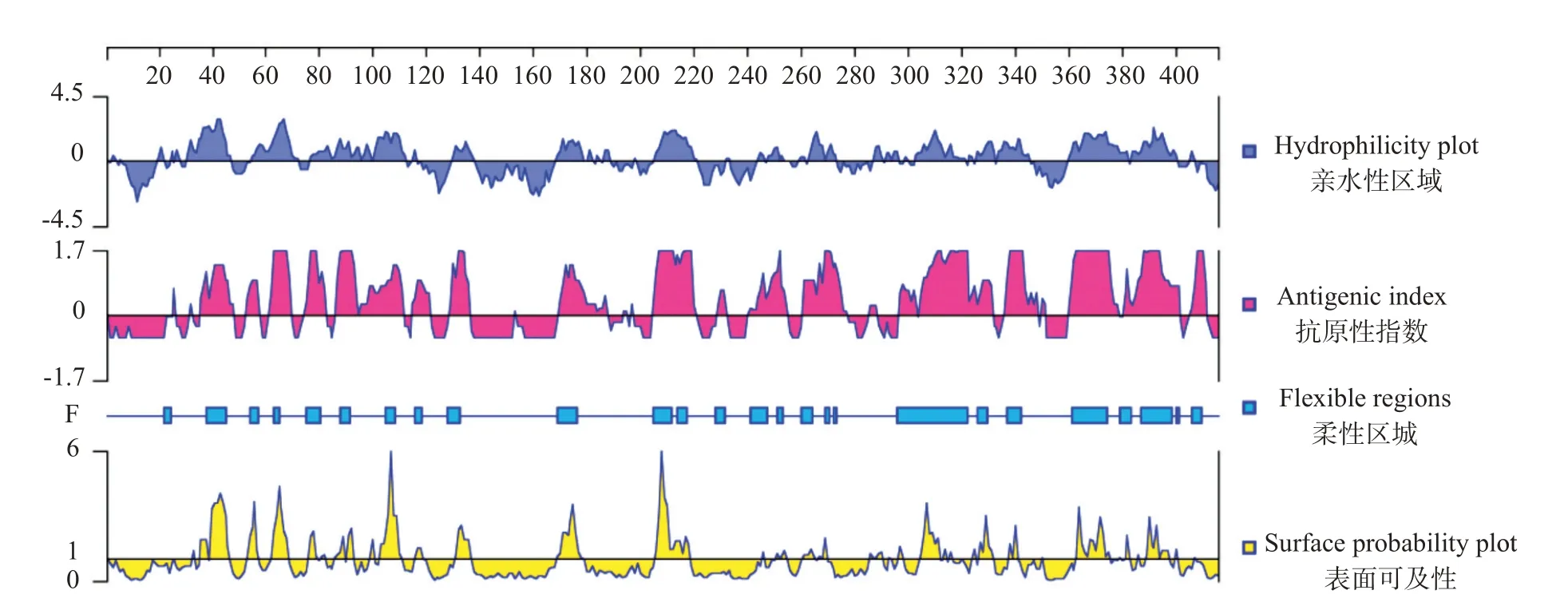
圖1 斑馬魚zgc113227蛋白親/疏水性、抗原指數(shù)、柔性區(qū)域和表面可及性等參數(shù)預(yù)測結(jié)果Fig.1 Parameter prediction of hydrophilicity,antigen index,flexible region and surface probability of zgc113227 protein in zebrafish
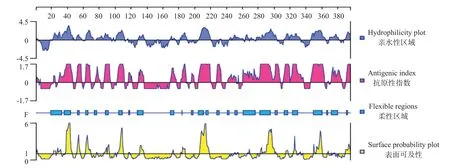
圖2 卵形鯧鲹EVM0008813多肽親/疏水性、抗原指數(shù)、柔性區(qū)域和表面可及性等參數(shù)預(yù)測結(jié)果Fig.2 Parameter prediction of hydrophilicity,antigen index,flexible region and surface probability of EVM0008813 polypeptide in T.ovatus
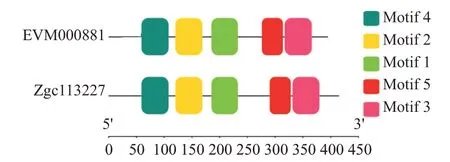
圖3 斑馬魚zgc113227和卵形鯧鲹EVM0008813蛋白的保守基序預(yù)測結(jié)果Fig.3 Conserved motif prediction of zebrafish zgc113227 and T.ovatus EVM0008813
2.5 斑馬魚zgc113227和卵形鯧鲹EVM0008813原核表達鑒定結(jié)果
以構(gòu)建的原核表達載體pET-B2m-zgc113227和pET-B2m-EVM0008813分別轉(zhuǎn)化B21感受態(tài)細胞,經(jīng)0.5 mmol/L IPTG誘導(dǎo)3 h,結(jié)果發(fā)現(xiàn)均能獲得對應(yīng)的目的蛋白條帶,其中,zgc113227蛋白在62.0 kD處有1條明顯的目的條帶(圖5-A),EVM0008813多肽在39.0 kD處出現(xiàn)1條明顯的目的條帶(圖5-B)。
2.6 zgc113227蛋白和EVM0008813多肽的提取與純化效果
收集成功表達融合蛋白的大腸桿菌進行裂解提取并純化,稀釋上樣經(jīng)SDS-PAGE檢測,結(jié)果顯示,在62.0 kD處有1條清晰條帶(圖6-A),與預(yù)期的斑馬魚zgc113227蛋白分子量相符;在39.0 kD處也出現(xiàn)明顯的目的條帶(圖6-B),與預(yù)期的卵形鯧鲹EVM0008813多肽分子量相符。
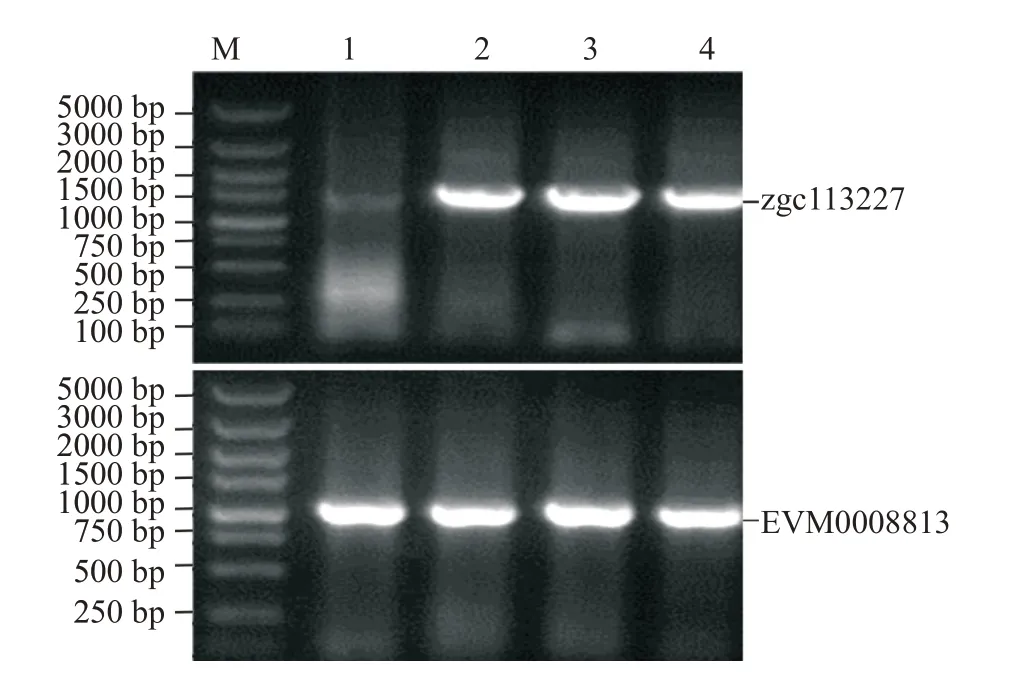
圖4 斑馬魚zgc113227和卵形鯧鲹EVM0008813基因表達載體的PCR檢測結(jié)果Fig.4 PCR detection of gene expression vector in zebrafish zgc113227 and T.ovatus EVM0008813
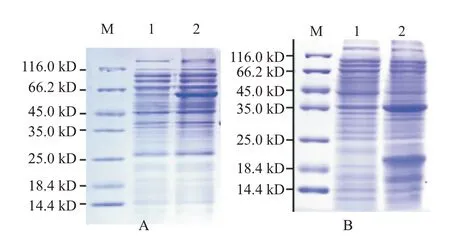
圖5 斑馬魚zgc113227和卵形鯧鲹EVM0008813蛋白的誘導(dǎo)表達產(chǎn)物SDS-PAGE檢測結(jié)果Fig.5 Induced expression product SDS-PAGE detection of zebrafish zgc113227 and T.ovatus EVM0008813
2.7 多克隆抗體效價測定結(jié)果
以免疫前的血清為陰性對照,通過間接ELISA測定抗血清效價。結(jié)果顯示,zgc113227多克隆抗體效價為1∶256000(圖7),具有高水平的特異性;將EVM0008813多肽與BSA偶聯(lián)成完全抗原并免疫日本大耳兔制備的抗血清,抗體效價超過1∶256000(圖8),說明EVM0008813多肽抗體具有良好的免疫效果。
2.8 zgc113227多克隆抗體特異性檢測及純化結(jié)果
采用Western blotting鑒定斑馬魚zgc113227多克隆抗體特異性,結(jié)果(圖9-A)顯示,在62.0 kD處有1條清晰的特異性蛋白條帶,說明制備的zgc113227多克隆抗體有良好的特異性。經(jīng)Protein G親和層析柱純化抗體,將純化后的抗體進行SDS-PAGE鑒定,結(jié)果(圖9-B)顯示分別在62.0和31.0 kD處出現(xiàn)1條清晰重鏈條帶和1條輕鏈條帶,表明zgc113227蛋白多克隆抗體純化成功。
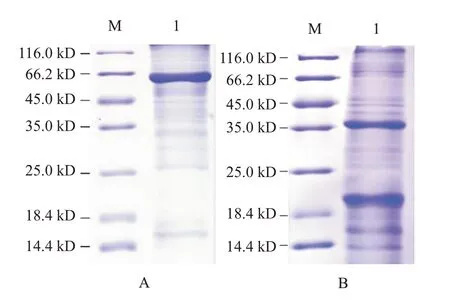
圖6 斑馬魚zgc113227蛋白和卵形鯧鲹EVM0008813多肽純化產(chǎn)物的SDS-PAGE檢測結(jié)果Fig.6 Purified polypeptide product SDS-PAGE detection of zebrafish zgc113227 and T.Ovatus EVM0008813
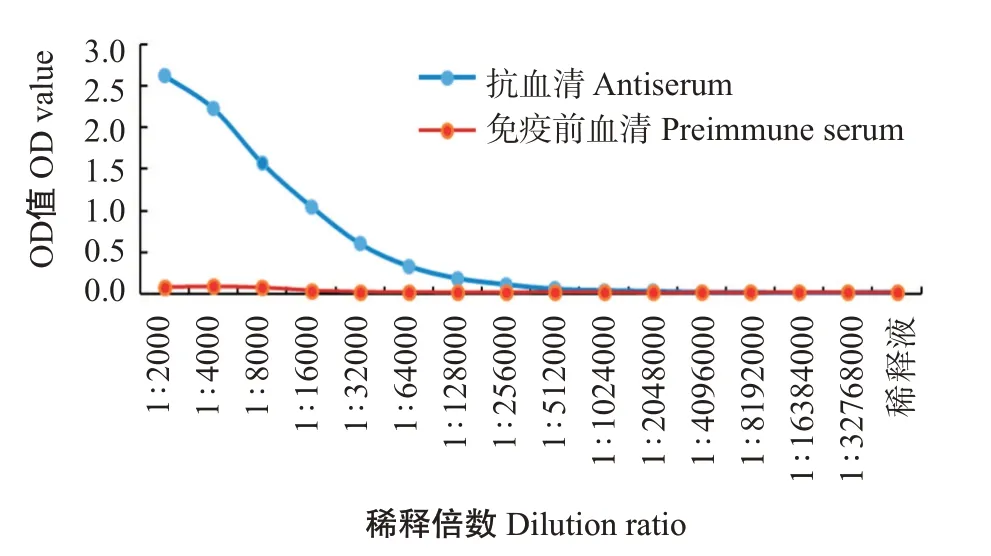
圖7 斑馬魚zgc113227多克隆抗體效價曲線Fig.7 Titer curves of polyclonal antibody of zebrafish zgc113227 protein
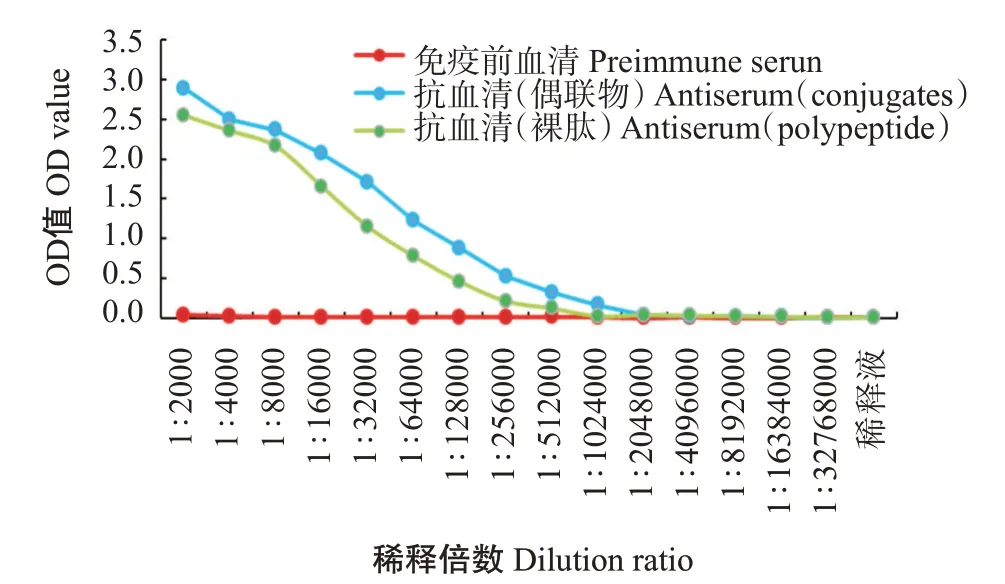
圖8 卵形鯧鲹EVM0008813多肽抗體效價曲線Fig.8 Titer curves of polyclonal antibody of T.ovatus EVM0008813 polypeptide
2.9 EVM0008813多肽抗體純化與鑒定結(jié)果
為獲得高純度的EVM0008813多肽抗體,采用HPLC-MS進行純化鑒定,結(jié)果(表2)表明,EVM0008813多肽抗體主峰集中在8~9 min,平均為8.601 min,無雜峰出現(xiàn),說明制備的多肽抗體純度較高,由峰面積計算可知其純度達95.35%,達到預(yù)期值。
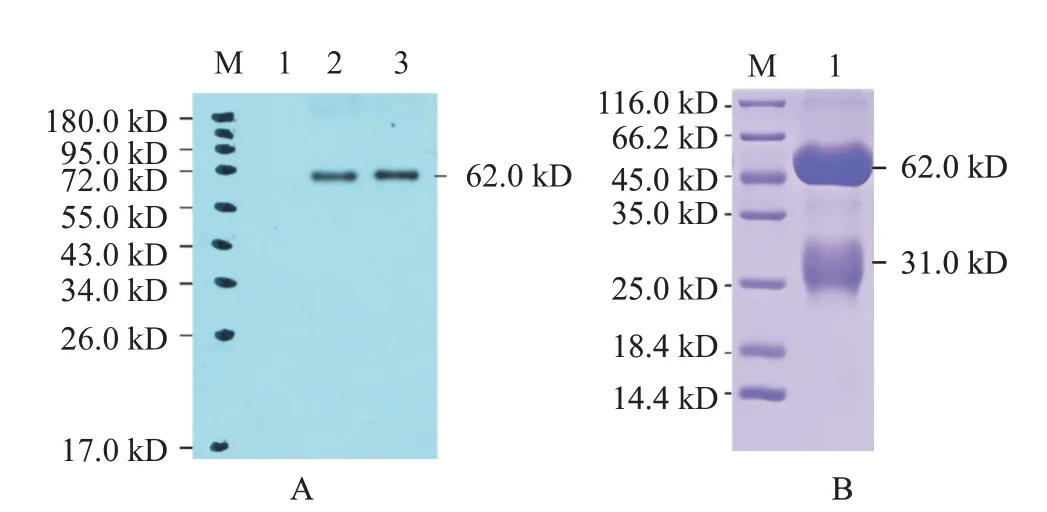
圖9 zgc113227多克隆抗體Western blotting鑒定及SDS-PAGE檢測結(jié)果Fig.9 Western blotting identification and SDS-PAGE detection of zgc113227 polyclonal antibody

表2 EVM0008813多肽抗體純度HPLC-MS檢測結(jié)果Table 2 Purity HPLC-MS detection of EVM0008813 polyclonal antibody
3 討論
本課題組前期在卵形鯧鲹轉(zhuǎn)錄組研究中發(fā)現(xiàn)與胚胎發(fā)育相關(guān)的EVM0008813基因,以此為查詢序列,通過BLASTp搜索獲得斑馬魚同源基因zgc113227(NP_001014341)。生物信息學(xué)分析結(jié)果表明,斑馬魚zgc113227和卵形鯧鲹EVM0008813均屬于親水性蛋白,二者的氨基酸序列相似性達58.12%,同時表現(xiàn)為潛在抗原位點多、柔性較強、表面可及性高,且無跨膜結(jié)構(gòu)和信號肽存在。基因表達調(diào)控的關(guān)鍵步驟是轉(zhuǎn)錄因子(TF)與其DNA識別位點的序列特異性結(jié)合(Huber and Bulyk,2006)。由于轉(zhuǎn)錄因子結(jié)合位點(TFBS)通常較短,且典型的序列特異性TF與彼此相似的位點結(jié)合,進而獲得一個稱為基序的保守模式,因此識別分析基序在研究調(diào)控基因轉(zhuǎn)錄表達方面具有重要意義(Bulyk,2003)。本研究采用MEME分析斑馬魚zgc113227和卵形鯧鲹EVM000881的保守結(jié)構(gòu)域,結(jié)果均獲得5個保守基序(Motif 1~Motif 5),且這5個基序是氨基酸序列中最保守的區(qū)域,故推測zgc113227基因和EVM0008813基因在轉(zhuǎn)錄表達過程中扮演著相似的角色。此外,在斑馬魚zgc113227和卵形鯧鲹EVM0008813的分子中均發(fā)現(xiàn)1個相同的保守結(jié)構(gòu)域(DDE_Tnp_4),推測二者均屬于DDE_Tnp_4家族。DDE_Tnp_4家族蛋白大部分為轉(zhuǎn)座酶家族,在催化DNA斷裂和重組過程中起關(guān)鍵作用(劉建政,2018)。DNA轉(zhuǎn)座是基因多樣性產(chǎn)生的關(guān)鍵步驟,其通過催化核酸酶結(jié)構(gòu)域折疊進行DNA位移,以混合的α-螺旋特征性RNase H樣折疊結(jié)構(gòu)(RNase H-like catalytic domains)最常見,其催化中心由2個天冬氨酰(DD)殘基和1個谷氨酸(E)殘基組成,雖然殘基的排列不同,但通過蛋白折疊可形成對移位所必需的DD(E/D)結(jié)構(gòu)域參與催化(Rice and Mizuuchi,1995;Haren et al.,1999;de Palmenaer et al.,2008),即斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因均參與基因轉(zhuǎn)錄。
本研究在設(shè)計卵形鯧鲹EVM0008813基因擴增引物時,通過軟件預(yù)測和分析該蛋白的結(jié)構(gòu)特征與抗原表位,選擇其編碼蛋白三維結(jié)構(gòu)無法暴露且抗原性高的非跨膜區(qū),兼顧處于連接二級結(jié)構(gòu)(SS)基序長環(huán)中的肽鏈,又避免位于螺旋區(qū)域中的肽鏈,其目的是增加抗體識別天然蛋白的概率,同時與斑馬魚zgc113227基因同源性高、具有穩(wěn)定構(gòu)象的B細胞線性表位(張佳鳳等,2015)的多肽基因序列作為擴增引物。對斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因進行人工合成,然后分別克隆至表達載體pET-B2m(+)上,構(gòu)建原核表達載體進行IPTG誘導(dǎo)表達。融合蛋白的表達效果受溫度及誘導(dǎo)時間的影響尤為重要,為了確保表達的融合蛋白具備良好活性且產(chǎn)量高,本研究選擇在37℃、A=0.6的條件下進行zgc113227蛋白和EVM0008813多肽誘導(dǎo)表達。此外,在免疫環(huán)節(jié)由于EVM0008813多肽僅由12個氨基酸組成,分子量較小,穩(wěn)定性較差,在動物體內(nèi)易被降解,且小分子多肽的抗原性普遍不高,無法激起足量的免疫反應(yīng)誘導(dǎo)動物產(chǎn)生抗體。本研究中,zgc113227蛋白與等容積弗氏完全佐劑混合為抗原,而EVM0008813多肽與BSA偶聯(lián)成完全抗原,然后經(jīng)皮下多點注射方式免疫日本大耳兔,間接ELISA檢測結(jié)果證實,兔抗zgc113227血清和兔抗EVM0008813血清效價均達1∶256000,表明獲得了高效價的多克隆抗體。
經(jīng)Western blotting鑒定和Protein G親和層析柱純化,zgc113227多克隆抗體特異性好,純度高,可滿足后續(xù)實驗需求;而采用Protein G親和層析柱純化EVM0008813多肽抗體,發(fā)現(xiàn)所得目的多肽純度較低,質(zhì)量差,可能是EVM0008813多肽與層析介質(zhì)不能特異性結(jié)合,或鏈接載體后表位暴露不充分造成結(jié)合效果不理想,或親和層析柱方法不適宜大量多肽純化等原因所造成。本研究將親和層析柱純化法換成HPLC-MS進行純化,得到的EVM0008813多肽抗體純度達95.35%,純度較高。此外,在檢測EVM0008813多肽抗體特異性時發(fā)現(xiàn)以純化后的EVM0008813多肽抗體進行Western blotting檢測無法檢測到特異性條帶,可能是EVM0008813多肽僅含有12個氨基酸,而目的蛋白氨基酸數(shù)量大(395個),二者結(jié)合概率較小,或?qū)υ摰鞍捉Y(jié)構(gòu)和抗原表位預(yù)測不精確等原因所引起。因此,后續(xù)研究將考慮采取其他方式對EVM0008813多肽抗體進行特異性檢測。
4 結(jié)論
制備獲得的斑馬魚zgc113227多克隆抗體和卵形鯧鲹EVM0008813多肽抗體純度高、抗體效價高、特異性良好,可為斑馬魚zgc113227基因和卵形鯧鲹EVM0008813基因功能研究提供有利工具。
