小麥AP2/ERF轉錄因子家族生物信息學分析
2022-10-15 08:01:02卞云迪王雪晴楊晨曉王光鈺劉曉穎王振英
天津師范大學學報(自然科學版) 2022年4期
關鍵詞:分析
卞云迪,張 馳,王雪晴,楊晨曉,王光鈺,劉曉穎,王 穎,方 芳,王振英
(1.天津師范大學生命科學學院,天津 300387;2.天津市動植物抗性重點實驗室,天津 300387;3.天津市寶坻區林業發展服務中心,天津 301899)
植物在適應各種逆境脅迫的過程中進化出了復雜的調控網絡系統[1],其中,轉錄因子可以通過識別不同的順式作用元件來調控下游一系列相關基因的表達,進而提高植物的抗逆性[2].APETALA2/乙烯響應因子(APETALA2/ethylene-responsive factor,AP2/ERF)是植物特有的轉錄因子,一般包含一個或多個AP2保守結構域[3],該家族在植物的生長發育、生物和非生物逆境響應等過程中發揮了極其關鍵的調控作用[4-7].AP2/ERF超家族可以分為DREB(dehydration-responsive element binding protein)、ERF(ethylene-responsive element binding factors)、AP2(AP-ETALA 2)、RAV(related to ABI3/VP1)以及Soloist 5個亞族[8].隨著測序技術的發展,在多種植物中鑒定出了AP2/ERF家族成員[9-13].小麥基因組約為16 Gb,重復序列達85%.目前,高質量的中國春小麥基因組序列已釋放(http://www.wheat genome.org/)[14],但關于小麥TaAP2/ERF轉錄因子家族生物信息學方面的研究報道相對較少.本研究利用小麥全基因組數據庫信息,在全基因組范圍內鑒定小麥AP2/ERF轉錄因子家族成員,分析該家族成員的基因結構、順式作用元件及其可能調控的下游基因,為小麥轉錄因子的功能研究和利用提供參考.
1 材料與方法
1.1 TaAP2/ERF基因家族成員鑒定與序列特征分析
從Ensembl Plants數據庫(http://plants.ensembl.org/index.html)下載小麥的全基因組數據、開放閱讀框、染色體定位信息等.從Pfam數據庫(http://pfam.xfam.org)下載AP2/ERF基因特有的保守結構域模型(PF09425和PF06200),利用該模型在小麥蛋白數據庫Blastp中進行比對,得到候選基因,將其提交到Hmmer、NCBI-CDD等在線網站進行保守域的結構分析,最終得到TaAP2/ERF基因.利用在線工具ExPASy(http://web.expasy.org/protparam/)預測TaAP2/ERF基因編碼蛋白的等電點(pI)、相對分子質量、氨基酸長度和染色體定位信息等屬性,利用CELLO V.2.5(http://cello.life.nctu.edu.tw/)在線網站進行亞細胞定位分析.
1.2 TaAP2/ERF基因家族成員保守motif和基因結構預測
利用在線數據庫MEME(http://meme-suite.org/)預測TaAP2/ERF基因家族成員的保守結構域.按照WebLogo(http://weblogo.berkeley.edu/)默認設置獲取序列標識進行基因結構分析.
1.3 順式作用元件分析作圖
選取TaAP2/ERF基因上游1 500 bp基因序列,利用PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)進行啟動子的順式作用元件分析.把具有類似功能的啟動子元件歸為一類,再用TBtools軟件匯總歸類.
1.4 TaAP2/ERF家族基因表達分析
利用在線數據庫expVIP(http://www.wheat-expression.com/)分析TaAP2/ERF家族各基因的表達量,分析基因在不同生物/非生物脅迫下的表達水平,繪制基因表達熱圖.
2 結果與分析
2.1 TaAP2/ERF基因家族成員的理化性質
以小麥全基因組數據為基礎,共鑒定出74個TaAP2/ERF基因,根據基因的同源關系進行命名.利用ExPASY網站分析TaAP2/ERF基因編碼蛋白質的基本理化性質,具體如表1所示.預測TaAP2/ERF基因家族編碼蛋白質在180~484個氨基酸之間,相對分子質量為18.77×103~51.07×103,等電點為4.62~10.79.TaAP2/ERF家族編碼的氨基酸多數為酸性,其中47個蛋白的等電點小于7,偏酸性;其余27個蛋白的等電點大于7,偏堿性.利用CELLO在線預測亞細胞定位,發現53個TaAP2/ERF蛋白定位在細胞核中,這與多數轉錄因子的報道相一致;而定位在其他部位的蛋白則需通過實驗確定其功能.

表1 小麥AP2/ERF轉錄因子家族基因的基本信息Tab.1 Basic information of AP2/ERF transcription family genes in wheat

續表
2.2 TaAP2/ERF基因家族成員的保守motif和基因結構特征
對小麥的74個TaAP2/ERF蛋白序列進行分析,結果如圖1所示.TaAP2/ERF基因家族共有10個保守基序,利用TBtools軟件結合進化樹對得到的結果進行可視化分析(圖1(b)),結果發現,鑒定出的TaAP2/ERF蛋白中保守基序的數量和類型略有不同.所有蛋白均含有Motif1和Motif3,這2個結構共同組成了較為保守的AP2/ERF結構域,大多數的保守結構域位于蛋白N端,表明這些Motif對于TaAP2/ERF蛋白行使功能特別重要.相鄰蛋白之間的Motif分布雖然有較高相似性,但在保守基序的種類、數量以及分布上存在差異,暗示TaAP2/ERF家族蛋白各亞家族成員之間可能存在功能上的分化,而保守基序的功能尚待研究.
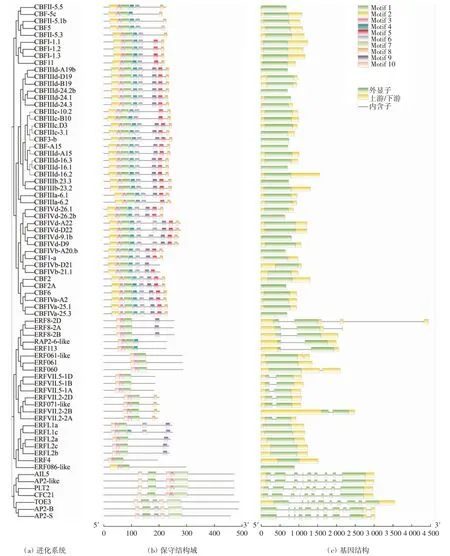
圖1 小麥TaAP2/ERF基因的系統進化、保守結構域和基因結構Fig.1 Phylogenetic evolution of TaAP2/ERF genes,conservative structure domain and genetic structure
對TaAP2/ERF家族成員結構進行分析,繪制基因結構示意圖,如圖1(c)所示.在鑒定到的74個TaAP2/ERF基因中,外顯子數量從0到4個不等,內含子數量從0到10個不等.少數家族成員含有多個內含子插入,如AP2-B含有10個外顯子和9個內含子;CBF亞家族成員沒有內含子,如CBF-5C、CBF-2A、CBF11等.由此可見,不同分類群間內含子的數量差異較大,相同亞族中內含子數量的差異較小,即具有相似的結構,如ERF061-like、ERF071-like等.TaAP2/ERF成員之間的內含子長度、外顯子位置和數目存在差異,說明小麥TaAP2/ERF家族在進化過程中產生了較強的分化,可能也是導致其成員功能不同的原因之一.
2.3 TaAP2/ERF基因家族成員的順式響應元件分類特點
選取小麥TaAP2/ERF基因上游1 500 bp的基因組序列,分析小麥TaAP2/ERF基因的順式作用元件,整理出與激素和非生物脅迫相關的順式作用元件,結果如圖2所示.
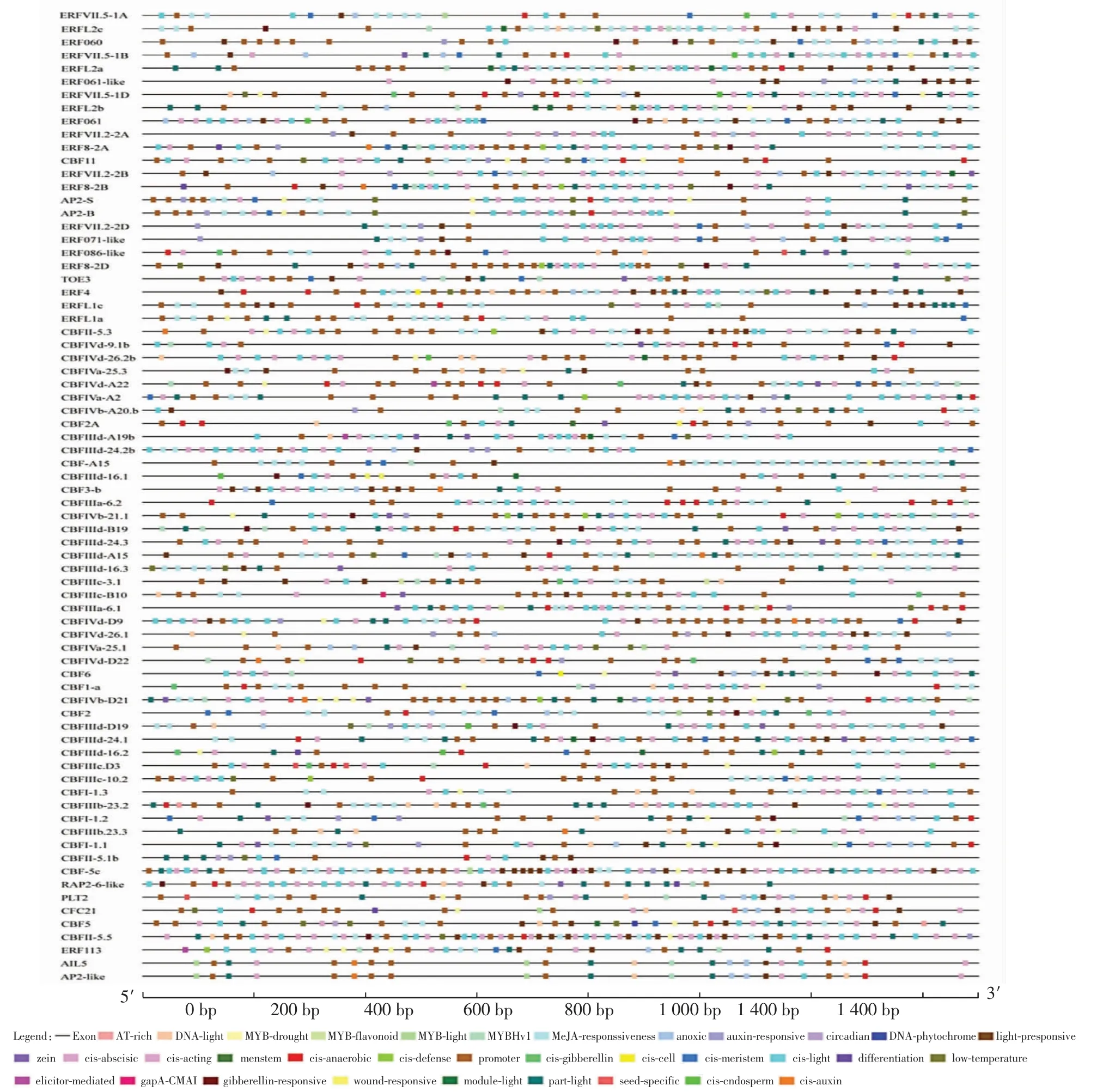
圖2 小麥TaAP2/ERF基因的激素和非生物脅迫響應相關的順式作用元件分析Fig.2 Analysis of cis-acting elements related to hormone and abiotic stress response of wheat TaAP2/ERF genes
由圖2可知,TaAP2/ERF啟動子區域包含多種應答元件,包括光、干旱、激素等逆境相關的應答元件.另外,還發現了厭氧誘導、創傷響應、晝夜節律調控、細胞周期調控、低溫響應的應答元件.其中,光應答元件分布在所有TaAP2/ERF基因家族成員中,推測光在誘導TaAP2/ERF表達調控中可能發揮著重要作用.激素響應元件分布也較為廣泛,如48.64%的家族成員啟動子序列中存在生長素響應元件,44.59%存在赤霉素響應元件,83.78%存在茉莉酸甲酯響應元件,91.89%存在脫落酸響應元件,這說明激素與TaAP2/ERF的表達有密切關系.與非生物逆境脅迫相關的元件有3種,如83.78%的家族成員具有厭氧誘導響應元件,48.64%的家族成員含有干旱誘導的響應元件,43.24%的家族成員含有低溫響應元件.除鑒定到參與干旱、厭氧、低溫等逆境損傷的應答元件外,還鑒定到了參與創傷的響應元件(CBF2A、CBFIIId-16.1)、參與晝夜節律調控的響應元件(ERFVII.2-2B、CBFIIId-B19、CBFIVd-26.1、CBFII-5.1b、CBF5).這 些 結 果 表 明,TaAP2/ERF基因可能在植物光合作用、逆境脅迫響應以及激素調控途徑中發揮了重要作用.
2.4 TaAP2/ERF家族基因在不同發育時期和生長條件下的表達分析
利用expVIP數據庫分析TaAP2/ERF家族各基因的表達,部分基因的熱圖分析如圖3所示.
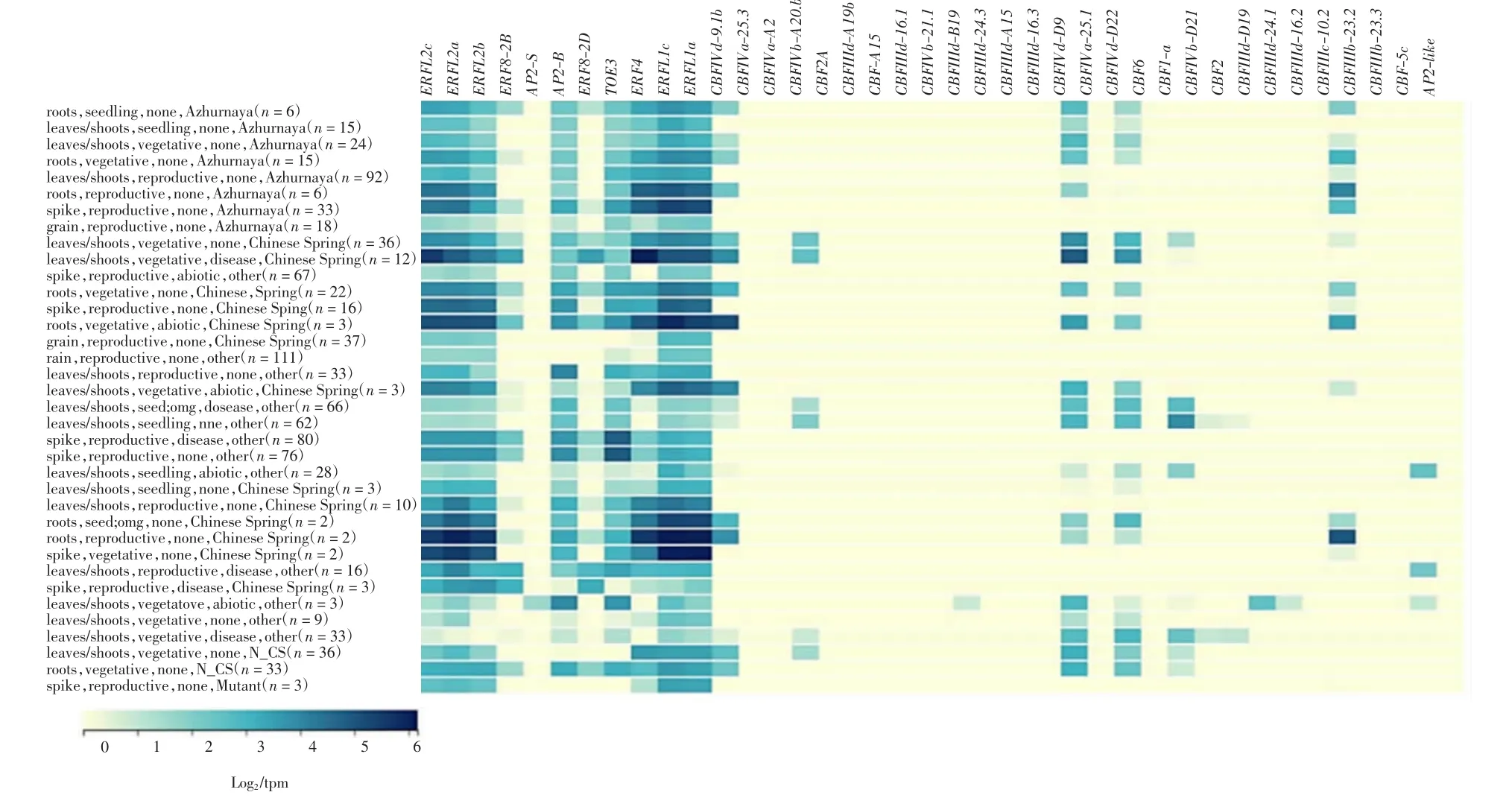
圖3 小麥部分TaAP2/ERF基因的熱圖分析Fig.3 Heat map analysis of partial TaAP2/ERF genes of wheat
由圖3可以看出,不同生長發育時期和生長條件下,各基因的表達水平呈現多樣化:ERFL2c、ERFL2a、ERFL1c和ERFL1a基因在所有條件下均保持較高表達水平;CBFIVa-25.3等16個基因在所有條件下均未檢測到表達;CBFIVd-D9等16個基因在生物或非生物脅迫下的表達量呈現波動,其中,AP2-S、CBFIIId-B19、CBFIIId-D19和CBFIIId-24.1響應非生物脅迫的基因在小麥葉片和根中表達上調,CBFIVb-D21和CBF2基因在病原菌侵染后的幼苗期葉片中平均表達量下調,營養生長期平均表達量上調.雖然不同生長時期的CBFIVb-D21和CBF2基因表達模式不同,但二者均能夠對病原菌侵染做出響應.
3 討論與結論
隨著水稻、玉米、小麥等重要農作物基因組數據的釋放,利用生物信息學技術分析基因家族序列的特征和進化關系,為作物基因功能的研究提供了很多幫助[15-16].AP2/ERF轉錄因子通常參與植物生長發育、細胞周期調控及逆境脅迫響應等重要生物過程[17].本研究對小麥基因組中的TaAP2/ERF轉錄因子進行了全基因組鑒定,對家族成員進行了系統的生物信息學分析.以中國春小麥基因組信息為基礎,共鑒定到74個小麥TaAP2/ERF轉錄因子基因,不規則分布在1~7染色體上.與多數轉錄因子的亞細胞定位情況一樣,大部分TaAP2/ERF蛋白位于細胞核內,少部分存在于葉綠體、細胞質和細胞膜,表明TaAP2/ERF蛋白的多個成員可能在小麥的生長發育過程中行使不同功能.TaAP2/ERF蛋白的等電點范圍為4.62~10.79,超過63%的TaAP2/ERF蛋白為酸性蛋白.進化分析結果表明,TaAP2/ERF轉錄因子家族保守基序較多且較為復雜,基因結構相似的轉錄因子親緣關系相近,如CBFIIIc-B10和CBFIIId-D19、ERFL1a和ERFL2a、AP2-B和AP2-S等.TaAP2/ERF轉錄因子的順式作用元件分析結果表明,所有TaAP2/ERF基因家族成員的啟動子區均含有光響應元件和不同的激素響應元件(如赤霉素和生長素順式作用元件[18-19])、非生物脅迫響應順式作用元件(如干旱脅迫和低溫脅迫響應元件等[20]).這說明TaAP2/ERF家族基因通過自身順式作用元件的差異響應小麥的多個生長發育過程.進一步分析TaAP2/ERF家族基因在生物和非生物脅迫下的表達量變化,結果發現,AP2-S、CBFIIId-B19、CBFIIId-D19和CBFIIId-24.1在非生物脅迫下發生特異表達,且CBFIIId-B19、CBFIIId-D19和CBFIIId-24.1這3個基因結構相同,推測CBFIIId亞家族基因可能在小麥響應非生物脅迫的過程中發揮了重要作用;CBFIVb-D21和CBF2為病原菌侵染后的特異表達基因,基因結構相似度也很高.因此,可將這些基因作為后續研究的切入點,分析TaAP2/ERF在小麥響應生物和非生物脅迫中的功能.
綜上所述,本研究利用多個生物學網站和軟件系統,分析了小麥TaAP2/ERF基因家族的定位、基因結構、順式作用元件以及表達模式,篩選獲得6個在生物脅迫或非生物脅迫下特異表達的基因作為備選基因,為后續研究基因功能及其在小麥生長發育和脅迫應答過程中的調控機制提供參考.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
