牛乳乳清中主要過敏原的B細胞表位定位
2022-10-31 08:56:22胡永芯譚宏凱熊子奕袁娟麗潘麗娜汪家琦
食品科學 2022年20期
關鍵詞:血清
胡永芯,譚宏凱,胡 巍,熊子奕,袁娟麗,潘麗娜,汪家琦,李 欣,
(1.南昌大學 食品科學與技術國家重點實驗室,江西 南昌 330047;2.南昌大學食品學院,江西 南昌 330047;3.南昌大學藥學院,江西 南昌 330047;4.澳優(yōu)乳業(yè)(中國)有限公司,湖南 長沙 410005)
牛乳及乳制品因含有較高的營養(yǎng)價值而備受廣大消費者的青睞,但牛乳過敏引發(fā)的食品安全問題也被人們廣泛關注。牛乳及牛乳制品是世界衛(wèi)生組織與聯(lián)合國糧食及農業(yè)組織認定的八大類主要過敏食物之一。牛奶蛋白過敏(cow’s milk allergy,CMA)是生命早期的一種常見疾病,患病率為2%~7%,且呈現(xiàn)增加的趨勢。據(jù)估計,歐洲國家2%~3%的嬰兒會出現(xiàn)牛乳過敏癥狀。在一項針對6 768 名中國嬰兒的最新調查中,發(fā)現(xiàn)CMA患病率為2.69%。CMA分為免疫球蛋白E(immunoglobulin E,IgE)介導、非IgE介導和混合(IgE與非IgE)介導。大約60%的CMA是IgE介導,即與體液免疫反應異常有關,盡管因研究人群和年齡的不同而略有差異。非IgE介導或兩者混合介導的CMA相對少見。IgE介導的CMA發(fā)生機制是牛乳過敏原致敏機體免疫細胞并使之產生特異性IgE(specific IgE,sIgE),隨后sIgE與效應細胞結合使其處于致敏階段;機體再次接觸到同種過敏原會導致效應細胞脫顆粒和釋放介質,如組胺等,最終導致皮膚系統(tǒng)、呼吸系統(tǒng)、胃腸道系統(tǒng)等出現(xiàn)臨床癥狀,如濕疹、呼吸困難、腹痛、嘔吐等,牛乳過敏嚴重時甚至會導致患者休克和死亡。
牛乳中含有30多種蛋白質,都具有潛在致敏性。目前認為酪蛋白、-乳球蛋白(-lactoglobulin,BLG)和-乳白蛋白(-lactalbumin,ALA)是主要過敏原。在20 ℃條件下調節(jié)脫脂乳pH值至4.6,可得到兩部分:其中凝固狀態(tài)的酪蛋白占80%,溶液狀態(tài)的乳清蛋白占20%。乳清蛋白中主要過敏原是BLG和ALA,分別占乳清蛋白組分的50%和25%。BLG是一種屬于脂鈣蛋白超家族的具有162 個氨基酸殘基的視黃醇結合蛋白,在天然狀態(tài)下以36 kDa二聚體的形式存在,它具有2 個二硫鍵和1 個游離半胱氨酸。ALA是一種屬于溶菌酶家族的具有123 個氨基酸殘基的單體球狀鈣結合金屬蛋白,分子質量為14.4 kDa,具有4 個二硫鍵和1 個鈣的高親和力結合位點。有研究發(fā)現(xiàn)大約82%的牛乳過敏患者對BLG過敏,大約75%的牛乳過敏患者可對多種牛乳蛋白過敏。
表位是過敏反應的物質基礎,可分為線性表位和構象性表位,前者由氨基酸殘基線性排列組成,后者是氨基酸殘基在空間相互靠近形成的能被免疫活性物質識別的特定三維結構。表位也可以按照表位受體分為T細胞表位與B細胞表位,B細胞表位指的是B細胞受體或B細胞分泌的特異性抗體識別的表位,如IgE表位和IgG表位。B細胞線性表位的長度通常有8~15 個氨基酸。盡管有報道稱線性表位可短至5 個氨基酸,但能與IgE高活性結合的表位至少含有8 個氨基酸。近年來,肽陣列、噬菌體展示、X射線晶體學等技術都已經運用于食物過敏原表位的定位。噬菌體展示技術是一種基于噬菌體表面展示的目標多肽或蛋白進行篩選的體外選擇系統(tǒng),能實現(xiàn)抗體基因型和表現(xiàn)型的統(tǒng)一,由于其便捷、特異性強、操作方便等特點,被廣泛用于表位定位。
通過各種物理方法、化學方法、生物方法破壞牛乳蛋白表位能有效降低牛乳蛋白致敏性,其中酶法水解乳蛋白已經被廣泛應用于低致敏配方奶粉的制備,也是預防和緩解嬰幼兒牛乳過敏的重要途徑。市面上的牛乳低致敏水解配方奶粉主要通過酶解酪蛋白或乳清蛋白獲得。理論而言,基于乳清蛋白主要過敏原表位進行定向酶解能有效降低致敏性,將是低致敏乳蛋白研發(fā)的新趨勢。本研究瞄準牛乳乳清中主要過敏原,即ALA和BLG,進行表位定位,采用酶聯(lián)免疫吸附法(enzyme linked immunosorbent assay,ELISA)篩選16 個牛乳過敏兒童患者的血清,以Hitrap Protein G HP親和層析柱純化的高純度牛乳特異性抗體為靶分子,對噬菌體隨機十二肽庫進行親和淘選,并利用生物信息學進行預測,最后獲得ALA和BLG的表位信息。本研究獲得的表位數(shù)據(jù)可用于指導定向打斷表位的蛋白酶篩選和基于表位的低過敏性乳清蛋白制品開發(fā)。
1 材料與方法
1.1 材料與試劑
BLG、ALA、明膠、生物素標記羊抗人IgE、辣根過氧化物酶(horseradish peroxidase,HRP)標記羊抗人IgG美國Sigma-Aldrich公司;Ph.D.-12噬菌體試劑盒英國Biolabs公司;BCA蛋白定量試劑盒 上海碧云天生物技術有限公司;96 孔酶標板 深圳金燦華實業(yè)有限公司;HRP標記的親和素、3,3′,5,5′-四甲基聯(lián)苯胺(3,3′,5,5′-tetramethylbenzidine,TMB)底物溶液北京欣博盛生物科技有限公司;Protein G HP預裝柱美國GE公司;預染蛋白Marker 美國Thermo公司;5×上樣緩沖液(含-巰基乙醇)、0.45 μm 濾膜北京索萊寶科技有限公司;HRP酶標記的抗M13二抗北京義翹神州科技股份有限公司;其他試劑均為國產分析純。
1.2 儀器與設備
微量蛋白測定儀、PowerPac 3000電泳儀、Mode 1860酶聯(lián)免疫檢測儀、Quantity One凝膠成像系統(tǒng)美國Bio-Rad公司;Amicon Ultra-15超濾離心管、超純水生成器 美國Millipore公司;高速冷凍離心機 美國Thermo公司;pH計 德國賽多利斯集團;ME104型電子天平 梅特勒托利多儀器上海有限公司;紫外檢測儀上海青浦滬西儀器廠。
1.3 方法
1.3.1 牛乳過敏患者血清池的構建
本實驗使用的過敏患者血清由北京協(xié)和醫(yī)院惠贈。過敏患者血清中針對牛乳蛋白的sIgE水平由ImmunoCAP檢測,選擇牛乳蛋白sIgE水平大于等于0.35 kU/L并且年齡在3~12 歲的血清進行ELISA。參考李柳栩的方法并作出適當調整,利用間接ELISA法測定牛乳過敏患者血清中抗ALA特異性免疫球蛋白E(-lactalbumin specific immunoglobulin E,ALA-sIgE)、抗ALA特異性免疫球蛋白G(-lactalbumin specific immunoglobulin G,ALAsIgG)、抗BLG特異性免疫球蛋白E(-lactoglobulin specific immunoglobulin E,BLG-sIgE)、抗BLG特異性免疫球蛋白G(-lactoglobulin specific immunoglobulin G,BLG-sIgG)水平。包被液為pH 9.6的0.05 mol/L碳酸鹽緩沖液,洗液、封阻液、血清、二抗等均使用0.01 mol/L磷酸鹽緩沖液(phosphate buffered saline,PBS),洗液含有0.05%(/)Tween-20,封阻液含有3%明膠。使用包被液將ALA或BLG稀釋到1 μg/mL,加入到96 孔酶標板,100 μL/孔,4 ℃過夜。隔日取出酶標板并扣干,洗板3 次,每次5 min。加入封阻液,250 μL/孔,37 ℃保溫保濕1 h。取出酶標板扣干后洗滌3 次。加入稀釋好的過敏患者血清,100 μL/孔,37 ℃保溫保濕1 h。測定IgG水平時,血清稀釋倍數(shù)為2 000 倍。測定IgE水平時,血清稀釋倍數(shù)為200 倍。取出酶標板扣干后洗滌3 次,加入1∶5 000稀釋的HRP標記羊抗人IgG或生物素標記羊抗人IgE,100 μL/孔,37 ℃保溫保濕1 h。對于IgG板,二抗孵育結束后,直接取出酶標板扣干后洗滌3 次,加入TMB底物溶液,100 μL/孔,37 ℃保溫保濕15 min。加入2 mol/L HSO溶液終止反應,50 μL/孔。測定450 nm波長處的吸光度。對于IgE板,二抗孵育結束后,還需額外加入HRP標記的親和素,具體步驟為:取出酶標板扣干后洗滌3 次,加入1∶60稀釋的HRP標記的親和素,100 μL/孔,37 ℃保溫保濕1 h。后續(xù)顯色步驟與IgG板保持一致。
以/>2且>0.2時血清判斷為陽性,其中和分別為樣品與陰性血清的OD值。根據(jù)間接ELISA結果,篩選出ALA-sIgE、BLG-sIgE、ALA-sIgG、BLG-sIgG水平均呈現(xiàn)陽性的患者血清混合血清池。
1.3.2 特異性抗體的純化
參考袁水林方法使用Protein G柱進行牛乳蛋白抗體的純化,并加以改進。先后采用200 μL活化液(1 mol/L Tris-HCl,pH 9.0)和10 倍柱體積的平衡緩沖液(20 mmol/L PBS,pH 7.4)沖洗親和柱,流速均為1 mL/min。采用平衡緩沖液以1∶1(/)的比例稀釋血清后通過0.45 μm水系濾膜。過濾后的血清加入親和柱后,將親和柱上下的蓋子旋緊,37 ℃搖動結合1 h。以平衡緩沖液和洗脫液(0.1 mol/L 甘氨酸-HCl,pH 2.7)分別對親和柱進行非特異性和特異性洗脫,流速均為1 mL/min,收集流出液200 μL/管。在收集特異性洗脫峰的過程中,加入中和液(1 mol/L Tris-HCl,pH 9.0)到收集管,標準是每毫升特異性洗脫液中加入60~200 μL中和液,直到pH值達到7左右。特異性洗脫結束之后,分別用10 倍柱體積的平衡緩沖液和20%乙醇溶液洗滌親和柱。收集到的非特異性洗脫液與特異性洗脫液用超濾離心管濃縮并更換緩沖液為0.01 mol/L PBS。采用還原型聚十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)鑒定純化的抗體,采用BCA試劑盒鑒定濃度。將獲得的抗體于-20 ℃冰箱保存。參考1.3.1節(jié)進行純化抗體的IgE和IgG水平測定,作出如下調整:IgE與IgG水平測定時,樣品(濃縮非特異性洗脫液與特異性洗脫液)的稀釋倍數(shù)分別是20 倍和200 倍。
1.3.3 噬菌體展示技術篩選IgE與IgG表位
參考NEB噬菌體試劑盒說明書,利用噬菌體隨機肽庫進行IgE與IgG模擬表位的淘選。第1~4輪,包被的IgE質量濃度為600、450、300、300 μg/mL,包被的IgG質量濃度分別為100、75、50、50 μg/mL,洗液體積分數(shù)分別為0.1%、0.25%、0.5%、0.5%,噬菌體孵育結合時間分別為60、45、30、30 min,甘氨酸洗脫時間分別為6、8、10、10 min。逐步嚴格的淘選條件有助于選擇到特異性更強的噬菌體序列。從IgE或IgG組第4輪測滴度的平板上挑取各10 株噬菌體送往金唯智公司進行測序,測序引物為-96III。
1.3.4 間接ELISA鑒定陽性噬菌體克隆
通過噬菌體ELISA 檢測所選多肽對抗體的結合。按照噬菌體試劑盒說明書,并基于實驗方案作出以下調整:包被蛋白是2 μg/mL的抗牛乳蛋白IgE(IgG),100 μL/孔,包被于96 孔微孔板上,4 ℃孵育過夜,同時包被3% BSA-PBS作陰性對照。傾去孔內液體,用含0.05%(/)Tween-20的PBS洗滌3 次。用3%的明膠封阻,300 μL/孔,37 ℃孵育1 h。洗板3 次,加入經擴增純化的噬菌體,100 μL/孔,37 ℃孵育1 h。加入的噬菌體數(shù)量有4 組梯度,分別約為10、10、10、10個/孔,陰性對照加入噬菌體量為10個/孔,空白對照加PBS。洗板6 次,加入HRP標記的抗M13二抗,100 μL/孔,37 ℃孵育1 h。洗板6 次,加入TMB顯色液,100 μL/孔,37 ℃避光顯色15 min。加入2 mol/L HSO溶液終止反應,50 μL/孔,測定OD值。/>2且>0.2時,噬菌體判斷為陽性,其中和分別為樣品與陰性對照的OD值。
1.4 數(shù)據(jù)處理
用于序列分析的ALA和BLG的氨基酸序列從NCBI數(shù)據(jù)庫(www.ncbi.nlm.nih.gov)獲取。使用DNAStar的Editseq軟件模塊對插入的噬菌體外源基因序列進行翻譯。分別使用DNAman 7.0軟件和The Pepitope Server網絡服務器(http://pepitope.tau.ac.il/)進行線性表位與構象性模擬表位的定位。采用GraphPad Prism 8.0.1處理ELISA數(shù)據(jù),使用檢驗方法評價準確性,<0.05,差異顯著。
2 結果與分析
2.1 牛乳過敏患者血清信息
牛乳過敏患者血清信息見表1。16 名患者包括9 名男性7 名女性,患者年齡的平均值為4.4 歲,6 歲以上兒童僅有4 名,可能原因是牛奶過敏在6 歲后消退。患者的臨床癥狀表現(xiàn)為濕疹、特異性皮炎、食物過敏反應等,其中食物過敏包括皮膚系統(tǒng)癥狀(風團、瘙癢、充血等)、胃腸道系統(tǒng)癥狀(腹痛、腹瀉、惡心、嘔吐等)、呼吸系統(tǒng)癥狀(咳嗽、喘息、胸悶、呼吸困難等),以及嚴重過敏反應(頭暈、暈厥、黑朦等)。ImmunoCAP是檢驗IgE水平的金標準,已經證實在牛奶、雞蛋和花生過敏的診斷上有很高的準確性。本實驗根據(jù)ImmunoCAP系統(tǒng)檢測牛乳蛋白sIgE抗體水平,高于0.35 kU/L閾值的血清被視為陽性。本實驗所用的患者sIgE值集中在1.0 kU/L附近,其中15號血清的sIgE值最高,超過100 kU/L。15號患者未顯示出較嚴重的過敏反應,說明sIgE水平與過敏癥狀無正相關關系。Anvari等報道sIgE水平高,過敏反應發(fā)生的幾率更高,這是值得注意的問題。
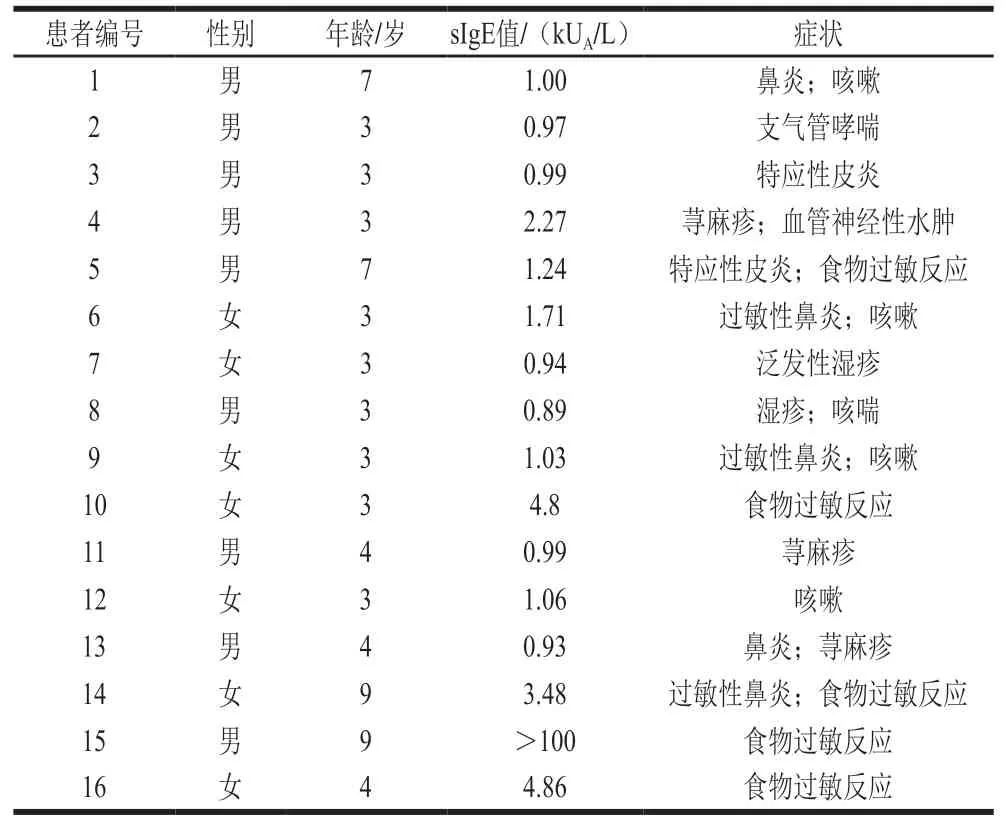
表1 牛乳過敏患者的特征Table 1 Characteristics of patients with cow milk allergy enrolled in this study
2.2 牛乳過敏患者ALA和BLG IgE水平測定
通過間接ELISA測定患者血清中針對乳清主要過敏原的特異性抗體水平。本研究中有75%(14份)牛乳過敏患者血清對BLG存在IgE陽性反應,有25%(4 份)患者對ALA存在IgE陽性反應。本實驗結果說明BLG確實是主要過敏原,同時,在此16 個牛乳過敏患者中,ALA可被認為是次要過敏原,這與Tsabouri等研究結果相似,他們獲得的牛乳患者對BLG和ALA過敏率分別為51%和19%。另外過敏患者的牛乳蛋白sIgE水平并未顯示出與性別等因素相關。從圖1還發(fā)現(xiàn),ImmunoCAP的牛乳蛋白sIgE結果與ELISA結果存在差異,因為前者是通過粗蛋白進行檢測,后者只檢測了某一種組分蛋白,而部分不可能代表整體,后者會遺漏部分特異性抗體的檢測。而對于sIgG,ALA和BLG具有相似的特異性識別,主要是1、3、4、7、12、13、14、15號血清,其中7號血清對ALA識別的顯著性差異最為明顯。
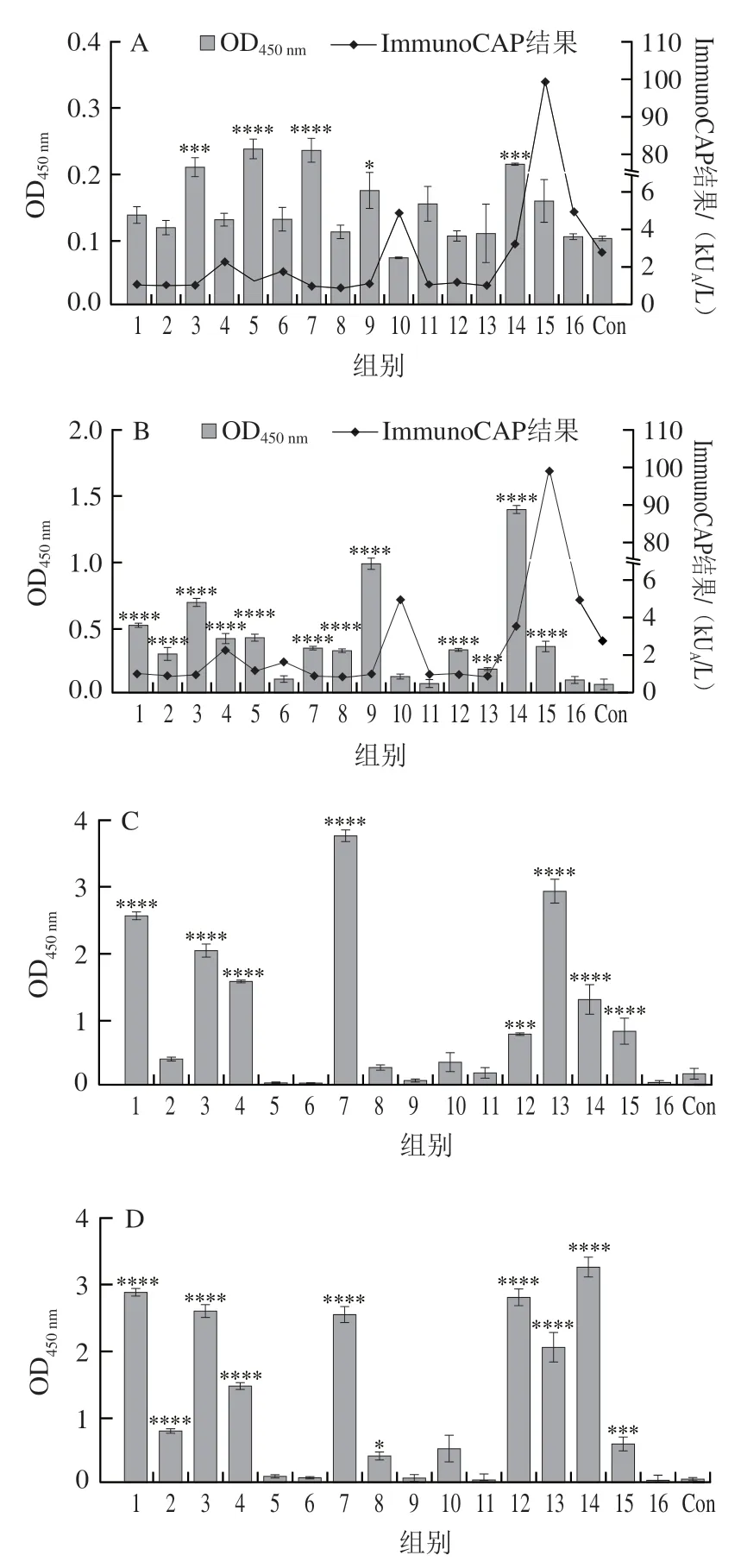
圖1 牛乳過敏患者特異性抗體水平Fig.1 Levels of specific antibodies in patients with milk allergy
為后續(xù)噬菌體展示技術篩選表位,需要挑選特異性抗體水平較高的血清制備血清池。血清5、6、9、11、16的ALA-sIgG和BLG-sIgG水平都偏低,6、10、11、16號血清的ALA-sIgE和BLG-sIgE水平較低,但血清5和9號的BLG-sIgE水平較高。綜上,6、10、11、16號不參與血清池構建,其余12 份混血清池。
2.3 抗牛乳蛋白抗體的親和純化與效價測定結果
采用蛋白G柱純化血清池的IgE與IgG抗體。鏈球菌G蛋白是鏈球菌菌株的細胞壁中的一種蛋白質,能在不改變IgG結構的前提下與其特異結合。與蛋白A柱相比,蛋白G柱具有親和力強,結合譜廣,結構穩(wěn)定等特點。
IgG由兩條相同的重鏈和兩條相同的輕鏈組成“Y”形結構,每條重鏈包含兩個不同的區(qū)域:可變區(qū)和恒定區(qū)。經Protein G柱純化后的IgG(圖2泳道3),在還原型電泳的結果中顯示為兩條蛋白條帶,分別位于50 kDa和25 kDa,僅含有少量的雜蛋白,主要集中在125 kDa左右。由圖2可知,未經任何處理的血清含有較多的雜蛋白(泳道1)。泳道2為非特異性洗脫液,在72 kDa附近有明顯的條帶,在50 kDa處沒有明顯條帶,說明其中的IgG已經被分離。經過Image J軟件計算,泳道2和泳道3的純度分別是90%和95.2%,因此認為獲得了沒有IgG抗體的非特異性洗脫液和高純度的抗牛乳IgG抗體。
純化前的血清池體積為200 μL,經過超濾濃縮更換緩沖液,將獲得的IgG保存于PBS中,最終各獲得600 μL的非特異性洗脫液和特異性洗脫液。BCA測定結果表明,非特異性洗脫液和特異性洗脫液的質量濃度分為為3 000 μg/mL和1 700 μg/mL。進行間接ELISA測定效價,由圖2B可知,特異性洗脫峰收集液中,ALA-sIgG和BLG-sIgG水平很高,而ALA-sIgE和BLG-sIgE水平很低,相反,非特異性洗脫液有較高的ALA-sIgE和BLG-sIgE水平。因此,認為蛋白G柱能有效從血清中分離IgG(特異性洗脫液)與IgE(非特異性洗脫液)。
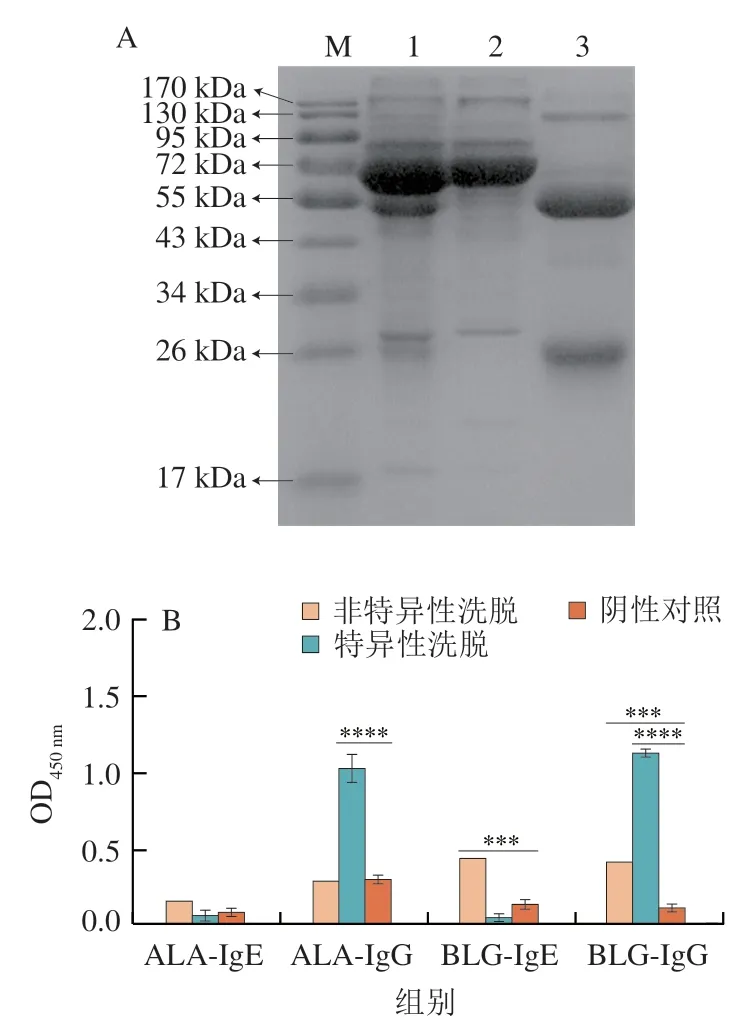
圖2 Protein G親和純化SDS-PAGE(A)和純化抗體的ELISA(B)Fig.2 SDS-PAGE patterns of proteins purified by Protein G affinity chromatography (A) and results of ELISA for purified antibodies (B)
2.4 親和淘選過程中噬菌體的富集
本實驗共進行了4 輪淘選,逐漸降低抗體(IgG與IgE)的包被量,增加洗液的Tween-20含量,噬菌體結合時間逐漸減少,洗脫時間逐漸延長,目的是增加淘選的嚴格程度以便篩選出特異性更強的噬菌體。表2展示了淘選過程中噬菌體的富集效果,可以發(fā)現(xiàn)每經過1 次淘選,噬菌體產出量和回收率都有所增加。在第4輪中,在IgE與IgG洗脫液中,每10 μL噬菌體的空斑形成單位滴度分別達到1.2×10PFU和1.3×10PFU,表明特異性結合的噬菌體在此過程中得到了有效富集。

表2 兒童IgE和IgG 4 輪親和淘選中噬菌體的富集Table 2 Phage enrichment in four rounds of affinity panning for IgE and IgG in children
2.5 噬菌體測序與噬菌體ELISA結果
第4輪的平板中,分別在IgG和IgE組中挑選10 個噬菌斑,分別命名為IgG1~IgG10、IgE1~IgE10,表3展示了噬菌體外源序列。送往金唯智公司進行測序并獲得19 個12肽隨機序列,另外一個克隆子未檢測到序列,可能是由于外源基因的丟失。在這19 個12肽中,有3 個序列出現(xiàn)了2 次,分別為VVGRAMAYSTIP、VSVPGIITGTLR、CFARTPSILMLA,其余序列均出現(xiàn)1 次。采用間接ELISA檢測所選多肽對抗體的結合能力。發(fā)現(xiàn)隨著加入噬菌體數(shù)量的降低,吸光度也呈現(xiàn)降低的趨勢。圖3展示了噬菌體加入量為10個/孔時的OD值。發(fā)現(xiàn)除了IgE5,其余克隆子的OD值全部高于陰性對照組的2 倍,因此18 個克隆子被判斷為陽性克隆。
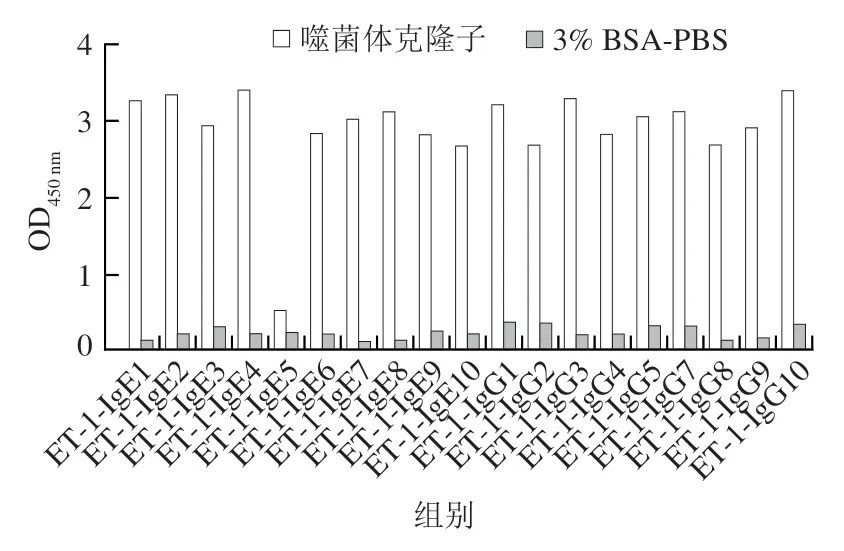
圖3 噬菌體克隆子的間接ELISA檢測結果Fig.3 Results of indirect ELISA for phage clones
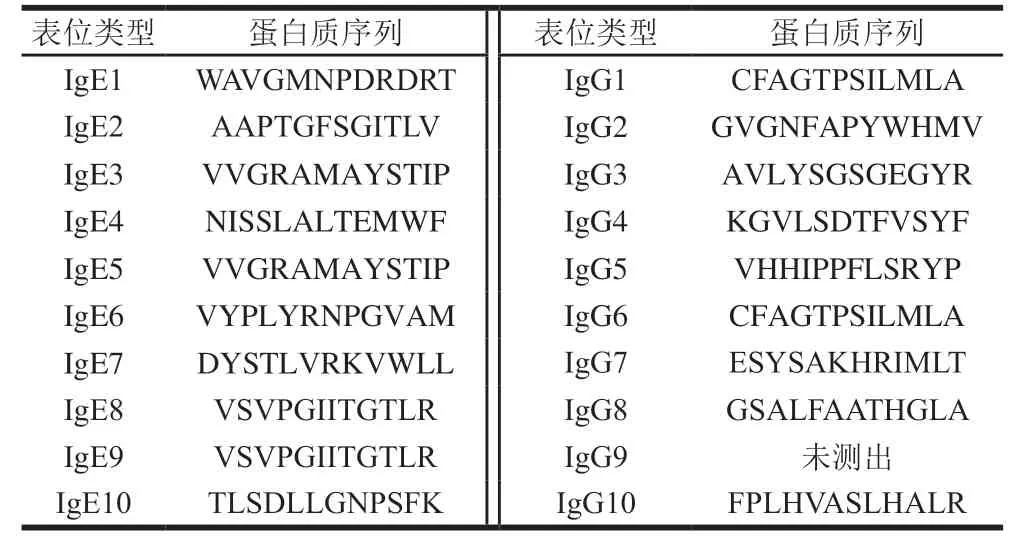
表3 兒童血清池模擬表位Table 3 Mimotopes panned in children’s serum pool
2.6 線性表位定位
使用軟件DNAman 7.0的多序列對比功能將噬菌體外源序列與ALA和BLG氨基酸序列進行對比,默認參數(shù)不變,分析肽段的識別位點與出現(xiàn)頻率,含有連續(xù)重合3 個氨基酸以上的肽序列和非連續(xù)重合5 個氨基酸以上的肽序列被定義為線性表位。由于雜蛋白不具有ALA和BLG特異性,所以假陽性序列只有極低的可能性與ALA或BLG序列的重合度較高。通過舍去多重對比結果中重合度較低的外源序列,能有效篩去噬菌體展示技術淘選到的可能結合雜蛋白的假陽性序列。圖4和圖5分別展示了ALA和BLG的線性表位。定位出ALA的IgE線性表位位于AA11~27,IgG線性表位位于AA17~29。BLG的IgE線性表位有3 個,分別為AA15~32、AA89~104、AA111~133,IgG線性表位為AA121~132。發(fā)現(xiàn)在測序結果中,重復出現(xiàn)兩次的VVGRAMAYSTIP和CFARTPSILMLA分別被定位為BLG的IgE和IgG線性表位,說明結合肽中的共有序列更可能是表位。從圖4和圖5可知,ALA的AA17~27區(qū)域和BLG的AA121~132區(qū)域能同時被IgE和IgG識別,可能是IgE和IgG的共有表位。研究表明,血清IgG4能阻止IgE與其受體的結合。IgG4抑制IgE活性可能有兩種機制:IgG4與IgE直接競爭結合過敏原或IgG4結合抑制性FcγRIIb。還有研究表明,能識別相同過敏原但亞類不同的抗體,即IgG1、IgG2、IgG3、IgA1和IgA2,能夠同等程度地阻斷嗜堿性粒細胞的激活。IgE和IgG共有表位的發(fā)現(xiàn)支持以上結論,即任何對過敏原具有足夠親和力的抗體都有可能通過與IgE競爭結合共有表位來有效防止IgE與其受體的交聯(lián)。

圖4 ALA的線性表位Fig.4 Linear epitopes of ALA

圖5 BLG的線性表位Fig.5 Linear epitopes of BLG
2.7 構象性表位定位
使用The Pepitope Server網絡服務器(http://pepitope.tau.ac.il/)進行構象性模擬表位的定位,考慮算法分值、值等參考因素,分別選擇前5 個評分最高的序列作為ALA和BLG的構象性模擬表位。表4、5分別展示了ALA和BLG構象性表位的氨基酸組成和評分,圖6、7分別展示了ALA和BLG的構象性表位的三維結構。發(fā)現(xiàn)ALA的構象性表位都位于-螺旋區(qū)域,這提示-螺旋結構在ALA構象性表位中非常重要。而BLG的多數(shù)構象性表位位于無規(guī)則區(qū)域,沒有在-折疊區(qū)域發(fā)現(xiàn)構象性表位,需要進一步深入探索和研究。

表4 ALA構象性表位Table 4 Conformational epitopes of ALA

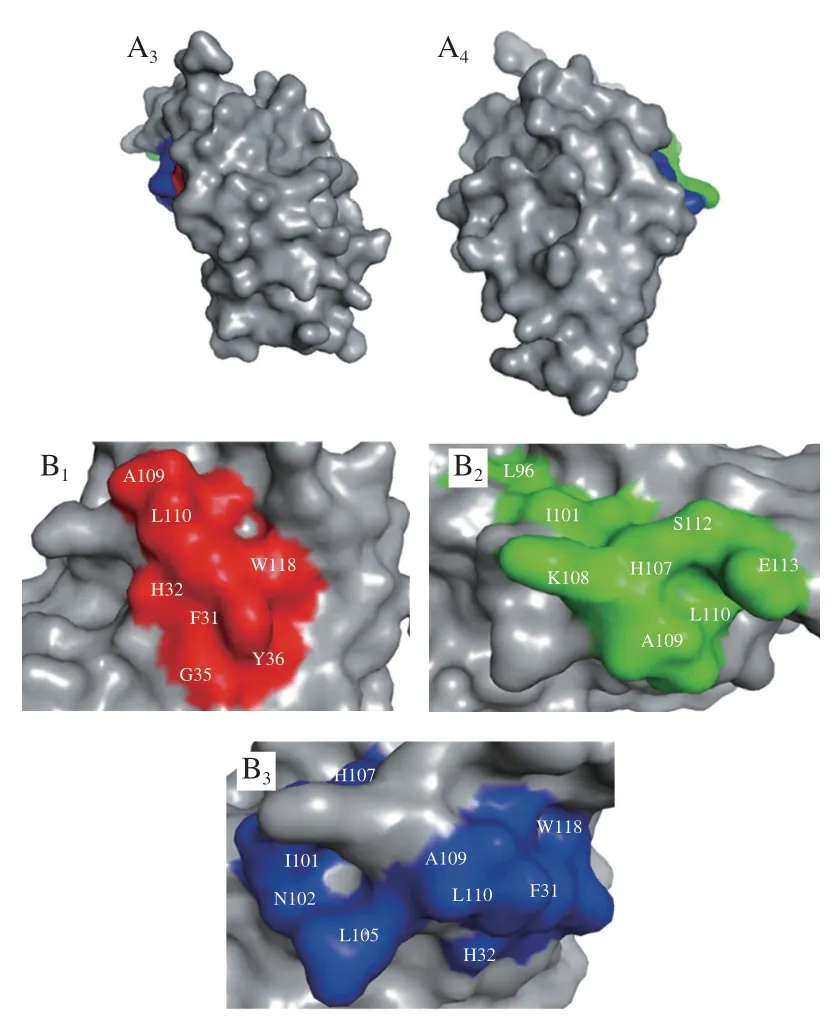
圖6 ALA的3 個構象性表位的分子表面(A)和構象性表位氨基酸組成(B)Fig.6 Molecular surface representation (A) and amino acid composition (B) of the three conformational epitopes of ALA
另外發(fā)現(xiàn)在ALA的構象性表位氨基酸組成中,L和A是3 個構象性表位的共有氨基酸,W和F能同時被IgG2和IgE4識別,H和I能同時被IgG7和IgE4識別,表明這些氨基酸可能在構象性表位的形成中發(fā)揮著至關重要的作用。ALA的3 個構象性表位的氨基酸組成相似,且在空間結構的位置較為緊密,集中在AA101~113,說明該區(qū)域很可能是ALA的關鍵表位。BLG的兩個構象性表位氨基酸組成不相似,空間位置不接近,表明牛乳過敏患者可能識別BLG的不同構象性表位。

表5 BLG構象性表位Table 5 Conformational epitopes of BLG
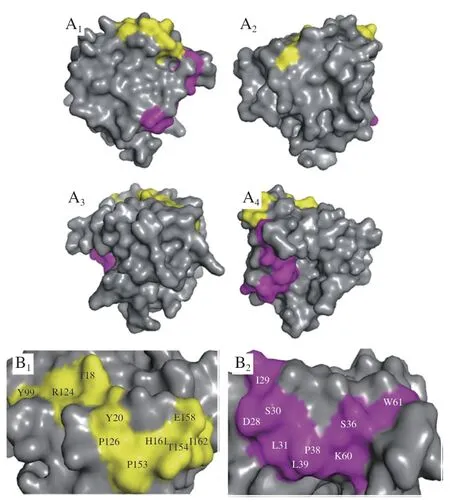
圖7 BLG的2 個構象性表位的分子表面(A)和構象性表位氨基酸組成(B)Fig.7 Molecular surface representation (A) and amino acid composition (B) of the two conformational epitopes of BLG
3 討論
牛乳過敏大多數(shù)是IgE介導的I型速發(fā)型超敏反應,一般在攝入牛乳24 h內出現(xiàn)過敏癥狀,通常基于臨床病史和牛乳蛋白sIgE水平或皮膚點刺實驗確診。除了過敏原sIgE,IgG在食物過敏中的作用同樣值得關注。IgG是血清中含量最多的免疫球蛋白(約占70%),IgG水平的升高與兒童牛奶過敏的自然消退呈正相關。在花生蛋白sIgE抗體檢測呈陽性的受試者中,Santos等發(fā)現(xiàn)更高水平的花生蛋白sIgG4與耐受性相關。IgG4也已經被確定為皮下過敏原免疫療法中非常強大的生物標志物。
眾所周知,X射線是一種精確的表位定位方法,但天然IgE抗體的缺乏限制了X射線在表位定位的廣泛應用。噬菌體展示技術可作為一種食物過敏原表位定位的替代方法,通過生物淘選噬菌體庫定位出線性與構象表位。袁水林等采用兩步親和層析法從8 名兒童中分離純化了抗ALA的特異性抗體,隨后,分別從Ph.D.-12和Ph.D.-C7C中淘選到ALA的線性表位與構象性表位。花生主要過敏原Ara h 1的IgE和IgG4表位也通過噬菌體展示隨機七肽庫的競爭性免疫淘選被鑒定,結果顯示每個患者都有自己獨特的IgE和IgG4表位識別譜,但一些重要的IgE表位對所有患者都是通用的。
本研究識別到ALA的IgE線性表位位于AA11~27,IgG線性表位位于AA17~29。前者與J?rvinenet等報道的IgE線性表位AA13~26完全重合,后者與叢艷君等識別到的IgG線性表位AA21~35完全重合,也識別到他們報道的關鍵氨基酸Pro。本研究與Li Xin等均采用噬菌體展示和二代測序相結合的方法篩選不同血清樣本之間共享的表位,但定位出的ALA線性表位并不一致,可能是血清的樣本差異導致識別不同表位圖譜。定位出BLG的IgE線性表位位于AA15~32、AA89~104、AA111~133,IgG表位為AA121~132。Cong Yanjun等的初始數(shù)據(jù)也顯示AA17~31、AA92~106是BLG的IgE線性表位,AA127~141是BLG的IgG表位。另一項對14 名牛乳過敏患者的研究表明,所有患者血清均識別AA95~113,而單個過敏患者血清顯示出與AA12~27及AA124~135的反應。本研究還發(fā)現(xiàn)一些IgE線性表位與IgG線性表位存在重疊,如ALA的AA17~27區(qū)域和BLG的AA121~132區(qū)域能同時被IgE和IgG識別,表明這些區(qū)域可能是IgE和IgG的共有表位。研究表明,非IgE抗體在過敏反應中的對IgE抑制作用可能是由于它們與IgE競爭結合過敏原上的共有表位,這些阻斷抗體是一種潛在的有價值的免疫治療抗體,可用于被動過敏原免疫療法。同時,表位重疊位置的共同氨基酸可能是檢測和診斷牛奶過敏的潛在有效信息標記物。
在本研究中,定位出ALA的2 個IgG構象性表位和1 個IgE構象性表位,BLG的1 個IgG構象性表位和1 個IgE構象性表位,其中ESLAKHIL與Li Xin等定位出的ALA的構象性LHKL高度吻合,表明L、H、L這3 個氨基酸可能是ALA構象性表位的關鍵組成氨基酸。同時,在本研究中,一些氨基酸能同時被IgE和IgG構象性識別,如ALA的L、A、W等,表明IgE和IgG的構象性表位存在一定相似性。研究還發(fā)現(xiàn)一些線性表位的殘基也出現(xiàn)在構象性表位中,如BLG的AA28~31,這些發(fā)現(xiàn)也支持了Ferdous等的觀點:在過敏反應中,超過90%的表位是構象表位,一些線性表位是構象表位的組成部分。
4 結論
通過噬菌體展示技術結合生物信息學成功定位出ALA和BLG的B細胞表位,包括線性與構象性表位。ALA的IgE線性表位為AA11~27;IgG線性表位為AA17~29;BLG的IgE線性表位為AA15~32、AA89~104、AA111~133;IgG 線性表位為AA121~132。研究發(fā)現(xiàn)IgE和IgG線性表位存在共同區(qū)域,表明對同種過敏原具有特異性的非IgE抗體可能通過與IgE競爭結合過敏原表位而發(fā)揮阻斷作用。同時,定位出ALA和BLG的構象性表位,并發(fā)現(xiàn)線性表位與構象性表位存在重疊區(qū)域,證明某些線性表位是構象性表位的一部分。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫(yī)科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫(yī)科大學學報(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(2021年1期)2021-01-26 00:56:02
昆明醫(yī)科大學學報(2020年12期)2021-01-26 00:44:04
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業(yè)科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫(yī)學院學報(2015年5期)2015-12-05 08:22:29
