三葉香茶菜提取物對肝星狀細胞和肝細胞TLR4/NF-κB/NLRP3通路的影響Δ
2022-10-31 16:06:50黃桂東吳瑞勝劉代華周至品廣西中醫藥大學藥學院南寧53000廣西醫科大學附屬柳州市人民醫院藥學部廣西壯族自治區衛生健康委員會廣西臨床疾病生物技術研究重點實驗室柳州市胃腸道中成藥工程技術研究中心廣西柳州545006
中國藥房 2022年20期
覃 樂,陳 勇,黃桂東,吳瑞勝,劉代華,周至品#(.廣西中醫藥大學藥學院,南寧 53000;.廣西醫科大學附屬柳州市人民醫院藥學部/廣西壯族自治區衛生健康委員會廣西臨床疾病生物技術研究重點實驗室/柳州市胃腸道中成藥工程技術研究中心,廣西柳州 545006)
三葉香茶菜Isodon ternifolius(D.Don)Kudo 為唇形科植物,又名細葉香茶菜,常用于治療黃疸、風濕腫痛等,其復方制劑可用于臨床治療急慢性肝炎[1-2]。有研究表明,三葉香茶菜乙醇提取物具有保肝、降酶活性,其主要藥效成分為齊墩果酸和熊果酸[3-4]。另有研究發現,三葉香茶菜的多種苯基木脂素成分可減少小鼠巨噬細胞RAW264.7 的炎性損傷,可能是其藥效成分之一[5]。動物實驗表明,三葉香茶菜能減輕四氯化碳致大鼠肝纖維化(hepatic fibrosis,HF)程度,下調Toll 樣受體4(Tolllike receptor 4,TLR4)、核因子κB(nuclear factor-κB,NF-κB)、磷酸化核因子κB 抑制蛋白(phosphorylated inhibitor of NF-κB,p-IκB)、NOD 樣受體蛋白3(NODlike receptor protein 3,NLRP3)的表達,減少白細胞介素1β(interleukin-1β,IL-1β)、IL-18、IL-6、腫瘤壞死因子α(tumor necrosis factor-α,TNF-α)等炎癥因子的釋放,提示其可通過抑制TLR4/NF-κB/NLRP3 通路來減輕大鼠的炎性損傷,從而發揮抗HF 作用[6-7]。有研究指出,NLRP3炎癥小體可誘導肝細胞死亡,并可在細胞外被肝星狀細胞吞噬,從而激活肝星狀細胞,使得IL-1β、α-平滑肌肌動蛋白(α-smooth muscle actin,α-SMA)分泌上調,進而加重HF[8],提示肝星狀細胞和肝細胞所構成的肝臟微環境在HF的發展過程中發揮了重要作用。基于此,本研究在前期體內實驗的基礎上,以TLR4/NF-κB/NLRP3通路為切入點,探討三葉香茶菜提取物對肝星狀細胞和肝細胞的影響,進一步揭示其抗HF的分子機制。
1 材料
1.1 主要儀器
本研究所用主要儀器包括CC-170T-8型三氣培養箱(新加坡ESCO 公司)、FilterMax F3 型多功能酶標儀[美谷分子儀器(上海)公司]、FC-1100型超微量核酸檢測儀(杭州遂真生物技術有限公司)、AriaMx型實時聚合酶鏈反應(real-time PCR)系統(美國Agilent 公司)、Tanon 5200型凝膠成像系統(上海天能科技有限公司)等。
1.2 主要藥品與試劑
三葉香茶菜飲片(批號20190501,產地廣西)于2019年10月購自廣西仙茱中藥科技有限公司,由廣西中醫藥大學藥用植物學教研室梁子寧教授鑒定為唇形科植物三葉香茶菜I.ternifolius(D.Don)Kudo的干燥根莖。
TAK-242 對照品(TLR4 阻斷劑,批號SML0832,純度≥98%)、秋水仙堿對照品(陽性對照,批號C3915,純度≥95%)均購自美國Sigma 公司;脂多糖(LPS,批號L8880)、MTT 細胞增殖及毒性檢測試劑盒(批號M1020)均購自北京索萊寶科技有限公司;ECL 化學發光底物(批號BL523A)購自北京蘭杰柯科技有限公司;IL-1β、IL-18、Ⅰ型膠原蛋白(type Ⅰcollagen,COL-Ⅰ)、COL-Ⅲ、丙氨酸轉氨酶(ALT)、天冬氨酸轉氨酶(AST)酶聯免疫吸附測定(ELISA)試劑盒(批號分別為JYM0419Ra、JYM0650Ra、JYM0413Ra、JYM0811Ra、JYM0722Ra、JYM0994Ra)均購自武漢基因美生物科技有限公司;α-SMA、轉化生長 因子β1(transforming growth factor-β1,TGF-β1)ELISA 試劑盒(批號分別為ml038078、ml002856)均購自上海酶聯生物科技有限公司;兔源TLR4、NF-κB p65、NLRP3、Gasdermin D(GSDMD)、凋亡相關微粒蛋白(apoptosis-associated speck-like protein containing CARD,ASC)、β-肌動蛋白(β-actin)單克隆抗體(批號分別為ab13867、ab76302、ab263899、ab219800、ab180799、ab8227)和辣根過氧化物酶標記的山羊抗兔IgG 二抗(批號ab6721)均購自英國Abcam 公司;兔源p-IκBα單克隆抗體(批號2859S)購自美國CST公司;其余試劑均為實驗室常用規格,水為純凈水。
1.3 實驗動物與細胞
SPF 級雄性SD 大鼠,體質量為180~200 g,購自湖南斯萊克景達實驗動物有限公司,動物生產許可證號為SCXK(湘)2019-0004。所有動物均飼養于廣西中醫藥大學實驗動物中心動物房(室內保持通風、干燥、潔凈)內,并自由攝食、飲水。動物處置符合相關動物實驗倫理要求。
2 方法
2.1 三葉香茶菜提取物的制備
取適量三葉香茶菜飲片,加10 倍量水浸泡30 min,煮沸60 min,用紗布過濾,收集濾液。同法重復提取3次,合并濾液,加95%乙醇適量使乙醇最終體積分數為70%,攪拌,靜置24 h后,過濾除渣,收集濾液,回收乙醇并濃縮成浸膏(每1 g 浸膏相當于生藥20 g),冷藏。將浸膏用DMEM 高糖培養基稀釋,過濾除菌后用于細胞實驗。
2.2 大鼠原代肝星狀細胞、原代肝細胞的分離
參照張玉等[9]所建方法,依次通過肝逆向兩步酶灌注法原位消化,密度梯度離心分離,特征性表面抗原α-SMA、結蛋白免疫熒光法鑒定,獲得原代肝星狀細胞。參照葉娟等[10]所建方法,依次通過改良原位兩步灌流法和多次過濾低速離心法分離,抗肝細胞角蛋白18免疫化學法鑒定,獲得原代肝細胞。所獲細胞均接種于完全培養基[含10%胎牛血清、青鏈霉素雙抗的DMEM 高糖培養基,下同]中,在37 ℃、5%CO2條件下培養(下同)。
2.3 三葉香茶菜提取物對肝星狀細胞及肝細胞增殖影響的檢測
取原代肝星狀細胞或肝細胞適量,按6×104個/mL接種于96孔板中,分別用不同質量濃度的三葉香茶菜提取物[肝星狀細胞:0.2、0.4、1、2、4 mg/mL,肝細胞:0.1、0.5、1、1.5、2.5 mg/mL,均按生藥量計(下同),質量濃度參考前期預實驗結果設置]干預,并設不含藥物的空白對照組。培養24 h 后,每孔加入5 mg/mL 的MTT 試劑20 μL,繼續培養4 h。使用酶標儀于570 nm波長處測定各孔的吸光度,計算增殖抑制率[增殖抑制率=(空白對照組吸光度-實驗組吸光度)/空白對照組吸光度×100%]和半數抑制濃度(IC50)。每質量濃度設3個復孔,實驗重復3次。
2.4 細胞分組與處理
將原代肝星狀細胞或肝細胞傳代培養至對數生長期后,分為空白對照組、LPS 模型組、秋水仙堿(1 μmol/L)組、三葉香茶菜提取物(1 mg/mL,質量濃度參考“2.3”項下IC50設置)組、TLR4 阻斷劑(TAK-242,1 μmol/L)組和TLR4 阻斷劑+三葉香茶菜提取物(1 μmol/L+1 mg/mL)組,每組設3 個復孔。空白對照組細胞加入無LPS的完全培養基,培養24 h;LPS模型組細胞加入含LPS(100 ng/mL)的完全培養基,培養24 h;各藥物組細胞先用含相應藥物或阻斷劑的完全培養基培養24 h 后,再用含LPS 的完全培養基培養24 h。LPS、TLR4、秋水仙堿的質量濃度參考相關文獻設置[11-12]。
2.5 指標檢測
2.5.1 纖維因子和炎癥因子含量 收集各組細胞上清液,以2 000~3 000 r/min離心20 min,收集上清液,采用ELISA 法以酶標儀于570 nm 波長處檢測肝星狀細胞上清液中α-SMA、TGF-β1、COL-Ⅰ、COL-Ⅲ的含量和肝細胞上清液中ALT、AST、IL-1β、IL-18的含量。
2.5.2 TLR4/NF-κB/NLRP3 通路相關基因表達 采用real-time PCR 法進行檢測。收集各組細胞并提取其總RNA。待測定濃度后,將總RNA 逆轉錄成cDNA,隨后進行PCR 擴增。PCR 反應體系包括:cDNA 模板1 μL、SYBR Premix Ex Taq(Ti RNaseH plus)7.5 μL、上/下游引物各0.6 μL、ddH2O 5.3 μL。PCR 反應條件為:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火10 s,72 ℃延伸30 s,共循環40 次。反應結束后,以β-actin 作為內參,采用2-ΔΔCt法計算目標基因mRNA 的表達量,并以空白對照組作為參照,計算各組目標基因mRNA 的相對表達量。實驗重復3次。所有引物均由生工生物工程(上海)股份有限公司設計、合成,引物序列及產物長度見表1。

表1 TLR4等目標基因的引物序列及產物長度
2.5.3 TLR4/NF-κB/NLRP3 通路相關蛋白表達 采用Western blot 法進行檢測。收集各組細胞,裂解后勻漿,以12 000 r/min離心5 min,取上清液。采用BCA法檢測蛋白濃度后,加buffer 適量并于100 ℃下變性10 min。取變性蛋白進行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳分離并轉移至聚偏二氟乙烯膜上,用5%脫脂奶粉封閉1 h,加入TLR4、p-IκBα、NF-κB p65、NLRP3、GSDMD、ASC、β-actin 一抗(稀釋度均為1∶1 000),于4 ℃下孵育過夜;用TBST溶液清洗15 min×3次,加入相應二抗(稀釋度為1∶4 000),于37 ℃下孵育90 min。以ECL 化學發光底物顯色后,置于凝膠成像系統下曝光成像并使用Image J 1.53 e軟件進行分析,以目標蛋白與內參蛋白的灰度值比值表示目標蛋白的表達水平。實驗重復3次。
2.6 統計學方法
采用SPSS 26.0軟件對數據進行統計分析。數據均以±s表示,多組間比較采用單因素方差分析,組間兩兩比較采用LDS-t檢驗。檢驗水準α=0.05。
3 結果
3.1 三葉香茶菜提取物對2種細胞增殖的影響
隨著三葉香茶菜提取物質量濃度的增加,肝星狀細胞和肝細胞的增殖抑制率均逐漸降低,提取物對上述2種細胞的IC50分別為1.66、1.88 mg/mL。因此,在不影響細胞增殖活力的前提下,本研究選擇1 mg/mL(此質量濃度下,該提取物對2 種細胞的增殖抑制率分別約為65%、71%)作為后續實驗中三葉香茶菜提取物的干預濃度。
3.2 三葉香茶菜提取物對2 種細胞上清液中纖維因子和炎癥因子含量的影響
與空白對照組比較,LPS模型組肝星狀細胞上清液中TGF-β1、α-SMA、COL-Ⅰ、COL-Ⅲ含量和肝細胞上清液中ALT、AST、IL-1β、IL-18 含量均顯著升高(P<0.05);與LPS 模型組比較,各藥物組上述指標含量均顯著降低(P<0.05)。與TLR4 阻斷劑組比較,TLR4 阻斷劑+三葉香茶菜提取物組細胞上清液中TGF-β1、α-SMA、COL-Ⅲ和ALT、AST、IL-1β、IL-18 含量均顯著降低(P<0.05);而三葉香茶菜提取物組上述指標與TLR4阻斷劑組比較,差異均無統計學意義(P>0.05)。結果見表2。
表2 三葉香茶菜提取物對2種細胞上清液中纖維因子和炎癥因子含量的影響(±s,n=3)
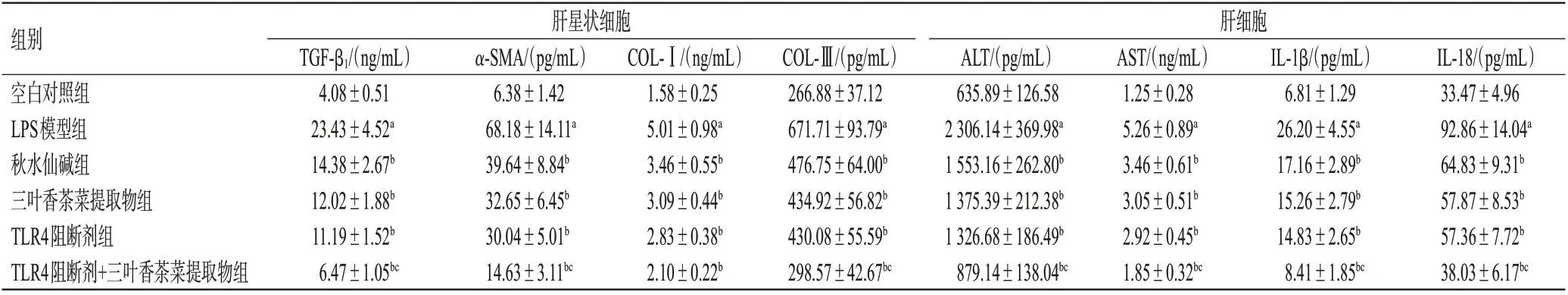
表2 三葉香茶菜提取物對2種細胞上清液中纖維因子和炎癥因子含量的影響(±s,n=3)
a:與空白對照組比較,P<0.05;b:與LPS模型組比較,P<0.05;c:與TLR4阻斷劑組比較,P<0.05
組別空白對照組LPS模型組秋水仙堿組三葉香茶菜提取物組TLR4阻斷劑組TLR4阻斷劑+三葉香茶菜提取物組肝星狀細胞肝細胞TGF-β1/(ng/mL)4.08±0.51 23.43±4.52a 14.38±2.67b 12.02±1.88b 11.19±1.52b 6.47±1.05bc α-SMA/(pg/mL)6.38±1.42 68.18±14.11a 39.64±8.84b 32.65±6.45b 30.04±5.01b 14.63±3.11bc COL-Ⅰ/(ng/mL)1.58±0.25 5.01±0.98a 3.46±0.55b 3.09±0.44b 2.83±0.38b 2.10±0.22b COL-Ⅲ/(pg/mL)266.88±37.12 671.71±93.79a 476.75±64.00b 434.92±56.82b 430.08±55.59b 298.57±42.67bc ALT/(pg/mL)635.89±126.58 2 306.14±369.98a 1 553.16±262.80b 1 375.39±212.38b 1 326.68±186.49b 879.14±138.04bc AST/(ng/mL)1.25±0.28 5.26±0.89a 3.46±0.61b 3.05±0.51b 2.92±0.45b 1.85±0.32bc IL-1β/(pg/mL)6.81±1.29 26.20±4.55a 17.16±2.89b 15.26±2.79b 14.83±2.65b 8.41±1.85bc IL-18/(pg/mL)33.47±4.96 92.86±14.04a 64.83±9.31b 57.87±8.53b 57.36±7.72b 38.03±6.17bc
3.3 三葉香茶菜提取物對2 種細胞中TLR4/NF-κB/NLRP3通路相關基因表達的影響
與空白對照組比較,LPS模型組肝星狀細胞和肝細胞中TLR4、NF-κB p65、NLRP3、GSDMD、ASC mRNA的表達均顯著上調(P<0.05),IκBα mRNA 的表達均顯著下調(P<0.05)。與LPS模型組比較,各藥物組2種細胞中TLR4、NF-κB p65、NLRP3、GSDMD、ASC mRNA的表達均顯著下調(P<0.05),IκBα mRNA 的表達均顯著上調(P<0.05)。與TLR4 阻斷劑組比較,TLR4 阻斷劑+三葉香茶菜提取物組2種細胞中TLR4、NF-κB p65、NLRP3、GSDMD、ASC mRNA 的表達均顯著下調(P<0.05),IκBα mRNA 的表達均顯著上調(P<0.05);而三葉香茶菜提取物組上述指標與TLR4 阻斷劑組比較,差異均無統計學意義(P>0.05)。結果見表3。
表3 三葉香茶菜提取物對2種細胞中TLR4/NF-κB/NLRP3通路相關基因表達的影響(±s,n=3)
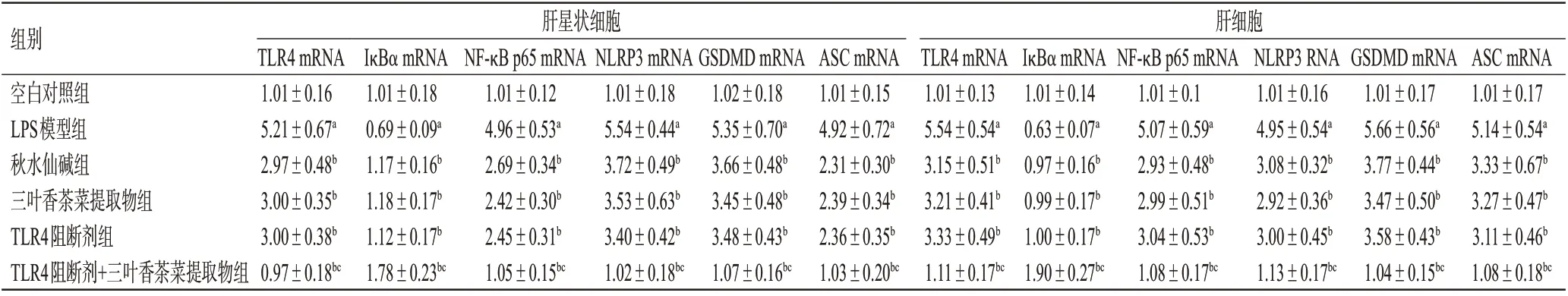
表3 三葉香茶菜提取物對2種細胞中TLR4/NF-κB/NLRP3通路相關基因表達的影響(±s,n=3)
a:與空白對照組比較,P<0.05;b:與LPS模型組比較,P<0.05;c:與TLR4阻斷劑組比較,P<0.05
組別空白對照組LPS模型組秋水仙堿組三葉香茶菜提取物組TLR4阻斷劑組TLR4阻斷劑+三葉香茶菜提取物組肝星狀細胞肝細胞TLR4 mRNA 1.01±0.16 5.21±0.67a 2.97±0.48b 3.00±0.35b 3.00±0.38b 0.97±0.18bc IκBα mRNA 1.01±0.18 0.69±0.09a 1.17±0.16b 1.18±0.17b 1.12±0.17b 1.78±0.23bc NF-κB p65 mRNA 1.01±0.12 4.96±0.53a 2.69±0.34b 2.42±0.30b 2.45±0.31b 1.05±0.15bc NLRP3 mRNA 1.01±0.18 5.54±0.44a 3.72±0.49b 3.53±0.63b 3.40±0.42b 1.02±0.18bc GSDMD mRNA 1.02±0.18 5.35±0.70a 3.66±0.48b 3.45±0.48b 3.48±0.43b 1.07±0.16bc ASC mRNA 1.01±0.15 4.92±0.72a 2.31±0.30b 2.39±0.34b 2.36±0.35b 1.03±0.20bc TLR4 mRNA 1.01±0.13 5.54±0.54a 3.15±0.51b 3.21±0.41b 3.33±0.49b 1.11±0.17bc IκBα mRNA 1.01±0.14 0.63±0.07a 0.97±0.16b 0.99±0.17b 1.00±0.17b 1.90±0.27bc NF-κB p65 mRNA 1.01±0.1 5.07±0.59a 2.93±0.48b 2.99±0.51b 3.04±0.53b 1.08±0.17bc NLRP3 RNA 1.01±0.16 4.95±0.54a 3.08±0.32b 2.92±0.36b 3.00±0.45b 1.13±0.17bc GSDMD mRNA 1.01±0.17 5.66±0.56a 3.77±0.44b 3.47±0.50b 3.58±0.43b 1.04±0.15bc ASC mRNA 1.01±0.17 5.14±0.54a 3.33±0.67b 3.27±0.47b 3.11±0.46b 1.08±0.18bc
3.4 三葉香茶菜提取物對2 種細胞中TLR4/NF-κB/NLRP3通路相關蛋白表達的影響
與空白對照組比較,LPS模型組肝星狀細胞和肝細胞中TLR4、p-IκBα、NF-κB p65、NLRP3、GSDMD、ASC蛋白的表達水平均顯著升高(P<0.05)。與LPS 模型組比較,各藥物組2種細胞中上述蛋白的表達水平均顯著降低(P<0.05)。與TLR4阻斷劑組比較,TLR4阻斷劑+三葉香茶菜提取物組2 種細胞中TLR4、p-IκBα(肝星狀細胞除外)、NF-κB p65、NLRP3、GSDMD、ASC 蛋白的表達水平均顯著降低(P<0.05);而三葉香茶菜提取物組上述指標與TLR4 阻斷劑組比較,差異均無統計學意義(P>0.05)。結果見表4、圖1。
表4 三葉香茶菜提取物對2種細胞中TLR4/NF-κB/NLRP3通路相關蛋白表達的影響(±s,n=3)
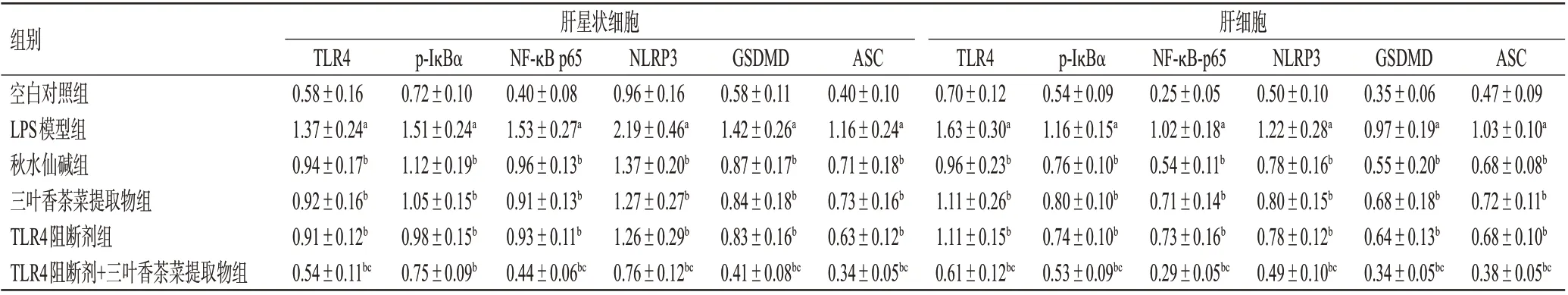
表4 三葉香茶菜提取物對2種細胞中TLR4/NF-κB/NLRP3通路相關蛋白表達的影響(±s,n=3)
a:與空白對照組比較,P<0.05;b:與LPS模型組比較,P<0.05;c:與TLR4阻斷劑組比較,P<0.05
組別空白對照組LPS模型組秋水仙堿組三葉香茶菜提取物組TLR4阻斷劑組TLR4阻斷劑+三葉香茶菜提取物組肝星狀細胞肝細胞TLR4 0.58±0.16 1.37±0.24a 0.94±0.17b 0.92±0.16b 0.91±0.12b 0.54±0.11bc p-IκBα 0.72±0.10 1.51±0.24a 1.12±0.19b 1.05±0.15b 0.98±0.15b 0.75±0.09b NF-κB p65 0.40±0.08 1.53±0.27a 0.96±0.13b 0.91±0.13b 0.93±0.11b 0.44±0.06bc NLRP3 0.96±0.16 2.19±0.46a 1.37±0.20b 1.27±0.27b 1.26±0.29b 0.76±0.12bc ASC 0.40±0.10 1.16±0.24a 0.71±0.18b 0.73±0.16b 0.63±0.12b 0.34±0.05bc GSDMD 0.58±0.11 1.42±0.26a 0.87±0.17b 0.84±0.18b 0.83±0.16b 0.41±0.08bc TLR4 0.70±0.12 1.63±0.30a 0.96±0.23b 1.11±0.26b 1.11±0.15b 0.61±0.12bc p-IκBα 0.54±0.09 1.16±0.15a 0.76±0.10b 0.80±0.10b 0.74±0.10b 0.53±0.09bc NF-κB-p65 0.25±0.05 1.02±0.18a 0.54±0.11b 0.71±0.14b 0.73±0.16b 0.29±0.05bc NLRP3 0.50±0.10 1.22±0.28a 0.78±0.16b 0.80±0.15b 0.78±0.12b 0.49±0.10bc GSDMD 0.35±0.06 0.97±0.19a 0.55±0.20b 0.68±0.18b 0.64±0.13b 0.34±0.05bc ASC 0.47±0.09 1.03±0.10a 0.68±0.08b 0.72±0.11b 0.68±0.10b 0.38±0.05bc
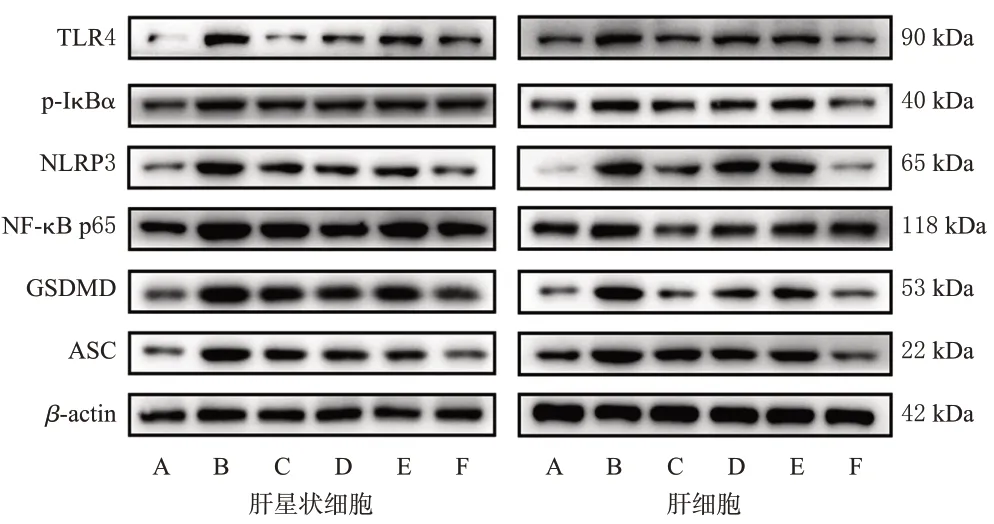
圖1 三葉香茶菜提取物對2 種細胞中TLR4/NF-κB/NLRP3通路相關蛋白表達影響的電泳圖
4 討論
本研究在前期動物實驗的基礎上,采用原代肝星狀細胞和原代肝細胞進行體外實驗,研究三葉香茶菜提取物對TLR4/NF-κB/NLRP3通路的影響,以進一步揭示其抗HF 的分子機制。研究表明,肝星狀細胞和肝細胞所構成的肝臟微環境直接影響HF 的進展,肝細胞通常是損傷的初始目標,隨后通過與肝星狀細胞間的相互作用(包括炎癥反應和再生反應)來直接或間接激活肝星狀細胞,促進細胞外基質(extracellular matrix,ECM)的合成、分泌[8,13]。因此,本研究采用原代肝星狀細胞和原代肝細胞開展體外實驗。
ECM 沉積是HF 的主要特征,也是該癥的主要驅動因素[14]。膠原蛋白是ECM 的主要結構性纖維不溶性蛋白,也是ECM 的主要成分[15]。TGF-β1在多種纖維化疾病(含HF)中起核心作用,能激活肝星狀細胞,促進ECM產生、沉積,從而導致HF 的形成[16-17]。在肝損傷期間,肝星狀細胞被激活并轉化為表達α-SMA的收縮性肌成纖維細胞,導致血管變形和血管阻力增加,從而引發門靜脈高壓,可見抑制α-SMA 可減緩HF 的進展[18]。本研究結果顯示,三葉香茶提取物能降低肝星狀細胞上清液中TGF-β1、α-SMA、COL-Ⅰ、COL-Ⅲ的含量,表明其可抑制TGF-β1活化,從而使肝星狀細胞轉化為收縮性肌成纖維細胞的途徑受阻,最終抑制了COL-Ⅰ、COL-Ⅲ等ECM成分的分泌。
在肝細胞與肝星狀細胞組成的微環境中,瀕死的肝細胞會觸發先天免疫系統,激活無菌性炎癥反應,從而對鄰近肝細胞造成進一步損傷并激活肝星狀細胞[19]。有證據表明,從死亡肝細胞中釋放的物質可作為肝星狀細胞激活和分化的重要介質,從而上調TGF-β1和COL-Ⅰ的分泌[20]。研究表明,LPS 可通過刺激肝細胞、激活NF-κB 通路來促進肝細胞中NLRP3、IL-1β、IL-18 的表達[21],上述炎癥因子可激活肝星狀細胞,促進膠原沉積,從而加重HF[8]。本研究結果顯示,三葉香茶菜提取物能降低肝細胞上清液中ALT、AST、IL-1β、IL-18的含量,表明其能減輕肝細胞的炎性損傷。
有研究指出,TLR4/NF-κB/NLRP3是一條重要的炎癥反應通路,TLR4 與LPS 結合可活化其下游IκB 激酶,使IκBα 磷酸化后降解,從而激活NF-κB,促進NLRP3、ASC 和胱天蛋白酶1 前體表達并組裝形成NLRP3 炎癥小體,活化的NLRP3 炎癥小體可促進細胞焦亡關鍵蛋白GSDMD活化、成熟,使細胞腫脹、裂孔而釋放IL-1β、IL-18 等炎癥因子,最終造成肝星狀細胞或肝細胞受損[22-23]。經LPS 刺激后,肝星狀細胞可通過TLR4 信號轉導和NF-κB 活化而被激活,分泌TGF-β1、α-SMA、COL-Ⅰ、COL-Ⅲ等因子,從而促進HF的發展;經LPS刺激后,肝細胞的TLR4 被激活,使IL-1β、IL-18 等炎癥因子分泌增加,從而造成炎性損傷[24-25]。此外有研究表明,NLRP3炎癥小體可誘導肝細胞死亡并釋放至細胞外被肝星狀細胞吞噬,導致IL-1β 和α-SMA 分泌的增加,從而加重HF[8]。本研究結果顯示,三葉香茶菜提取物能下調肝星狀細胞和肝細胞中TLR4、NF-κB p65、NLRP3、GSDMD、ASC mRNA 的表達,上調IκBα mRNA 的表達,并可降低TLR4、p-IκBα、NF-κB p65、NLRP3、GSDMD、ASC 蛋白的表達,降低肝星狀細胞上清液中TGF-β1、α-SMA、COL-Ⅰ、COL-Ⅲ的含量和肝細胞上清液中IL-1β、IL-18 的含量,提示三葉香茶菜提取物可通過抑制TLR4/NF-κB/NLRP3 通路活化來減輕肝細胞的炎性損傷,抑制肝星狀細胞的活化,減少纖維因子的合成分泌。
此外,本課題組前期動物實驗研究表明,作為陽性對照的秋水仙堿可明顯減輕大鼠HF 程度,本研究結果也得出了相似的結果。TLR4 阻斷劑可阻斷TLR4/NFκB/NLRP3 通路,抑制炎癥因子和纖維因子的表達[12]。本結果顯示,TLR4 阻斷劑同樣可下調肝星狀細胞和肝細胞中上述指標的分泌或表達,且TLR4 阻斷劑聯合三葉香茶菜提取物對上述多數指標的下調趨勢更加明顯,表明三葉香茶菜提取物抗HF作用除調控TLR4/NF-κB/NLRP3 通路外,可能還存在其他作用途徑,但尚需進一步驗證。
綜上所述,三葉香茶菜提取物能通過抑制TLR4/NF-κB/NLRP3通路活化,減少纖維因子、炎癥因子釋放,減輕肝細胞炎性損傷,抑制肝星狀細胞活化,從而發揮保護肝細胞、抗HF的作用。但是,本研究尚未明確三葉香茶菜對肝星狀細胞與肝細胞相互作用的影響,有待后續研究予以完善。
