鹽分脅迫下噴施γ-氨基丁酸對水稻秧苗生長的影響
2022-11-22 10:41:26馮棣高倩崔凱亓娜朱崴唐景春華澤田
中國稻米 2022年6期
馮棣 高倩 崔凱 亓娜 朱崴 唐景春 華澤田*
(1天津天隆科技股份有限公司,天津 300457;2南開大學 環境科學與工程學院,天津 300350;3濰坊科技學院,山東 壽光 262700;#共同第一作者:fengdi2008sunny@163.com,gao21318@163.com;*通訊作者:15904054936@139.com)
小站稻是津沽名特產品,名揚海內外。天津市廣泛分布著濱海鹽堿地,雖然大部分鹽堿地經過改良和連年種植土壤鹽度已經大幅降低,但水稻鹽害現象依然存在。以往研究表明,水稻在鹽堿脅迫環境下會產生抗逆機制,但當鹽堿脅迫超過植株所能調節的范圍時,會抑制水稻種子的萌發進程以及水稻的芽長、根長和根數[1-2];抑制葉片的伸長、新生葉的形成以及引起葉片卷縮和枯萎,葉尖變黃[3];還會抑制分蘗數,推遲分蘗進程,減少總穎花數,減輕千粒重,增加水稻籽粒的堊白粒率,最終導致籽粒產量和外觀品質下降,同時還會降低稻米膠稠度;嚴重的直接致死[4-5]。在水稻生長過程中,一旦發生鹽害,那么很難采取及時有效的措施加以改善。為了緩解作物所受到的鹽害,探索外源物種類及應用效果的研究成為一種潛在的解決途徑和研究熱點。高倩等[6]總結發現,目前已有50種外源物被報道在緩解作物鹽害方面可以發揮作用。γ-氨基丁酸(gamma-aminobutyric acid,GABA)屬于非蛋白質氨基酸,與植物體內碳素和氮素兩大代謝途徑緊密聯系[7],可以通過調節離子平衡、誘導合成滲透調節物質、誘導抗氧化酶活性、改善光化學系統、誘導基因表達及信號轉導共5條途徑提高植物耐鹽性[6]。王春燕等[8]研究發現,在80和150 mmol/L NaCl脅迫下,添加5 mmol/L GABA水培可以促進黃瓜幼苗根系對K+、Ca2+和Mg2+的吸收,抑制對Cl-、Fe2+、Fe3+和Na+的吸收,從而提高黃瓜苗耐鹽性。王馨等[9]研究發現,在不高于300 mmol/L NaCl脅迫下,葉面噴施5或10 mmol/L的GABA可以有效提高西伯利亞白刺葉片中Mg2+-ATPase活性、葉綠素含量(Chl)以及凈光合速率(Pn)。羅黃穎等[10]發現,在150 mmol/L NaCl脅迫下,添加5 mmol/L GABA水培明顯提高了番茄葉片生長速率、抗氧化酶活性、葉綠素含量和光合速率,減少了活性氧和膜脂過氧化產物MDA的積累,從而改善了番茄幼苗耐鹽性。趙九洲等[11]研究發現,GABA噴施濃度在25~50 mmol/L時,對鹽堿脅迫下甜瓜幼苗生長有一定緩解作用,但75 mmol/L時緩解作用極小。趙宏偉等[12]在土壤NaCl含量約為0.15%的條件下在水稻分蘗期和孕穗期連續2 d向葉面噴施4 mmol/L GABA,發現葉片中超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)等酶活性升高,MDA含量降低,水稻產量顯著增加,且分蘗期噴施對產量的提升效果優于孕穗期。綜上可見,外源GABA的有效性與使用方式、濃度、植物種類、發育階段、脅迫強度和時間有關。
在生產上,水稻一般采取插秧的方法種植,水稻種子萌發階段通過設施育苗可以得到很好的環境調控,但在秧苗3葉1心期插到大田之后才真正開始面對各種逆境的脅迫。因此,開展外源物(如GABA)改善秧苗耐鹽性的研究具有重要的實用價值,然而這方面的研究鮮見報道。鑒于此,本文以天津主栽粳稻品種天隆優619為研究對象,待秧苗培育到3葉1心時,開展外源噴施GABA對水稻地上部和根系生長,以及對抗氧化酶活性影響的研究,以期為改進鹽堿地水稻生產提供參考。
1 材料與方法
1.1 試驗材料
本試驗于2021年7—9月在國家粳稻工程技術研究中心開展,供試材料為粳稻品種天隆優619,由天津天隆科技股份有限公司提供。GABA由山東盛源生物科技有限公司提供。育苗基質由草炭土、珍珠巖和蛭石按比例摻拌而成(金盛,山東壽光),養分量:有機質35.3%,水解氮2 531 mg/kg,有效磷338.9 mg/kg,速效鉀2 334 mg/kg。水稻營養液配方參考張瑞坤等[13]研究配置。
1.2 試驗設計
1.2.1 育苗
選取粒大飽滿、大小一致的水稻種子,用95%酒精消毒,蒸餾水沖洗干凈后浸種36 h(種子未露白),浸種結束后點播。借鑒蔬菜育苗方式,采用營養土穴盤(72穴)育苗,以保證每個植株根系獨立。育苗在室外進行,培育過程中及時補水,不施肥。
1.2.2 水培試驗
待秧苗培育至3葉1心無分蘗時開始室內水培試驗。于水培杯(高10 cm、直徑8 cm)中進行,選用5個NaCl溶液濃度處理,分別為0、25、50、75、100 mmol/L。GABA試驗組的噴施濃度依據沙漢景等[14]研究結果選定為4 mmol/L,每天噴施1次外源物質,葉片正反面均噴至完全濕潤,不噴GABA對照組噴施水以保持變量均衡。使用S1~S5代表葉面噴水5個處理,GS1~GS5代表葉面噴施GABA 5個處理。試驗共10個處理,每個處理4次重復,每次重復6株秧苗。以水稻營養液為基礎溶液,水培液用基礎溶液配制不同濃度NaCl溶液,平均2 d更換或添加水培液1次。室內補光燈為全光譜每天固定開啟14 h。試驗于水培后8 d結束。
1.3 取樣與測定方法
1.3.1 幼苗形態指標
開始水培前及水培后第8天使用直尺調查所有水稻苗株高。水培后第8天調查所有水稻苗黃葉發生情況,統計死葉率;并在每個處理選取長勢一致的5株秧苗使用直尺測定展開葉的長和寬,并使用鄧啟云和吳愈山[15]提出的公式計算葉面積,如式(1)和式(2);再選取長勢一致的6株秧苗(含用于測量葉面積的5株),平分為3組,獲取根系(葉片用于測定抗氧化酶活性),使用精度為0.001 g的電子天平稱取鮮質量,之后用烘箱在75℃下烘干至恒質量,稱取干物質量,由干質量與鮮質量之比獲得干鮮比;從其余植株獲取鮮活根系,并采用試劑盒(蘇州科銘生物科技有限公司)測定根系活力。

式中,S為總葉面積;L為葉長;D為葉寬;K為水稻葉面積換算系數;x=L/D。
1.3.2 幼苗葉片抗氧化酶活性
取所有處理剩余植株的生長點下第3片展開的真葉剪碎混勻用于測定抗氧化酶活性。其中,SOD和POD酶活性采用試劑盒(蘇州科銘生物科技有限公司)測定,APX使用抗壞血酸法[16]測定,每個處理3次重復,各指標計算方法如式(3)至式(6)。
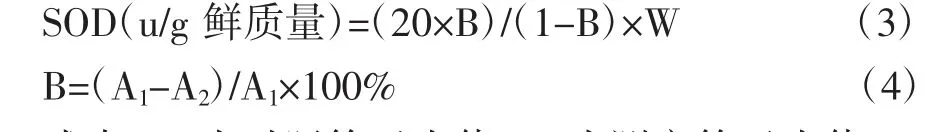
式中,A1為對照管吸光值;A2為測定管吸光值;W為樣本質量,g;B為抑制百分率,%。
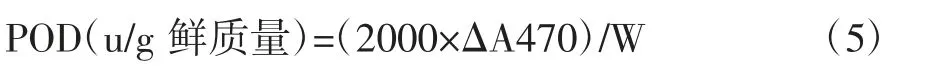
式中,ΔA470為1 min內470 nm處吸光值的變化;W為樣本質量,g。
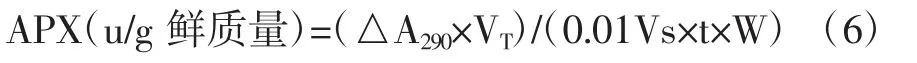
式中,△A290為1 min內290 nm處吸光值的變化;VT為提取液總體積,mL;Vs為酶液的體積;t為反應時間,s;W為葉片的鮮質量,g。
1.4 數據處理
用Excel 2019軟件處理數據和作圖,用SPSS 17.0數據處理系統進行差異顯著性分析和多因素方差分析。
2 結果與分析
2.1 GABA對鹽脅迫下水稻苗生長的影響
試驗8 d后不同處理水稻秧苗的生長狀態如圖1所示。由圖1可以看出,S4、S5、GS4和GS5處理秧苗生長狀態明顯差于S1、S2、S3、GS1、GS2和GS3處理,水稻葉片干尖現象更為嚴重;同時可以看出,噴施GABA處理較噴水處理存在改善效果。由圖2可以看出,隨著鹽分脅迫程度的增加,水稻的株高增量、葉面積呈下降趨勢,尤以株高增量在處理間差異最為明顯;死葉率呈上升趨勢,所有處理與相應0 mmol/L NaCl溶液水培處理相比均顯著(P<0.05)增加;株高因受到初始值的影響較大,在處理間未表現出一致性差異。在相同鹽分脅迫下,與不噴施GABA對照相比,噴施GABA后在0、25、50、75、100 mmol/L NaCl溶液水培處理下,株高增量分別增加-12.9%、1.7%、2.3%、32.4%和18.8%,葉面積分別增加-6.2%、4.9%、4.3%、9.1%和18.1%,死葉率分別降低63.6%、23.3%、4.8%、1.6%和5.1%。可見,噴施GABA可以緩解水稻受鹽脅迫的不良影響,改善秧苗地上部生長狀況,其作用效果與鹽分脅迫強度和表征指標等有關。
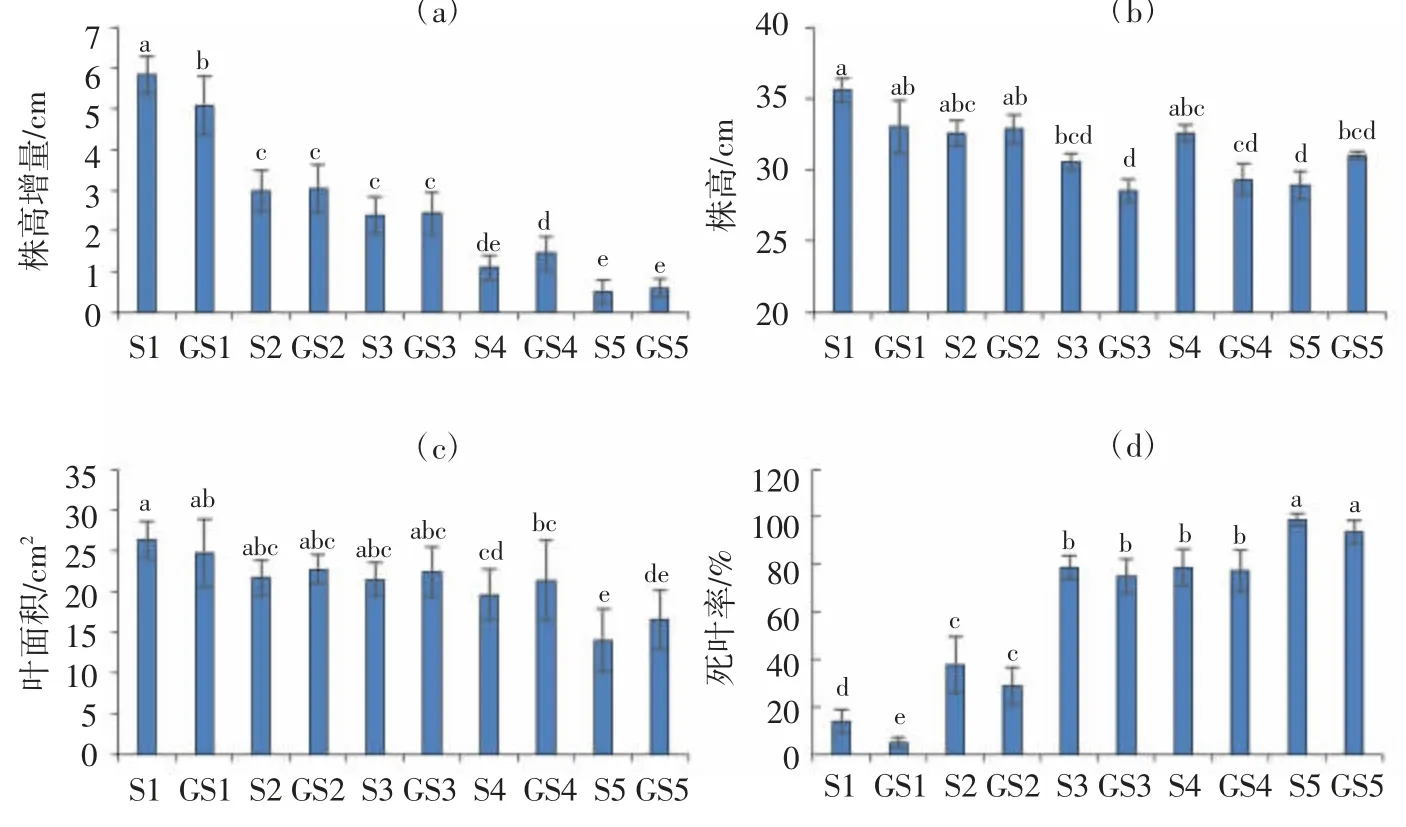
圖2 不同處理水稻幼苗生長情況調查
2.2 GABA對鹽脅迫下水稻苗根系的影響
由表1可以看出,隨著鹽分脅迫程度的增加,水稻根系干鮮比均呈上升趨勢,根系活力呈先升后降趨勢,根鮮質量和根干質量處理間存在小幅波動,但整體呈降低趨勢。在相同鹽分脅迫下,與不噴施GABA對照相比,噴施GABA后在0、25、50、75、100 mmol/L NaCl溶液水培處理下,根鮮質量分別減少18.1%、0.2%、12.5%、14.2%和11.7%,根干質量分別減少24.5%、9.3%、22.0%、19.6%和11.1%,根系活力分別增加76.1%、64.8%、48.6%、94.4%和1.2%。可見,噴施GABA降低了水稻根系干、鮮質量,提高了根系活力。
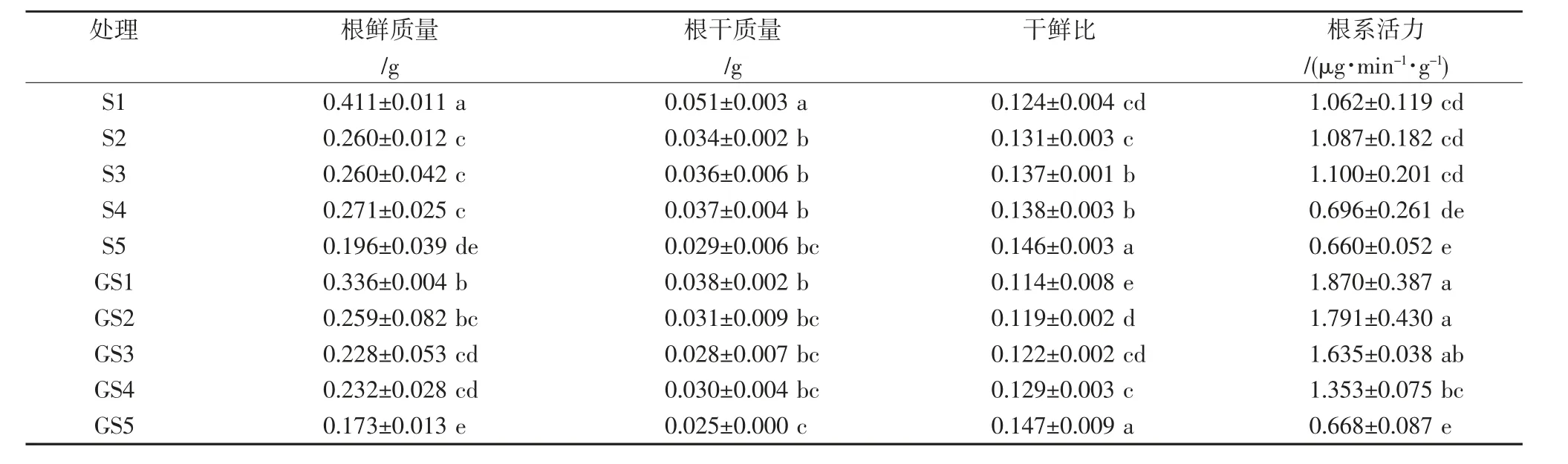
表1 不同處理水稻根系生長情況調查
2.3 GABA對鹽脅迫下水稻苗葉片抗氧化酶活性的影響
由圖3可以看出,隨著鹽分脅迫程度的增加,水稻葉片的POD和SOD酶活性均呈上升趨勢(圖3a、圖3b),APX酶活性呈先升后降趨勢(圖3c)。在相同鹽分脅迫下,與不噴施GABA處理相比,噴施GABA除0 mmol/L NaCl溶液水培處理SOD酶活性降低外,其余處理的3種酶活性均增加;在0、25、50、75 mmol/L NaCl溶液水培處理下,葉片POD酶活性分別增加25.0%、20.8%、5.0%和7.1%,SOD酶活性分別增加-6.5%、17.3%、15.8%和9.8%,APX酶活性分別增加4.6%、7.2%、54.8%和64.4%。可見,在鹽分脅迫下噴施GABA增加了水稻葉片的抗氧化酶活性。
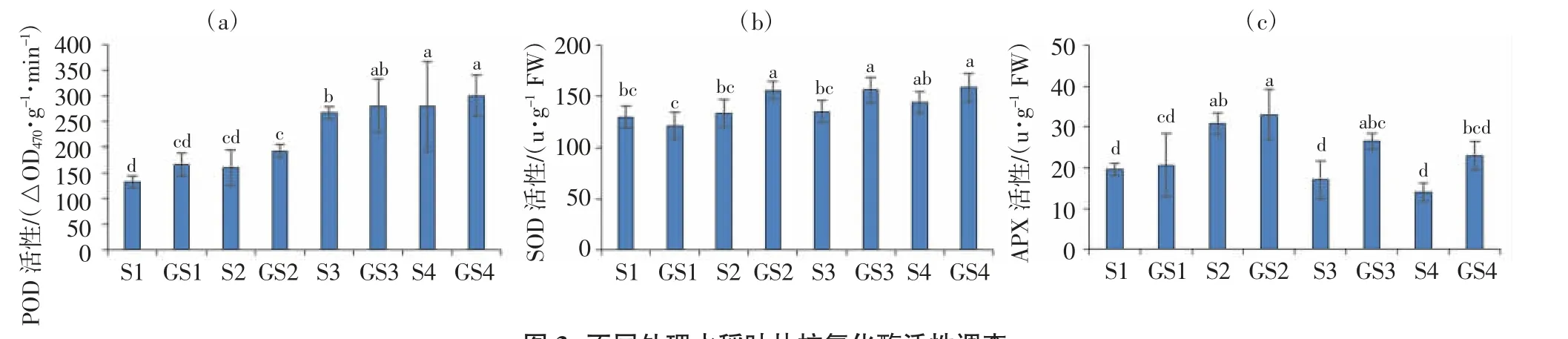
圖3 不同處理水稻葉片抗氧化酶活性調查
2.4 影響水稻秧苗生長的因素效應分析
由表2可以看出,是否噴施GABA對株高、株高增量和葉面積的影響不顯著,對死葉率、根鮮質量/干質量和根系活力的影響達到顯著水平(P<0.05);鹽分脅迫對除根鮮干質量以外所有生長指標的影響均達到顯著水平(P<0.05);二因素交互作用僅對根系活力產生顯著影響(P<0.05)。分析結果表明,鹽分脅迫對水稻苗生長指標的影響作用最大,其次為是否噴GABA,再者是兩因素互作。可見,噴施GABA可以影響水稻苗生長,且不同生長指標的響應程度存在差異,其中對根系的影響達到顯著水平。
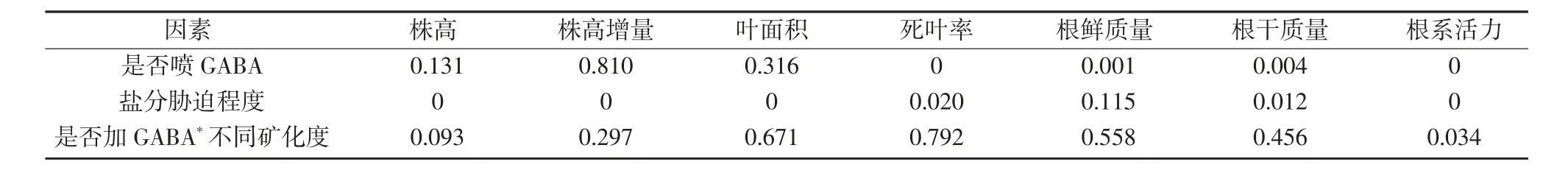
表2 水稻幼苗生長指標的單因變量二因素方差分析P值
3 討論
3.1 鹽分脅迫對水稻苗生長的抑制作用
本文模擬水稻插秧后即遭受鹽分脅迫。與以往研究結果一致,隨著鹽分脅迫程度的增加,水稻株高增量、葉面積、根干/鮮質量呈下降趨勢,這是因為鹽脅迫抑制了水稻光合作用[13],進而減少同化物的積累;水稻死葉率呈上升趨勢,這是因為Na+和Cl-的過量累積會灼傷葉片,導致葉片黃化[17];根系干鮮比呈上升趨勢,這是因為鹽分脅迫程度越高植株吸水越困難,植株體內自由水含量減少,表現為干鮮比上升。
與以往報道鹽分脅迫會降低水稻苗根系活力的結果不同,本試驗中根系活力呈先升后降趨勢,這可能是因為徐晨等[18]的研究是在80 mmol/L NaCl溶液脅迫下開展的,而本文設置的鹽分梯度更多,較低鹽分脅迫未對根系活力產生抑制作用。此外,水稻在遭受鹽分脅迫時會通過提高抗氧化酶活性來進行自我調整與適應,且抗氧化酶活性(SOD、POD、CAT)隨著鹽濃度增加先增后降[19]。在本試驗中,隨著鹽分脅迫程度的增加,水稻葉片POD和SOD酶活性呈上升趨勢,APX酶活性呈先升后降趨勢,與前人研究結果一致。通過分析影響水稻苗生長的因素效應,發現鹽分脅迫程度對水稻苗生長指標的影響作用最大,其次為是否噴施GABA,然后是兩者互作。
3.2 噴施GABA在提高水稻耐鹽性中起到的作用
3.2.1 噴施GABA對鹽脅迫下水稻秧苗抗氧化酶活性的影響
鹽脅迫下水稻內部細胞的離子平衡會遭到破壞,細胞膜遭受滲透脅迫,活性氧(ROS)累積過量,作物體內氧化還原平衡被打破,噴施GABA有利于幫助作物維持體內氧化還原平衡及細胞膜的完整性。因此也可以說抗氧化酶活性含量可以表征植物的耐鹽能力,且該能力的強弱受鹽分脅迫程度等因素的影響并呈單峰變化,即特定鹽分脅迫下抗氧化酶活性有且只有一個最優值域,在此最優值域內作物的耐鹽能力最佳。在生產中發現,水稻發生鹽害時,通過噴施外源物來緩解鹽害具有較強的可操作性。通過水稻苗生長的多因素效應分析發現,噴施GABA影響水稻苗的生長且不同生長指標的響應程度存在差異。本試驗結果表明,在相同鹽分脅迫下,與不噴施GABA對照相比,噴施GABA后3個鹽分脅迫(25、50、75 mmol/L NaCl)處理的水稻抗氧化酶(POD、SOD和APX)活性呈不同程度增加的趨勢,而且增加量均在作物生長接受范圍內并有促進作物正常生長的作用,這與前人關于玉米[20]、小麥[21]、水稻[12]、番茄[10]等作物的研究結果一致。
3.2.2 噴施GABA對鹽脅迫下水稻秧苗地上部分及地下部分的影響
在相同鹽分脅迫下,與不噴施GABA對照相比,噴施GABA后4個鹽分脅迫(25、50、75、100 mmol/L Na-Cl)處理的株高增量、葉面積不同程度增加,死葉率降低,根鮮干質量減少,根系活力增加。說明在鹽分脅迫下噴施GABA促進水稻苗地上部生長的同時提升根系活力。以往關于外源GABA改善植物地上部生長的報道較多[8-10,22],然而關于減少根系質量的報道鮮見。王春燕等[8]取4葉1心黃瓜苗在80和150 mmo/L NaCl脅迫處理6 d后,發現添加GABA可顯著提高根系鮮干質量。KAUR和ZHAWAR[22]采用培養皿育苗,第5天開始,在100 mmol/L鹽堿脅迫72 h后,添加1.5 mmol/L GABA處理的水稻幼苗地上部和根系干質量均有所增加。出現與以往研究不同的結果可能是因為:1)作為水田作物,水稻的耐淹水能力強,當外源GABA起到緩解鹽分脅迫的作用后,不需要再分配更多的同化物用于生長根系以吸收水分、養分和加強呼吸;2)與本研究的秧苗生長階段不同,KAUR和ZHAWAR研究的是芽苗階段,該階段胚根、胚軸迅速伸長,芽苗生長所需營養物質主要由種子提供,不涉及同化物的分配問題,外源GABA作為一種氨基酸態氮肥,促進了芽苗地上部和根的生長;3)由于本試驗采用的水稻營養液微量元素與大量元素并存,根據張宿[23]的報道可以推測,本試驗中根系質量的減少是由于水培液中NO3->40 mmol/L,這時GABA會抑制水稻苗根系伸長。
3.2.3 噴施GABA對鹽脅迫下水稻秧苗內同化物的影響
減少同化物向根系的分配也是本次試驗的發現之一。有研究表明,水稻劍葉維管束特征、穗頸和莖葉非結構性碳水化合物(non-structural carbohydrates,NSC)的轉運受氮含量影響,而外源物質GABA有調控作物體內碳氮流走向的作用,即GABA可以增強葉部氮同化能力[24]。因此,同化物向根系分配減少的原因可能是噴施GABA促使了莖葉同化物的轉運。
4 結論
外源物質GABA通過對水稻秧苗的生長調控,提高了鹽分脅迫下秧苗的抗氧化酶活性和根系活力,維持逆境中秧苗光合作用正常進行的同時促進了同化物優先向地上部分配轉運,進而提高作物水稻的耐鹽能力,為鹽分脅迫下水稻的穩產提供條件。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
小讀者(2021年2期)2021-03-29 05:03:48
文苑(2020年6期)2020-06-22 08:41:52
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
文苑(2019年22期)2019-12-07 05:29:00
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
