黃海綠潮生消過程及其主導因素*
2022-11-25 05:29:44張海彥
海洋與湖沼 2022年6期
王 圣 趙 亮① 張海彥 李 菲
(1. 海洋資源化學與食品技術教育部重點實驗室 天津 300457; 2. 天津科技大學海洋與環境學院 天津 300457; 3. 天津大學海洋科學與技術學院 天津 300072)
綠潮是一種在世界沿岸海域中普遍發生的大型海洋綠藻極速增殖的現象, 近年來成為一種世界范圍內的海洋生態災害, 主要發生在水體富營養嚴重的河口、瀉湖、人類活動密集的海岸等區域(王廣策等, 2020;張海波等, 2020)。據研究, 綠潮主要是由石莼屬(Ulva)、滸苔屬(Enteromorpha)、剛毛藻屬(Cladophora)、硬毛藻屬(Chaetomorpha)等大型藻類脫離其固著基后迅速增殖所造成的藻類災害。綠潮的暴發時間為每年的春夏之交, 暴發時間一般會持續2~3 個月, 并于秋季前完全消亡(劉湘慶, 2014)。我國黃海綠潮暴發主要是由滸苔引起的, 自2008 年至今, 黃海已連續13 a 暴發了不同規模的綠潮災害, 因此, 探究綠潮的生消過程對認識和預防綠潮有重要的作用。
從世界范圍來說, 綠潮在包括英國、法國、美國、日本、韓國、中國等10 多個國家的沿海地區均有發生(唐啟升等, 2010), Taylor 等(2001)在實驗室內研究了光照、溫度、氮營養鹽對綠潮生長的影響, Nelson等(2003)研究了美國華盛頓州沿海水域綠潮的空間及季節分布, Aurousseau (2001)和Cugier 等(2005)在法國綠潮災害嚴重的Brest 灣, 建立了三維生物地球化學模式, 模擬了該海域的漂浮綠潮的生消過程。Ruiz(1999)指出綠潮的生消過程受到了生物活動以及化學過程的影響, 同時綠潮的暴發會導致生態系統層面的變化。Merceron 等(2007)指出法國Brittany 灣綠潮藻類的生長受到光照強度、溫度和鹽度的影響。
我國學者們同樣做了大量關于綠潮生消過程的研究。吳洪喜等(2000)在實驗中發現滸苔適應環境能力較強, 是一種廣溫廣鹽性的海藻, 具有很強的環境適應能力。衣立等(2010)利用衛星遙感數據, 結合普林斯頓海洋模型(Princeton ocean model, POM)分析了綠潮暴發期間的水文條件以及滸苔定向漂移的原因。張海波等(2020)根據衛星監測數據及綠潮前后水文要素及氮營養鹽等數據, 對綠潮生消規律進行了分析,結果發現在不同階段綠潮受氮營養鹽影響不同。丁月旻(2014)探究了綠潮發生過程中滸苔成分的變化, 同時探究了滸苔生消過程中生源要素的轉化以及滸苔腐爛對環境的影響。白雨等(2019)分析了溫度、光照、降水在綠潮生消過程中的影響, 結論表明降水是影響綠潮生長規模的重要因素, 較高的溫度是導致綠潮消亡的主要原因。李雪娜等(2016)研究了海表面溫度與滸苔生消之間的關系, 認為滸苔暴發的適宜溫度范圍為23~26 °C。馮立娜等(2020)通過實驗探討了滸苔綠潮消亡過程中的營養鹽釋放規律以及滸苔綠潮聚積腐爛對海水水質的影響, 結果表明滸苔分解以有機態及顆粒態營養鹽為主。目前關于滸苔生消過程的研究已有很多, 但大多都是基于單個影響因子或實驗室條件下進行的研究。
本文基于考慮了溫度、光照、營養鹽的物理—生態耦合綠潮模型(LTRANS-GT), 對黃海滸苔的生消過程進行模擬, 探究了溫度、光照、營養鹽等影響因子在黃海綠潮生消過程不同階段中的作用。
1 生態模型建立與配置
1.1 綠潮生態模型及參數設置
黃海綠潮生態動力學模型主要分為兩個部分, 分別是漂移模塊以及生態模塊(圖1)。漂移模塊采用了拉格朗日粒子追蹤方法, 基于LTRANS 模型, 以海表面風場和表層流場數據作為驅動來計算綠潮的漂移軌跡。生態模塊中考慮了海表面溫度、光照強度、營養鹽等環境因子,建立了滸苔生長死亡的生態方程。進一步將生態模塊與LTRANS 模型耦合, 建立了物理—生態耦合的綠潮模型(LTRANS-GT), 來計算綠潮分布及生消過程。
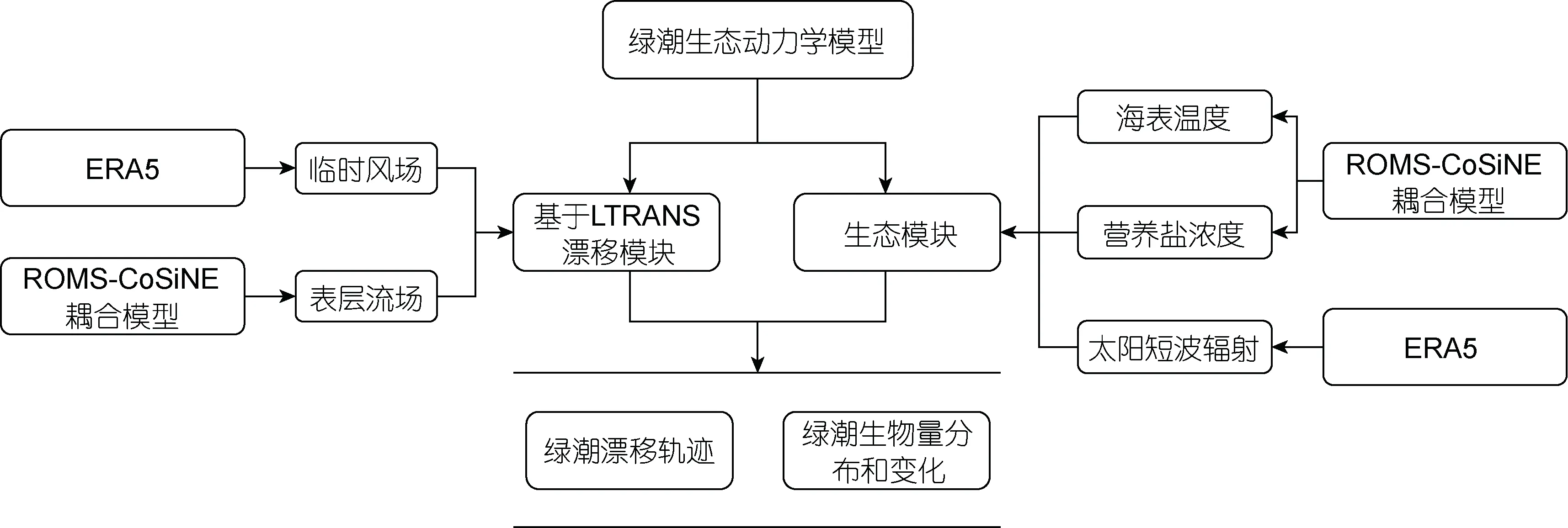
圖1 黃海綠潮生態動力學模型概念圖Fig.1 The model of ecological dynamics of green tide in Yellow Sea
漂移模塊考慮了海表面風以及海流下用的水平方向的物理運動過程。何恩業等(2021)指出, 滸苔漂移過程中會受到重力、浮力、波浪和湍流等作用, 這導致海流賦予滸苔的速度會有所下降, 在模型中我們將表層海流賦予滸苔的速度設定為表層海流速度的0.9 倍; 衣立等(2010)研究發現海表面風會賦予滸苔一部分速度, 同時在海表面風的作用下滸苔的運動方向會偏向盛行風右側約5°~40°, 在模型中我們將風速賦予滸苔的速度設定為風速的0.01 倍, 而由于科氏力造成的影響, 模型中設定滸苔的運動方向會偏向盛行風右側20°。生態模塊考慮了滸苔的生長及死亡, 其中模型中設定滸苔的生長受到了光照強度、溫度、營養鹽的控制, 而死亡僅受到了溫度的影響。在模型中我們將綠潮生物量的變化通過下面的公式來進行表達:
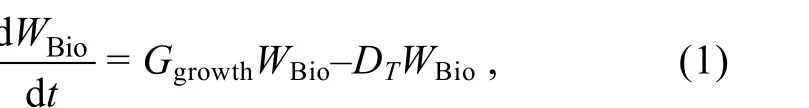
其中,WBio表示滸苔濕重, 單位是g,Ggrowth表示滸苔日生長率, 單位為d–1,DT表示滸苔受溫度影響的日死亡率, 單位為d–1。
溫度、光照和營養鹽對滸苔的生長起到了關鍵作用, 滸苔生長率的公式可以表達為
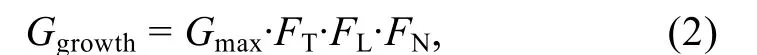
其中,Ggrowth為滸苔生長率,Gmax為滸苔最大生長率,FT、FL、FN分別為溫度、光照強度、營養鹽對滸苔生長速率的限制因子。
參考了Moisan 等(2002)建立的浮游植物動態生長-溫度模型, 改進了水溫對滸苔生長率的影響公式
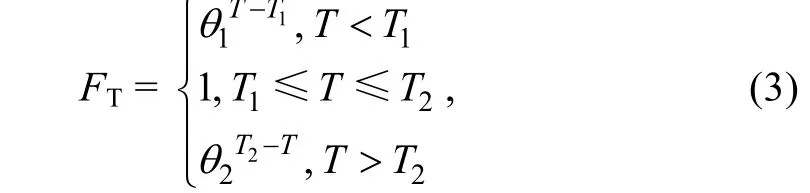
其中,T1、T2分別是最適溫度的上下限,θ1和θ2為生長率溫度調節系數。
海藻的光合作用速率在一定范圍內會隨著光照強度的增大而增加, 但超出了一定限度后反而會受到抑制(吳洪喜等, 2000), 在模型中, 光照對綠潮的影響參考Steele (1962)的公式:
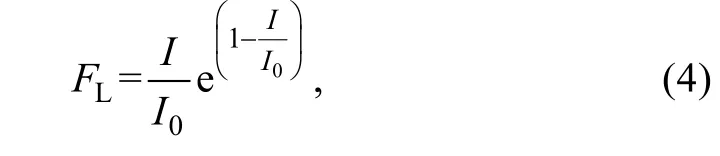
其中,I0表示最適合滸苔生長的光照強度。
滸苔的快速生長與較高的營養鹽濃度有著密切的關系, 本模型主要考慮了N營養鹽及P營養鹽對于滸苔生長的影響, 參考Monod 方程式我們給出滸苔生長受N 營養鹽及P 營養鹽的影響公式:

龐秋婷等(2013)指出, 在合適的磷酸鹽濃度下高濃度的硝酸鹽更能夠促進滸苔的生長, 在模型中, 我們設定當外界磷酸鹽濃度大于0.7μmol/L 時, 我們只考慮硝酸鹽對于滸苔生長的影響, 由此我們給出營養鹽對滸苔生長的影響公式:

其中,DT為滸苔受溫度影響的日死亡率;Dmax為日最大死亡率;θ3為死亡率系數;T3為最大死亡率臨界溫度。
上述模型中設計的變量如表1 所示。
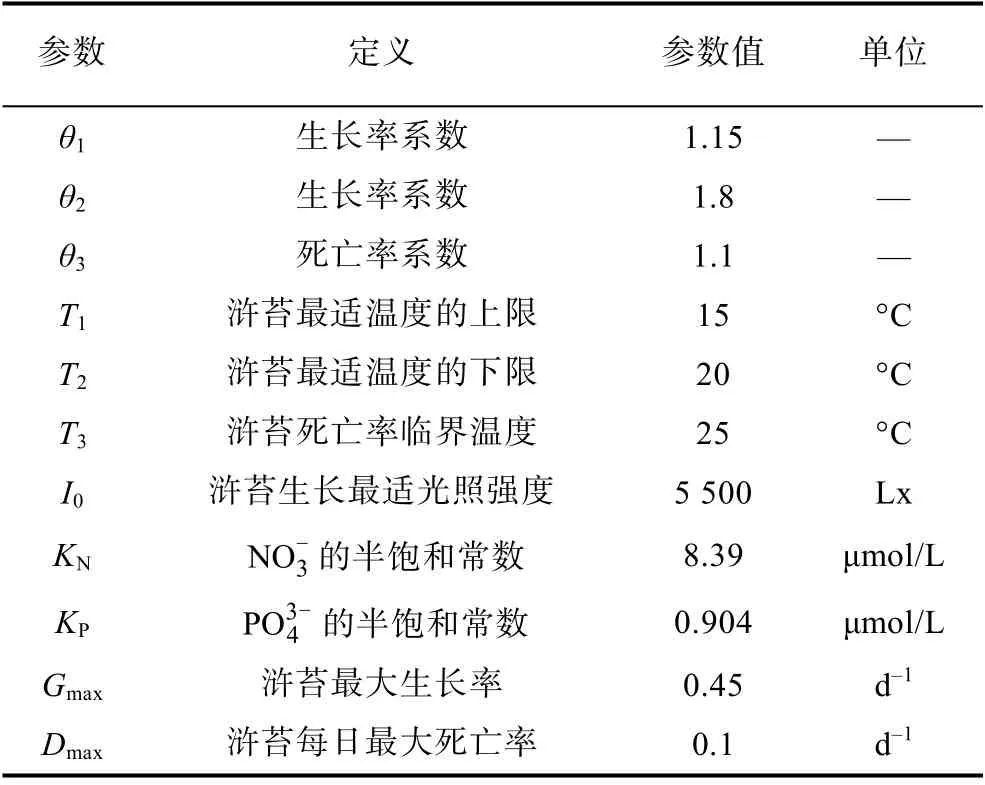
表1 模型參數Tab.1 Model parameters
1.2 模型配置及驗證
模型模擬的海域范圍是118°~128°E, 32°~38°N,模型模擬時間為2008~2017 年。模型每24 h 輸出一次結果, 結果包含滸苔粒子的經緯度、滸苔粒子的重量、滸苔粒子所在位置的環境(光照、溫度、營養鹽濃度)等。
用于模型輸入的表層流場、海表溫度和海表面營養鹽分布的數據, 來自ROMS-CoSiNE (regional ocean modelling system-carbon, silicate and nitrogen ecosystem)模型計算結果(錢思萌, 2018), 模型覆蓋了渤黃海及部分東海區域(117.01°~131.66°E, 29.04°~42.09°N)。數據垂向分層30 層, 空間分辨率為1/24°,時間分辨率為1 h, 數據時間范圍是從2008~2017 年。錢思萌(2018)、羅辰奕等(2019)對模型的物理場(環流、溫度、鹽度)和生態場(營養鹽、葉綠素、溶解氧)的時空分布特征進行了驗證。用于模型輸入的海面風場數據及光照強度數據來源于歐洲中期天氣預報中心(European Centre for Medium-Range Weather Forecasting, ECMWF)的ERA5 數據(Hersbachet al,2018), 其空間分辨率為1/4°, 時間分辨率為1 h, 所用到的數據時間范圍為2008~2017 年, 我們截取了渤黃海及部分東海區域(117°~131.75°E, 29°~42°N), 并插值到模型的計算網格上。
模型的初始場設置包括了粒子的初始位置、初始重量以及初始時間, 在模型中一次性釋放粒子4 000 個,本文利用郭偉(2017)、白雨等(2019)處理過的綠潮遙感信息, 對2008~2017 年衛星圖像中首次發現綠潮的時間與區域進行統計, 確定了輸入粒子的初始位置,劉湘慶(2014)在2012 年綠潮暴發過程中測得的數據表明, 滸苔發展初期滸苔密為0.45 g/m2, 結合綠潮首次發現時的分布面積, 反算得到每年綠潮初始生物量(濕重)的大小。將遙感首次發現綠潮的時間確定為模型的起算時間, 模型結束的日期為遙感影像最后一次觀測到有綠潮存在的日期。
得到模擬結果后, 我們將模擬結果與綠潮遙感信息進行了對比驗證, 如圖2 所示。整體上來看, 該模型對綠潮漂移路徑、轉向時間等方面的模擬結果與衛星遙感有較好的一致性, 基本可以刻畫綠潮的分布特征。
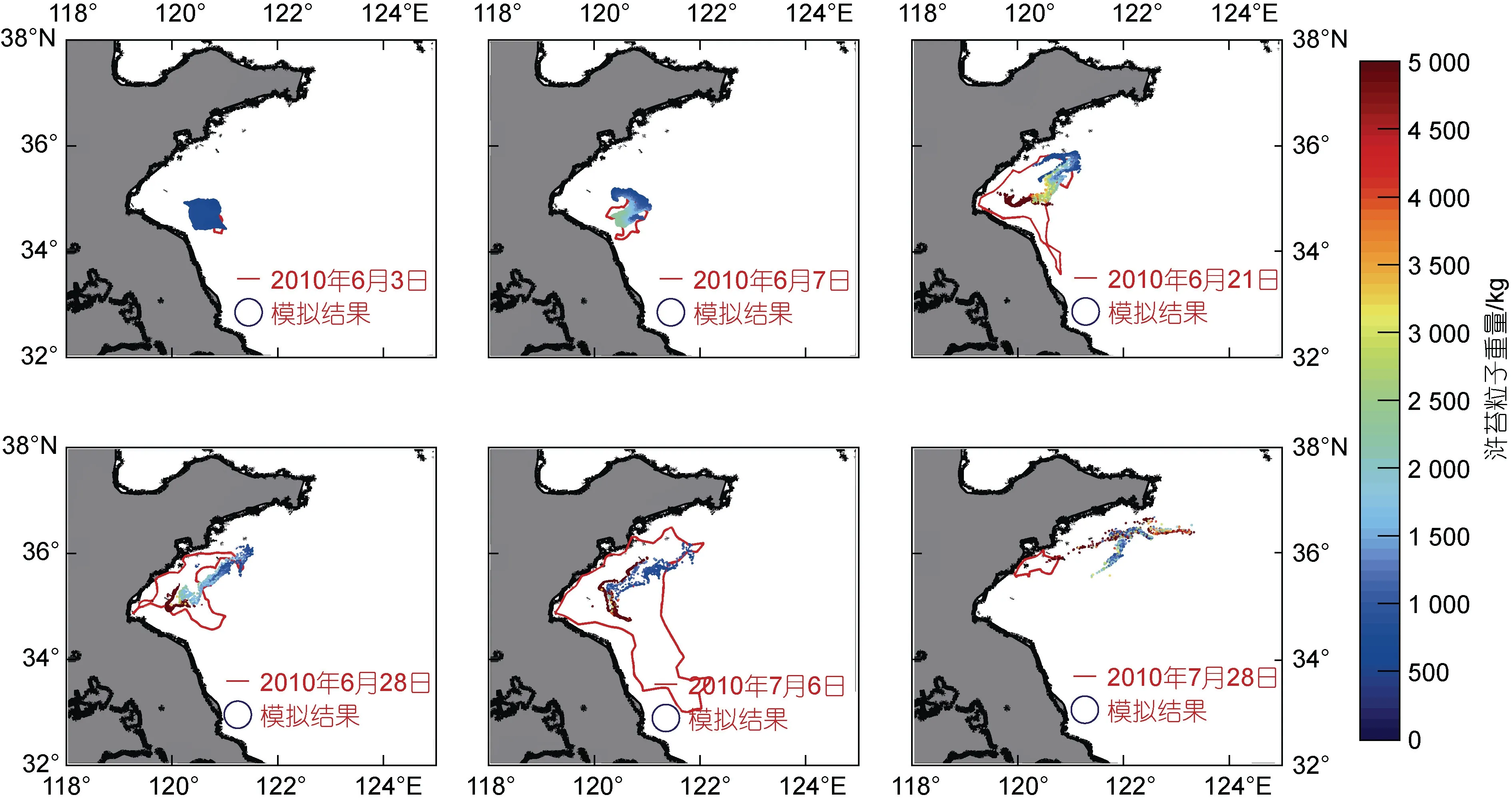
圖2 模型結果驗證圖Fig.2 Validation of the model results
2 結果與討論
根據模型模擬的2008~2017 年結果, 給出了黃海綠潮的生長消亡特征, 并分階段討論了溫度、光照、營養鹽在綠潮不同生長階段所起到的作用。
2.1 黃海綠潮生長消亡特征
模型模擬的整個綠潮生消過程持續62~94 d, 其中2010 及2017 年綠潮生消過程持續時間較短, 約60 d; 2008、2012、2014 年持續時間較長, 均在80 d以上; 其余年份均在70~80 d。過去的10 a 間, 綠潮的生物量也存在著明顯的年際差異, 如圖3 所示, 黃海綠潮生物量高值年份為2013、2015、2016 年, 這些年份最大生物量均超過50 萬t, 同時生物量極大值出現在2013 年, 當年最大生物量高達56 萬t。綠潮生物量低值年份為2009、2010、2017 年, 這些年份最大生物量均不足5 萬t, 同時生物量極小值出現在2017 年, 當年的最大生物量僅為2.5 萬t。其余年份最大生物量約10~36 萬t。
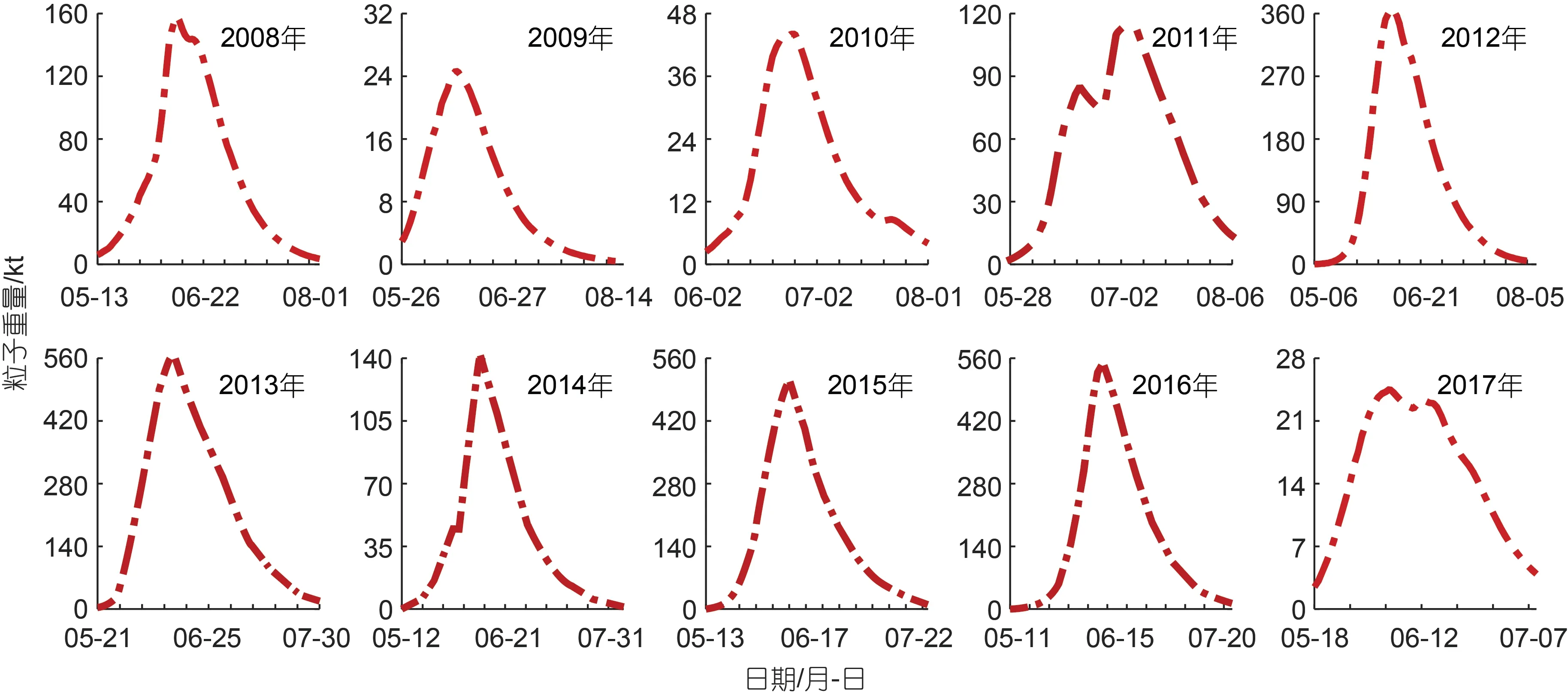
圖3 綠潮模擬生物量Fig.3 Simulated green tide biomass
雖然綠潮規模存在年際間差異, 但其季節規律較為類似, 除去2009~2011 這三年綠潮從6 月初左右開始暴發, 其余年份暴發時間均在5 月中旬前后, 隨后綠潮規模迅速擴大, 并于隨后的20~38 d 達到當年的生物量峰值, 統計顯示達到當年的生物量峰值平均需要29 d, 標準差為5 d, 說明每年綠潮在暴發后的約一個月生物量達到峰值。白雨等(2019)根據遙感數據分析也得出綠潮覆蓋面積約在30~40 d 達到當年峰值。之后綠潮于7 月份快速消亡, 至8 月份基本消失。
滸苔生物量不同年份間季節變化存在明顯差異,很多學者通過遙感影像得出綠潮分布面積呈現單峰值分布的結論, 但模擬顯示生物量變化形式并不單一。總體來說生物量季節變化可以分為以2011 年和2017 年為代表的雙峰值形式, 以及其他年份所表現出的單峰值形式, 以2011 年為例, 綠潮生物量于6 月18 日達到全年次高值, 之后生物量逐漸降低, 直到6月26 日達到近期最低值, 隨后又逐漸升高直到7 月4日達到全年生物量最高值。這一生物量分布特征也同樣出現在何恩業等(2021)的2019 年綠潮模擬結果中。
2.2 黃海綠潮生長階段主要控制因素
根據綠潮生物量的季節變化特征, 我們將綠潮開始暴發至到達最大生物量定義為綠潮的生長階段,將之后的時間定義為綠潮的消亡階段。在模型中滸苔的生長和消亡過程受溫度、光照、營養鹽的控制, 為了進一步了解綠潮在生長階段的受控因素, 我們對2008~2017 年連續10 a 的數據進行了統計分析, 結果如表2 所示。
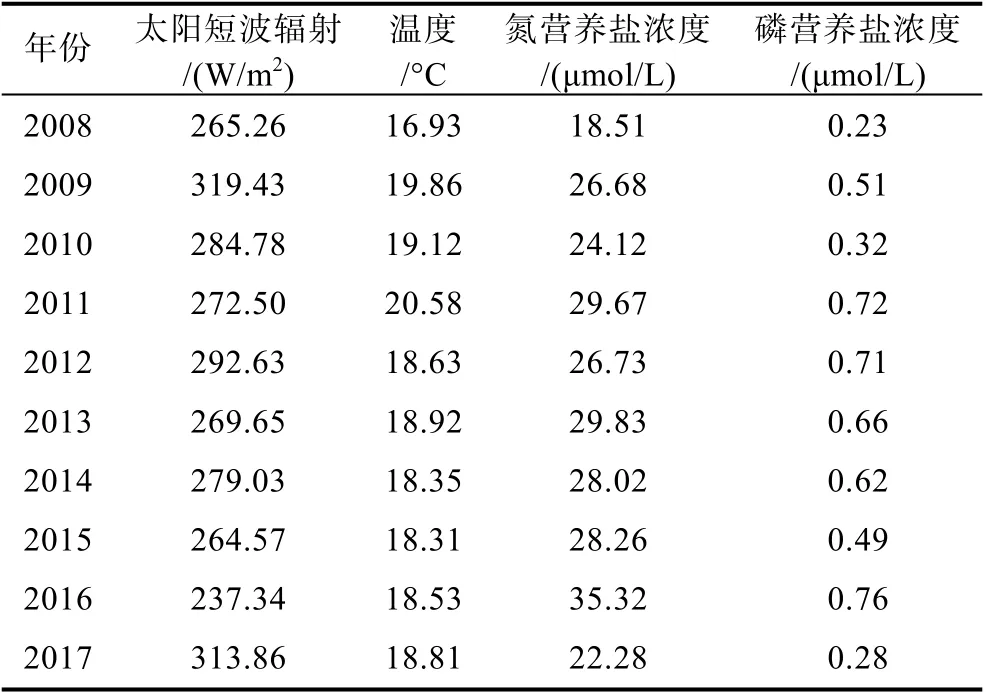
表2 生長階段綠潮暴發海域平均溫度、光照、營養鹽濃度Tab.2 Average temperature, light, and nutrient concentration of green tide in outbreak area during growth stage
從表2 中可以看出, 在綠潮覆蓋區域太陽短波輻射最低為237.34 W/m2, 最高為319.43 W/m2, 平均約為279.91 W/m2, 主要集中在260~290 W/m2的范圍內。吳洪喜等(2000)研究表明滸苔的最適光強為5 000~6 000 Lx, 馬汶菲等(2022)研究發現滸苔在光強144 μmol/(m2·s)條件下適合釋放孢子, 崔琳琳等(2014)研究表明, 在綠潮聚集前期短波輻射能量在200 W/m2左右, 光照條件有利于綠潮繁殖。而在模型中設定220 W/m2為滸苔生長的最適光照強度, 根據表中統計代入到生長公式中可以得知光照對滸苔生長的影響系數在0.89~0.98 范圍內, 表明在生長階段光照為滸苔生長提供了必要的條件, 但不是生長階段的限制性因素。
而綠潮覆蓋區在生長階段的海表面溫度普遍處于16~21 °C, 其中最低溫度為16.93 °C, 最高溫度為20.58 °C, 平均為18.81 °C。Taylor 等(2001)研究表明滸苔的最高生長速率出現在 15~20 °C, 吳洪喜等(2000)研究表明滸苔的最適溫度范圍為15~25 °C。在模型中設定15~20 °C 為滸苔生長的最適溫度, 同時在模型中此溫度下滸苔的日死亡率均小于8%, 因此從表中統計數據可以得出結果, 在生長階段溫度同樣為滸苔生長提供了必要的條件, 但并不是主要控制因素, 這與衣立等(2010)發現海表面溫度對滸苔暴發的影響不顯著的結論一致。同時模型在綠潮覆蓋區域的光照強度數據與溫度數據與白雨等(2019)的遙感結果非常接近, 這也間接證明了模型模擬效果較為理想。
而在生長階段綠潮覆蓋區的營養鹽濃度年際間波動較大, 氮營養鹽濃度處于18~36 μmol/L 范圍內,其中最低為18.51 μmol/L, 最高為35.32 μmol/L, 平均為26.94 μmol/L, 磷營養鹽濃度處于0.2~0.8 μmol/L范圍內, 其中最低為0.23 μmol/L, 最高為0.76 μmol/L,平均為0.53 μmol/L。代入模型中可以得知滸苔生長階段營養鹽對滸苔生長的影響系數在0.22~0.62 范圍內。張海波等(2020)研究表明, 在綠潮快速增殖階段,豐富的氮營養鹽是綠潮快速繁殖生長的物質基礎。龐秋婷等(2013)研究認為在磷酸鹽含量適中時, 高濃度的硝酸鹽更能促進滸苔的生長, 同時充足、連續的營養鹽補充是綠潮暴發的物質基礎。因此本文認為在合適的溫度、光照條件下, 營養鹽濃度可能是滸苔生長階段的主導因素。
為了驗證營養鹽濃度是其生長階段的主導因素這一認識, 本文設計了一組敏感性實驗, 在保持其他模型參數及輸入條件不變的情況下, 模型中不考慮營養鹽水平滸苔生長的影響, 即設定FN=1, 通過對比實驗前后滸苔的生消過程是否產生大的變化, 判斷營養鹽濃度是否是滸苔生長階段的主導因素。得到結果如圖4 所示。
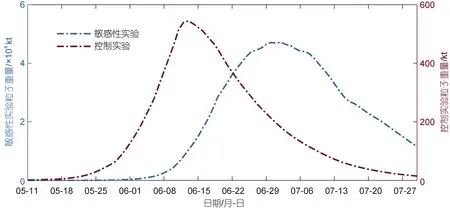
圖4 2016 年敏感性實驗生物量對比圖Fig.4 Comparison in sensitivity of experiment on biomass in 2016
以2016 年為例, 我們可以看到相較于正常狀態下的生物量曲線, 滸苔的生消時間在不考慮營養鹽影響的情況下得到了極大的延長, 同時最大生物量也有明顯的變化。2016 年在考慮營養鹽與不考慮營養鹽情況下, 生長階段所需時間分別為34 d 以及53 d, 敏感性試驗相較于控制實驗, 生長階段所需時間增加了19 d; 該年最大生物量分別為56 萬t 以及4 715 萬t, 敏感性試驗相較于控制實驗, 最大生物量增加了近100 倍。從結果可以看出, 營養鹽濃度影響生長階段的持續時間及最大生物量, 印證了在合適的溫度、光照條件下, 營養鹽濃度是滸苔生長階段的主要控制因素這一觀點。
滸苔生物量受其所在海域光照、溫度、營養鹽等環境因素的影響, 而造成滸苔所在海域環境因素差異的來源有兩個, 一個是滸苔不同年份分布區域不同所帶來的, 另一個是不同年份環境因素存在年際差異,以2009 年和2010 年為例討論這兩種來源的作用。
滸苔漂移受海表面風場及表層流場的影響, 流場及風場數據顯示, 2008~2017 年這10 a 間黃海表層流場及風場存在顯著的年際間差異。風、海流等動力因素不會直接影響滸苔的生物量, 但是會通過影響滸苔的漂移路徑及分布區域, 間接使不同年份間滸苔所在海域的溫度、光照、營養鹽產生差異, 從而造成滸苔生物量的差異。滸苔暴發一般集中于5 月中旬,同時在6 月經歷一個快速增長期, 圖5 給出了2009年與2010 年6 月表層平均流場及海表面平均風場的對比, 從圖5 中可以看出, 2009 年黃海海域6 月表層平均流速及海表面平均風速大小均高于2010 年, 同時流向及風向也存在顯著差異, 2009 年青島外海的流向與2010 年均呈現與岸大致平行的特征, 但2009 年的流向更加偏東。2009 年青島外海的風向為北向, 而2010 年的風向為西北向。表層流場與海表面風場的年際差異共同造成了滸苔分布區域的不同。與此同時,每年滸苔暴發的時間與初始位置存在差別, 也會使得滸苔分布存在差異。
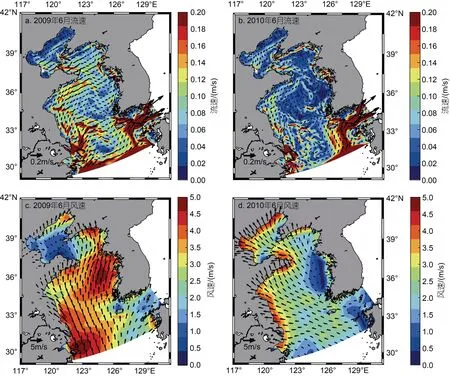
圖5 2009 年與2010 年表層流場及海表面風場對比圖Fig.5 Comparison of surface current field and sea surface wind field between 2009 and 2010
不同年份滸苔所在區域環境因素存在差異, 與滸苔分布區域的不同有著直接的關系, 但不排除環境因素空間分布存在年際間差異的影響, 圖6 給出了2009 與2010 年6 月海表面溫度數據及表層營養鹽濃度數據的對比, 同時標注了滸苔6 月所處的位置輪廓。從圖6 中可以看出, 黃海海域6 月溫度大小及結構存在差異, 但差異較小; N、P 營養鹽濃度大小及結構均無明顯差異, 營養鹽空間分布差異較為明顯,呈現出近岸高, 外海低的特點, 這與李伯志(2018)的統計與分析結果類似, 潘俊等(2018)也指出黃海中南部海域營養鹽分布并不均勻, 同時從圖6 中可以直觀看出滸苔所在區域營養鹽濃度存在顯著差異。因此,表層流場與海表面風場的年際差異共同造成了滸苔分布區域的不同, 使得滸苔所在區域環境因素存在差異, 從而對滸苔的生長造成影響。
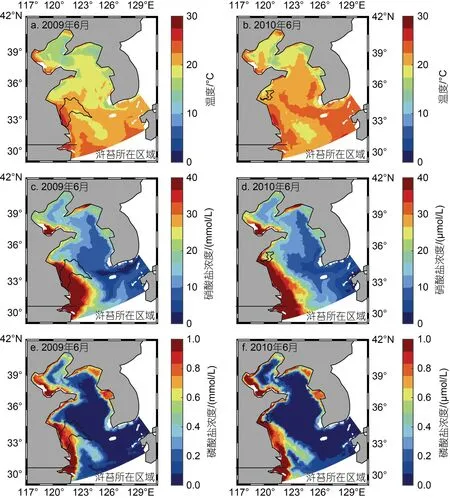
圖6 2009(a, b, c)年與2010(b, d, f)年溫度與營養鹽濃度對比圖Fig.6 Comparison in temperature and nutrient concentration between 2009(a, b, c) and 2010(b, d, f)
在大多數年份, 綠潮的季節變化較為相似, 生物量均呈現單峰值形式, 但在少部分年份, 由于短期極端天氣過程, 如臺風、強對流天氣等, 導致溫度、光照、營養鹽等環境因素急劇變化, 從而導致綠潮的季節變化特征與其他年份有所差異。以2011 年為例, 2011年第五號強熱帶風暴“米雷”于6 月22 日在菲律賓以東海面形成, 24 號加強為強熱帶風暴, 并沿中國東部沿海北移, 26 號下午到達山東半島以東海域, 并于26 號晚上于山東榮成登陸, 在臺風經過黃海的這段時間內, 綠潮覆蓋區域太陽短波輻射最低為72.84 W/m2, 最高為150.77 W/m2, 平均約為110.12 W/m2, 上文統計了正常天氣下太陽短波輻射約為主要集中在260~290 W/m2的范圍內, 相比之下臺風過境造成了太陽短波輻射的大幅度下降, 將臺風期間的光照強度數據代入到生長公式中可以得知光照對滸苔生長的影響系數在0.6~0.8 范圍內, 有明顯的下降。而在此期間綠潮覆蓋區域溫度下降約 0.5 °C, 氮營養鹽濃度增加約1.6 μmol/L, 磷營養鹽濃度增加約0.17 μmol/L, 代入到生長公式中可以算得溫度與營養鹽濃度的影響系數與臺風過境前無明顯差異, 因此我們推斷短期極端天氣現象而導致的光照強度驟降是生物量季節變化出現雙峰特征的主要原因。
2.3 黃海綠潮消亡階段主要控制因素
本文同樣對滸苔消亡階段區域的溫度、光照、營養鹽數據進行了統計分析, 結果如表3 所示。在滸苔消亡階段, 附近海域的太陽短波輻射最低為238.26 W/m2,最高為295.35 W/m2, 平均約為274.74 W/m2, 主要集中在260~290 W/m2的范圍內。相較于生長階段, 太陽短波輻射無明顯變化, 因此光照不是滸苔消亡階段的限制性因素。
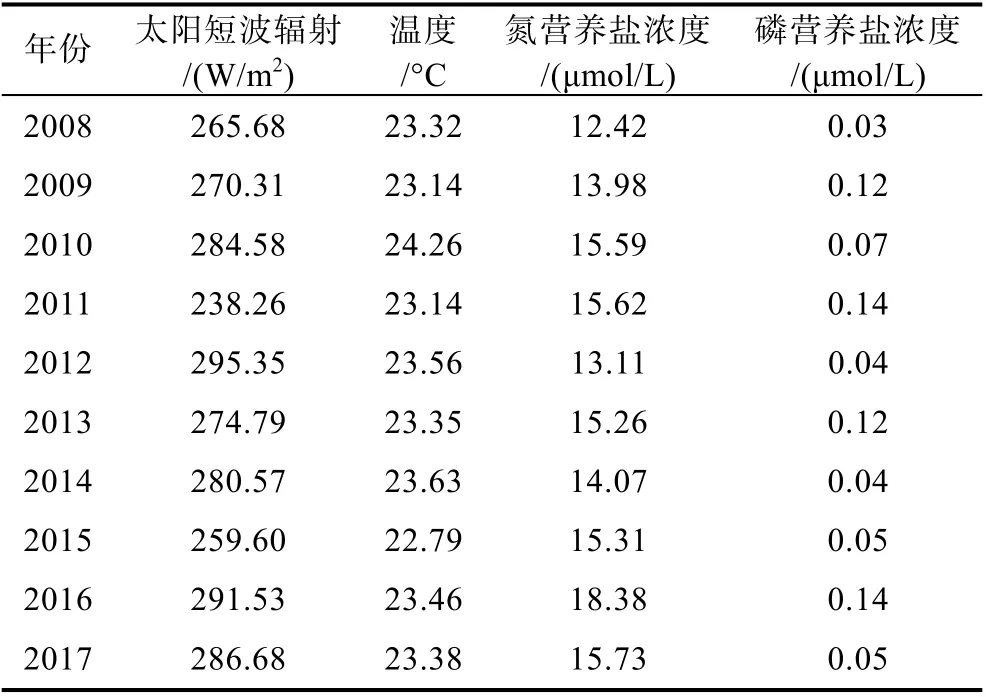
表3 消亡階段綠潮暴發海域平均溫度、光照、營養鹽濃度Tab.3 Average temperature, light, and nutrient concentration of green tide in outbreak sea area during the extinction stage
而在消亡階段綠潮覆蓋區的營養鹽濃度年際間波動不大, 氮磷營養鹽均處于較低水平, 氮營養鹽濃度處于12~19 μmol/L 范圍內, 其中最低為12.42 μmol/L,最高為18.38 μmol/L, 平均為14.95 μmol/L, 磷營養鹽濃度處于 0.03~0.1 μmol/L 范圍內, 其中最低為0.03 μmol/L, 最高為0.14 μmol/L, 平均為0.08 μmol/L。許多學者進行了有關滸苔對氮、磷營養鹽的實驗, 實驗結果均表明在其他條件不變的情況下, 較低的營養鹽濃度會限制滸苔的生長(邵魁雙等, 2011; 龐秋婷等,2013)。代入模型中可知滸苔消衰階段營養鹽對滸苔生長的影響系數在0.03~0.14 范圍內, 而從圖4 中的敏感性實驗結果我們可以看出, 在不考慮營養鹽的情況下, 滸苔消亡的趨勢并沒有停止, 說明在消亡階段營養鹽濃度不是唯一的主導因素。
消亡階段綠潮覆蓋區附近海域溫度處于22~25 °C之間, 最低為22.56 °C, 最高為24.26 °C, 平均約為23.41 °C, 相較于發展階段的平均溫度18.81 °C 有著很明顯的升高, 同時在消亡階段末期滸苔附近海域海表面平均溫度均超過了26 °C, 根據統計代入到生長公式中可以得知在消亡階段溫度對滸苔生長的影響系數在0.55~0.68 范圍內, 對滸苔死亡的影響系數在0.8~0.9 范圍內, 相比較而言, 滸苔生長階段溫度對滸苔生長的影響系數為1, 對滸苔死亡的影響系數普遍在0.5~0.6, 說明溫度的升高不僅降低了滸苔的生長率, 同時也增加了滸苔的死亡率。從圖7 中不同階段綠潮附近海域海表面溫度變化, 可以直觀得看出不同階段的溫度差異, 說明較高的溫度是滸苔的消亡的主導因素。辛蕾等(2018)根據遙感數據得出結論, 當表層水溫高于25 °C 時, 綠潮覆蓋面積開始下降。孟曉智等(2018)通過實驗室測定, 發現溫度高于25 °C 會明顯抑制滸苔生長。因此溫度同樣是滸苔消亡階段的主導因素。
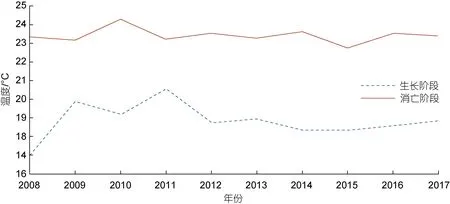
圖7 不同年份綠潮附近海域溫度對比圖Fig.7 Sea temperature in the areas near green tide in different years
綜上所述, 在滸苔消亡階段, 光照對于滸苔的消亡無顯著影響, 海表面風場及表層流場的差異造成滸苔分布區域不同, 帶來的海表面溫度及營養鹽濃度的差異共同影響了滸苔的消亡。這與其他學者的認識類似, 楊靜等(2017)表示因海水溫度上升、營養鹽含量降低以及其他環境因子的改變, 促使綠潮逐漸消亡。馮立娜等(2020)表明在高溫和乏營養鹽的條件下, 滸苔會很快進入腐爛分解階段。
3 結論
本文在LTRANS 輸運模型的基礎上, 增加了包含溫度、光照、營養鹽等環境因子的生態模塊, 構建了黃海滸苔生態動力學模型(LTRANS-GT), 并選用衛星遙感綠潮數據進行驗證, 最終模擬了2008~2017年黃海滸苔從暴發到消亡的全過程, 探究了海表面溫度、光照、營養鹽、海表面風及海流在滸苔生消過程的不同階段所起到的作用。
結果表明綠潮規模存在年際間差異, 但其季節規律較為類似, 每年于5 月中旬前后暴發, 隨后綠潮規模迅速擴大, 在暴發后的約一個月生物量達到峰值, 之后綠潮于7 月份快速消亡, 至8 月份基本消失。
生長階段滸苔周圍海域的溫度及光照處于適宜滸苔生長范圍內, 在模型中對滸苔的生長起不到顯著作用, 而營養鹽濃度年際間波動較大。海表面風場及表層流場的差異造成滸苔分布區域不同, 帶來的營養鹽濃度的差異是綠潮生長階段年際差異的主導因素。
消亡階段滸苔所在海域光照與生長階段相比無明顯差異, 因此光照對滸苔的消亡無顯著作用; 溫度相較于生長階段有著很明顯的升高, 同時在消亡階段末期滸苔附近海域海表面平均溫度均超過了26 °C,溫度的升高使得滸苔的生長率降低、死亡率升高; 此時氮磷營養鹽也處于較低水平, 限制了滸苔的生長。因此滸苔附近海域溫度的大幅升高及營養鹽濃度的下降共同影響了滸苔的消亡。
綠潮暴發受多種環境因素的控制, 而生態動力學模型在定量化描述黃海綠潮輸運及生消過程方面有著很強的優勢。雖然模型仍存在一些不足, 如使用的數據分辨率不夠高, 滸苔的生長和死亡過程的參數化方法還存在不確定性, 但模型的建立和應用, 可以為黃海綠潮的災害評估和應對措施的制定提供支撐。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
今日農業(2021年11期)2021-08-13 08:53:34
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
