溫度、冷層積和化學試劑處理對3種莎草科植物種子萌發的影響
2023-01-03 10:35:52張中華周華坤徐文華
草地學報 2022年12期
王 娜, 張中華, 周華坤, 徐文華*
(1.中國科學院西北高原生物研究所, 青海省寒區恢復生態學重點實驗室, 青海 西寧 810008; 2.中國科學院大學, 北京 100049)
莎草科植物(Cyperaceae)是陸地生態系統的重要構成者,具有水土保持、土壤改良、水源凈化等作用,也是青藏高原東部高寒草甸天然草地資源中飼用價值高、家畜喜食、耐牧性極強的一類優良牧草,是高寒草甸生態系統中分布最為廣泛、數量最多的科屬。莎草科植物適應性好,生境廣闊,具有很高的潛在價值,部分植物種是高寒草甸的建群種,其良好生長對高寒草甸植被恢復以及生態系統維持具有重要的現實意義及生態學價值。但莎草科植物種子普遍具有休眠、萌發困難的特性,嚴重制約了對其的開發利用。
種子的休眠與發芽特性因種而異,因此不同種子應選擇適宜其本身的方法來破除休眠,提高發芽率。目前對莎草科植物種子休眠與萌發特性的研究已取得了一定的進展,現有研究發現變溫和光暗交替有利于苔草屬種子的萌發[1],但是也有研究發現光照不影響莎草科植物種子萌發,甚至莎草科植物種子在自然光照梯度下幾乎不會萌發[2]。關于光照對莎草科植物種子萌發的影響還存在爭議,部分種子之間存在物種特異性[1]。激素和化學試劑處理已被證實是對莎草科植物種子萌發有利的技術[3],除此以外,低溫層積打破種子休眠,促進種子萌發的技術已經在多數實驗中得到驗證[4-5],而且低溫層積對嵩草屬和薹草屬種子的發芽率、初始發芽天數和發芽指數的影響因種皮的完整性、層積基質和草種不同而異[5]。
由于青藏高原高寒草甸地理環境特殊,探索該區種子休眠具有重要的生態學意義[6-8]。但是迄今為止,對于青藏高原莎草科植物種子萌發特性的研究方法較為單一,萌發技術研究的廣度、深度還不夠,對于高寒草甸這一特殊生境的研究更是鮮見報道,本研究通過不同物理和化學方法處理、化學處理+冷層積綜合處理,并且結合掃描電鏡觀測方法探索化學試劑處理對種子表皮的影響,試圖探討不同莎草科種子的適宜萌發溫度和光照條件,試驗處理對不同莎草科種子萌發的影響以及不同莎草科種子的休眠影響因素,從而尋求快速有效提高莎草科植物種子發芽率的方法,降低栽培草地建植成本,為青藏高原地區生態系統的恢復和畜牧業發展提供理論支持和技術指導。
1 材料與方法
1.1 材料與試劑
供試草種青藏薹草(Carexmoorcroftii)(以下簡稱QZ)、帕米爾薹草(Carexpamirensis)(以下簡稱PME)、華扁穗草(Blysmussinocompressus)(以下簡稱HB)的種子采集于海北高寒草地生態系統國家野外科學觀測研究站(青海省海北藏族自治州門源回族自治縣境內)。采集地生境:海拔多在3 400~3 450 m之間,地處37°29′~ 37°45′ N,101°12′~101°23′ E;年平均氣溫-1.7℃;最熱月7月平均氣溫10.1℃,最冷月1月平均氣溫-15.2℃。年降水量平均為582.1 mm,其中約80%降水量分布于植物生長季的5 — 9月;11月—次年2月降水量僅占年降水量的4%[9-10]。種子采集裝袋后帶回實驗室陰干(在此期間其果皮和種皮未受到影響),后置于牛皮紙信封中于室溫通風良好的條件下貯藏備用。供試的莎草科植物種子基本信息如表1所示。

表1 供試莎草科植物種子信息Table 1 Information of tested cyperaceae species seeds
試驗所用試劑:NaOH (AR,96%)、H2SO4(AR,98%)、NaHClO (60%)、戊二醛固定液(4%)、磷酸緩沖液(0.2 M,pH 3.5)、丙酮(AR,99.5%)、乙酸異戊酯(AR,99%,2.5 L)。
1.2 方法
1.2.1種子萌發實驗 萌發試驗開始于2022年1月底,采用培養皿內紙上萌發法,試驗開始前,所有種子用0.1% KMnO4溶液浸泡消毒10 min,再用蒸餾水沖洗3次,用濾紙吸干種子表面水分后在直徑為9 cm的圓形培養皿中放置2層用無菌蒸餾水潤濕的濾紙作為發芽床,將種子輕放于發芽床上,加蓋后放置在培養箱里,每個培養皿放置50粒顆粒飽滿的種子,設置3個重復。在整個試驗持續期間保持濾紙濕潤,每天統計發芽種子數。以露白為種子發芽的標準。計算各種子的發芽率、萌發時滯天數。
萌發時滯:指從發芽試驗開始到第1粒種子萌發所用時間。
發芽率=試驗結束時已萌發種子數/供試種子總數×100%。
1.2.2溫度與光照處理 根據種子采集地的氣溫設置了8個不同的溫度處理(包括5個恒溫和3個變溫)和2種光照條件,共計16類處理,具體設置見表2。
以上每種處理組合設置3個重復,每個重復50粒種子,每天統計萌發個數,試驗持續30天。
1.2.3冷層積處理 根據種子采集地的氣溫本研究確定了4℃的層積溫度。每種種子設置3個重復,每個重復50粒種子,分別置于培養皿中,加入無菌蒸餾水至剛好淹沒種子,用封口膜密封,于4℃冰箱中分別層積放置30天、60天、90天。每隔5天翻動檢查并沖洗換水1次,保持種子濕潤。以室溫條件下干燥貯藏未經層積處理的種子作為對照,以溫度試驗的結果選取萌發率適中的溫度條件作為層積處理后的培養條件,所以將層積處理完成的培養皿隨機置于設置為B25(15℃/25℃)溫度條件的培養箱內,光照條件設置為光照條件1(表2)下進行萌發實驗,以上處理均設置3個重復,每個重復50粒種子,每天統計萌發個數并補充水分,持續30 d。以露白為種子發芽的標準。
1.2.4化學試劑處理及化學試劑+冷層積處理 根據查詢到的關于莎草科植物種子萌發文獻資料中所用的萌發率較好的3種化學試劑和濃度處理方式對3種莎草科植物種子進行分別處理。
(1)用40%的氫氧化鈉(NaOH)溶液浸種30 min[11-13];
(2)用49%的硫酸(H2SO4)溶液浸種10 min[13-14];
(3)用10%的次氯酸鈉(NaHClO)溶液浸種1 h[15]。
試劑處理后的種子都用無菌蒸餾水反復沖洗后,放置在培養箱中培養。同樣的將B25溫度條件設置為試劑處理后的萌發溫度,置于光照條件1中(表2)進行萌發實驗,定期更換濾紙,及時處理發霉的種子,對照選取為同溫度下未經任何處理的種子。每個處理3個重復,每個重復50粒種子,每天統計萌發個數,持續30天。以露白為種子發芽的標準。
同時,對經化學試劑處理后的種子用無菌蒸餾水反復沖洗后,再置于4℃冰箱中冷層積處理1個月,期間每隔5 d翻動檢查并沖洗1次,保持種子濕潤,然后放置在B25、光照條件1(表2)的培養箱條件下進行萌發培養,每天統計萌發個數,持續30天。
1.2.5掃描電子顯微鏡觀察 選取上述供試種子及不同化學試劑處理后種子各20枚,置于解剖鏡下橫切或縱切,裝入青霉素小瓶,貼好標簽,然后將種子置于濃度4%的戊二醛中固定12 h,再用pH值為6.8的磷酸緩沖液沖洗5次,每次15 min,之后分別用30%,50%,70%,80%,90%,95%,100%丙酮梯度脫水,其中30%~95%濃度的丙酮只脫水一次,時間為20 min,而100 %濃度的丙酮進行脫水3次,每次30 min,脫水后用乙酸異戊酯置換2次,每次20 min,然后用二氧化碳臨界點干燥儀(IXRF VFD-21 s)干燥,最后粘臺,放置在掃描電鏡(SU8010)下觀察種皮、胚、胚乳及種子橫、縱切面結構并拍照。
用掃描電鏡觀察不同化學試劑處理對種皮的影響,結合發芽率試驗探究不同化學試劑處理對莎草科植物種子發芽的影響。
1.2.6數據處理 本文的所有數據在R語言(Version 4.1.1,R Foundation for Statistical Computing,Vienna,Austria)統計軟件中進行分析。首先對所有數據在分析前均進行Shapiro-Wilk正態分布和levene’s方差齊性檢驗,對非正態分布或方差不齊的數據進行反正弦平方根轉換;若轉換后不符合正態檢驗,則對數據進行kruskal非參數檢驗和Behrens-Fisher事后多重比較,對符合正態分布的數據進行單因素方差分析(One-way ANOVA)。然后Tukey’s HSD用于檢驗處理間多重比較的差異顯著性。統計數據以平均值標準誤差(Mean±SE)表示。采用Origin (Version 2018C SR,OriginLab Corporation,Northampton,MA,USA)軟件進行繪圖。
2 結果與分析
2.1 培養溫度和光照對種子萌發的影響
不同種子的萌發率受溫度和光照影響不同,其中華扁穗草(HB)種子萌發率受到溫度和光照的顯著影響(P<0.05),萌發時滯僅受到溫度的顯著影響(P<0.05)。帕米爾薹草(PME)種子的萌發率和萌發時滯僅受到溫度的顯著影響(P<0.05),而青藏薹草(QZ)在不同溫度和光照處理中沒有萌發(表3)。
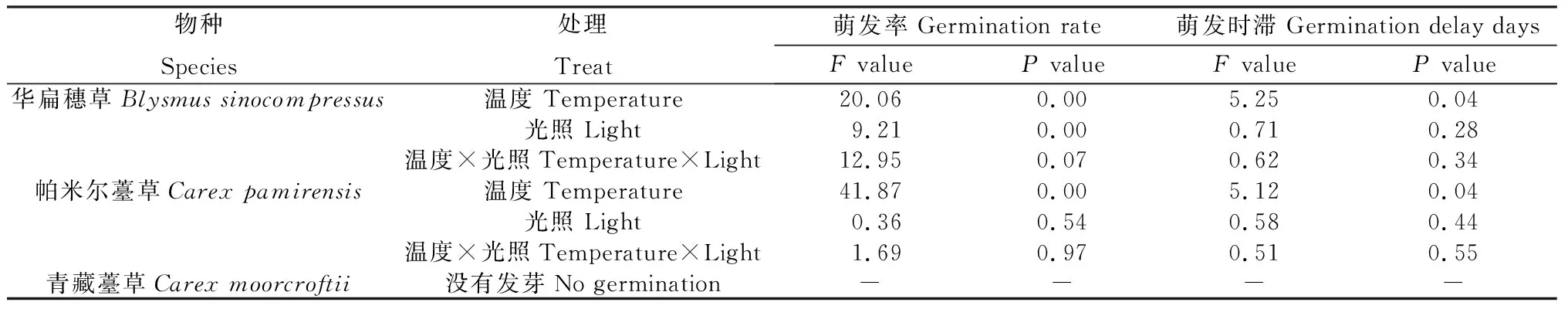
表3 不同溫度和光照對3種莎草科植物種子萌發率和萌發時滯的影響Table 3 Effects of different temperatures and light intensities on seed germination rate and germination delay of three cyperaceae plants
具體在每個溫度梯度上,有晝夜交替的光照條件下(表4),華扁穗草(HB)種子的最高萌發率為52.67%,出現在B25 (15℃/25℃)處理中,而隨著變溫溫度的升高,種子的萌發率逐漸降低,最低達到B35 (15℃/35℃)處理下的3.33%,顯著低于B25溫度處理的種子萌發率(P<0.05);帕米爾薹草(PME)種子萌發率隨著變溫溫度的升高,呈現先升高再降低的趨勢,萌發率在B30(15℃/30℃)處理中最高,為56.67%。而且這兩種種子在變溫處理中的萌發率均好于在恒溫處理中的萌發率。在24 h全黑暗的溫度處理中,HB種子僅在B35處理下有少量萌發,萌發率僅為1.33%,與晝夜交替光照條件下的萌發率差距明顯(P<0.05);PME種子的萌發率在全黑暗下與晝夜交替光照下沒有明顯差距,甚至略有上升,同樣表現為變溫條件下比恒溫條件下的萌發率高、隨著變溫梯度萌發率先上升后下降的變化趨勢。

表4 溫度和光照對種子萌發率的影響Table 4 Effects of temperature and light on seed germination rate
萌發時滯的變化與萌發率的變化情況類似(表5)。HB種子萌發時滯隨著變溫幅度增加顯著增加(P<0.05),恒溫條件下萌發時滯高于變溫條件。而PME種子無論在晝夜交替光照條件還是全黑暗條件下,隨著變溫溫度升高,均有萌發時滯天數先減少后增加的趨勢,盡管恒溫條件中出現了最短的萌發時滯,但萌發率并不高。

表5 溫度和光照對種子萌發時滯的影響Table 5 Effects of temperature and light on the time delay of seed germination 單位:d
通過數據分析,我們認為光照對HB種子的萌發是必需的,說明HB為喜光性種子;光照對PME種子的萌發幾乎沒有影響,說明PME種子為光不敏感性種子。對于HB和PME,變溫下的物種萌發率顯著高于恒溫下的萌發率(P<0.05),變溫下HB種子的萌發率最高達到52.67% (B25、光照),PME種子的萌發率最高達到56.67%(B30、光照),恒溫處理下普遍較低甚至為0,說明變溫處理更適合HB和PME種子的萌發。
2.2 冷層積處理對種子萌發的影響
本研究對3種莎草科植物種子施加了3種冷層積梯度(冷層積30天、60天、90天)處理,結果表明(表6、圖1),HB種子和PME種子萌發率和萌發時滯均顯著受冷層積時間的影響(P<0.05);青藏薹草(QZ)種子僅在層積時間90天有所萌發。

表6 冷層積時間對3種莎草科植物種子萌發率和萌發時滯的影響Table 6 Effects of stratification on seed germination rate and germination delay days of three cyperaceae species

圖1 層積對種子萌發的影響Fig.1 Effect of stratification on seed germination注:不同小寫字母表示同一物種層積不同時間萌發率之間存在顯著差異(P<0.05)。CJ30代表冷層積30天;CJ60代表冷層積60天;CJ90代表冷層積90天Note:Different lowercase letters indicate significant differences in germination rates at different stratification times for the same species (P<0.05). CJ30 represents cold stratification 30 days;CJ60 represents cold stratification 60 days;CJ90 represents cold stratification 90 days
所有種子在冷層積處理之后置于15℃/25℃的溫度培養條件下培養,如圖1所示,與對照組相比,華扁穗草(HB)種子經冷層積處理30天后萌發率顯著提高到81.33%(P<0.05);冷層積處理60天、90天的萌發率相同,而且顯著高于對照組的萌發率(P<0.05),但是并沒有與冷層積處理30天的萌發率有顯著差異。與對照組相比,帕米爾薹草(PME)種子經冷層積處理30天后萌發率反而顯著降低了(P<0.05),而冷層積處理60天、90天的萌發率比對照組萌發率有所提高但差異并未達到顯著性。另外,隨著冷層積時間延長,PME種子萌發率顯著提高(P<0.05),冷層積處理30天、60天、90天后的萌發率分別達到17.33%,48.00%和58.67%。
HB種子經冷層積處理60天時萌發時滯最小,為5天,與冷層積處理90天的萌發時滯沒有顯著性差異,但是與對照組及冷層積處理30天的萌發時滯均存在顯著性差異(P<0.05)。PME種子經冷層積處理90天的萌發時滯最小,為7天,與對照組的萌發時滯有顯著性差異(P<0.05)。青藏薹草(QZ)只有在冷層積90天處理下有少量種子萌發,萌發率為1.33%,萌發時滯為9天。
通過數據分析表明:與對照組相比,冷層積處理能顯著提高HB種子的萌發率(P<0.05),使萌發率最高達到90%(冷層積60天、90天處理),并降低其萌發時滯,使發芽時間提前到了5天(冷層積60天處理);冷層積處理使PME種子的萌發率最高達到58.67%(冷層積90天處理),同時使發芽時間提前到了7天。
2.3 化學試劑處理及化學試劑+冷層積處理對種子萌發的影響
本研究對3種莎草科植物種子進行了三種化學試劑處理及化學試劑處理后冷層積30天的處理。通過分析發現(表7、表8),化學試劑僅對HB種子萌發率有顯著影響(P<0.05),對PME和QZ種子萌發率的影響未達到顯著水平,對三種種子的萌發時滯均沒有顯著影響。化學試劑+冷層積處理對HB種子的萌發率和萌發時滯影響顯著(P<0.05),僅對PME種子的萌發率有顯著影響(P<0.05),對QZ種子萌發率和萌發時滯沒有顯著影響。
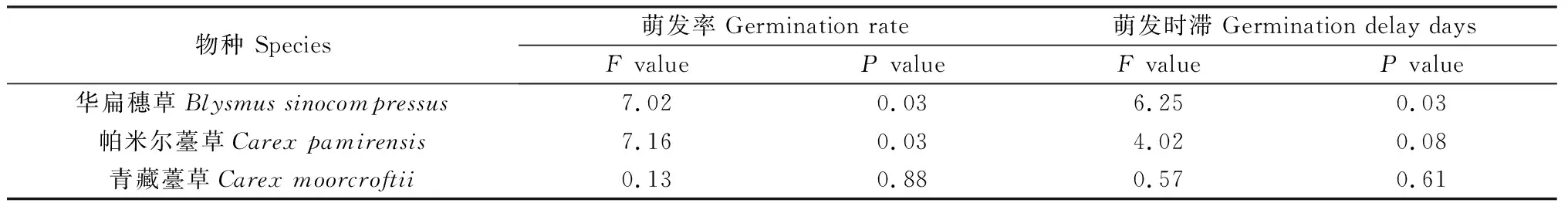
表8 化學試劑+冷層積處理對3種莎草科植物種子萌發率和萌發時滯的影響Table 8 Effects of chemicals + cold stratification on seed germination rate and germination delay days of three cyperaceae species
如圖2A所示,華扁穗草(HB)種子在H2SO4+冷層積綜合處理后的萌發率由僅用H2SO4處理后 21.33%提高到了92.67%(P<0.05),與對照組52.67%的萌發率相比也有顯著性差異(P<0.05),但是與僅通過冷層積處理30天后81.33%的萌發率沒有顯著差異;NaOH +冷層積綜合處理后的萌發率達到96.00%,比僅用NaOH處理后12.00%的萌發率提高了近7.00倍(P<0.05),與對照組52.67%的萌發率相比也有顯著性差異(P<0.05),但是與僅冷層積處理30天后81.33%的萌發率沒有顯著差異;NaHClO +冷層積綜合處理后的萌發率為83.33%,比僅用NaHClO處理后20.00%的萌發率提高了3.16倍(P<0.05),與對照組的萌發率相比顯著提高(P<0.05),但與僅冷層積處理30天后的萌發率沒有顯著性差異;如圖2B所示,H2SO4+冷層積綜合處理和NaOH +冷層積綜合處理后的萌發時滯最低,均為3天,與對照組的萌發時滯(10天)和僅冷層積30天處理后的萌發時滯(9天)相比均有顯著性差異。
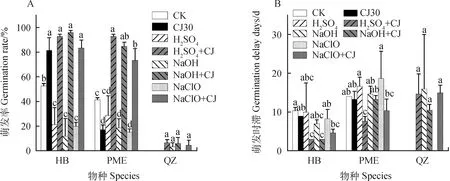
圖2 化學試劑處理及化學試劑+冷層積處理對種子萌發的影響Fig.2 Effects of different chemicals and chemicals + stratification on seed germination注:不同小寫字母表示同一物種不同處理之間萌發率或萌發時滯存在顯著差異(P<0.05)。CK代表對照組;CJ30代表冷層積30天;H2SO4 + CJ代表H2SO4處理后又冷層積處理30天;NaOH + CJ代表NaOH處理后又冷層積處理30天;NaHClO + CJ代表NaHClO處理后又冷層積處理30天Note:Different lowercase letters indicate significant differences in germination rates or germination delay days between treatments for the same species (P<0.05). CJ30 represents cold stratification for 30 days;H2SO4 + CJ represents cold stratification for 30 days after H2SO4 treatment. NaOH + CJ represents cold stratification for 30 days after NaOH treatment;NaHClO + CJ represents cold stratification for 30 days after NaHClO treatment
帕米爾薹草(PME)種子在H2SO4+冷層積綜合處理后的萌發率(92.67%)比僅用H2SO4處理后的萌發率(28.67%)提高了2.23倍(P<0.05),比對照組的萌發率(41.33%)提高了1.24倍(P<0.05),而且比僅冷層積處理30天后的萌發率(17.33%)提高了4.34倍(P<0.05);NaOH +冷層積綜合處理后的萌發率(84.67%)比僅用NaOH處理后的萌發率(16.11%)提高了5.00倍(P<0.05),比對照組的萌發率(41.33%)提高了1.04倍(P<0.05),比僅冷層積處理30天后的萌發率(17.33%)提高了3.89倍(P<0.05);NaHClO +冷層積綜合處理后的萌發率(73.33%)比僅用NaHClO處理后的萌發率(15.33%)提高了3.78倍(P<0.05),比對照組的萌發率(41.33%)提高了0.72倍(P<0.05),比僅冷層積處理30天后的萌發率(17.33%)提高了3.23倍(P<0.05);在萌發時滯中,H2SO4+冷層積綜合處理后的萌發時滯最低,為7天,與對照組的萌發時滯(14天)和僅冷層積30天處理后的萌發時滯(13天)相比均有顯著性差異。
青藏薹草(QZ)種子經過NaOH處理、NaOH +冷層積處理、H2SO4+冷層積處理和NaHClO +冷層積處理后均有種子萌發,但是平均萌發率最高只達到10%,萌發時滯最短為10天。
通過數據分析表明:不管是HB種子還是PME種子,NaOH +冷層積、H2SO4+冷層積和NaHClO +冷層積綜合處理后萌發率均顯著高于對照組的萌發率。對于PME種子,3種化學試劑+冷層積綜合處理后的萌發率不僅顯著高于對照組的萌發率,同時也顯著高于僅冷層積處理30天后的萌發率。HB種子萌發率最高達到96.00%(NaOH +冷層積處理),萌發時滯最低達到3天(NaOH +冷層積處理,H2SO4+冷層積處理);PME種子萌發率最高達到92.67%(H2SO4+冷層積處理),萌發時滯最低達到7天(H2SO4+冷層積處理)。
2.4 化學試劑處理對種子表皮的作用
研究中選取的化學試劑主要對種皮結構有影響,以青藏薹草為例,電鏡觀測發現不同化學試劑處理對青藏薹草種皮的影響不同,與沒有經過化學試劑處理的青藏薹草表皮,即與圖3A和圖3B對比分析,H2SO4,NaOH,NaHClO對果皮均有不同程度的機械破壞作用,并且使種皮明顯變薄(圖3D、圖3F、圖3H)。
其中49% 的H2SO4處理10 min,種皮表面己出現較明顯的酸蝕洞(圖3C),對柵欄層也有一定程度的破壞,并且使青藏薹草種皮最外層細胞出現大量的裂孔,種皮厚度約67.5 μm(圖3D)。
40% NaOH溶液處理30 min對種皮的機械破壞效果較好,不但去掉了薹草種子角質層(圖3E),而且使部分最外層大型厚壁細胞脫落,種皮厚度約79.4 μm(圖3F)。
10%的NaHClO溶液處理1 h對青藏薹草種皮的腐蝕較為明顯,去掉了種子角質層(圖3G),甚至去掉了種皮最外層的大型厚壁細胞,種皮厚度約68.5 μm,并且使中間縱向致密細胞層變得更加疏松(圖3H)。
3 討論
3.1 溫度與光照對種子萌發的影響
種子的萌發過程是種子的胚在內部多種酶的催化作用下發生的一系列物質轉化和能量傳遞過程[16],在這個過程中與酶的活性息息相關的就是溫度,酶保持最大活性需要適宜的溫度,溫度過高和過低都會降低酶的活性,進而體現在種子的萌發過程中[17]。本研究表明,HB種子萌發率最高的溫度是變溫15℃/25℃,而PME種子萌發率最高的溫度條件是15℃/30℃,這說明同一環境中的不同種莎草科植物種子具有不同的萌發適宜溫度,在自然環境中草種的萌發具有時空分異性;而且兩種種子在變溫條件下的萌發率均遠高于恒溫條件下,與其他植物種子的萌發試驗結果相似[18-23]。變溫可加快酶的催化作用,促進貯藏物質的轉化,其次變溫有利于種殼軟化,使種胚利于突破種皮,促進萌發[24]。
光照是影響種子萌發的諸多環境因素之一,光照誘導種子萌發的作用是作為指示萌發適宜環境的信號來終止種子休眠[25],光照對種子萌發的影響存在差異,有研究顯示光照可能促進[19]、抑制[26]或者不影響[27]種子的萌發過程。而本研究發現光照對HB種子的萌發是必需的,但是對PME種子的萌發幾乎沒有影響,說明HB為萌發喜光性種子,而PME為萌發光不敏感性種子,在HB種子的萌發中要注意光照條件。
3.2 冷層積處理對種子萌發的影響
冷層積是一種有效的解除種子休眠的方法,冷層積處理可降低種子中的抑制物質含量,使種皮軟化、提高透性、種胚發育后熟,解除種胚生理抑制作用引起的生理休眠型[28-30],對于HB種子來說,冷層積處理能顯著提高HB種子的萌發率,并顯著降低萌發時滯天數;PME種子在冷層積30天之后萌發率甚至顯著下降,而更長時間的冷層積會在一定程度上提升萌發率,冷層積同樣提升了QZ種子的萌發,冷層積90天之后QZ種子在培養期內出現了種子萌發。這說明HB,PME和QZ種子均存在一定程度的生理休眠現象。
種子的萌發時滯天數越低,說明其發芽越迅速。具有較低的萌發時滯天數的物種能在適宜的條件下迅速發芽,這直接關系著該物種繁衍、種群維持和發展[31-34]。本研究表明,冷層積對供試高寒草甸植物HB,PME,QZ種子萌發特性的影響因草種不同而異。HB種子和PME種子經冷層積處理后,萌發更迅速,且冷層積時間越長,萌發時滯天數越低,發芽越整齊。這也與魚小軍等對高寒草甸植物種子萌發特性的研究結果一致[32]。
3.3 化學試劑對種子萌發的影響
硬實是植物種子中普遍存在的現象,硬實種子是由于種皮堅硬而很難進行影響吸脹萌發的種子[35-36]。在本研究中,化學試劑處理降低了HB和PME種子的萌發率和萌發時滯,顯著促進了QZ種子的萌發過程,而且以QZ種子為例,結合電鏡觀察,可以發現一定濃度的H2SO4,NaOH,NaHClO均能不同程度的破壞種子木質堅硬的種皮結構和種皮最外層大型厚壁細胞,增大種皮細胞間隙,使中間縱向致密組織變得疏松,減輕其機械障礙和透性障礙,讓水分和氧氣易于進入種子,有利于種子與外部之間的物質交流,從而解除種子硬實外皮的物理休眠,提高其發芽率[13]。
有研究指出,化學處理聯合應用冷層積處理,可成功打破部分莎草科植物種子和其它植物種子的休眠,提高其萌發率[15,37-38],張國云等研究也表明在不考慮品種因素的情況下,NaOH溶液處理結合冷層積處理后平均發芽率最高[13]。本研究也表現為種子采用化學試劑浸種+冷層積綜合處理措施后萌發率比常規化學試劑處理和僅低溫層積處理的種子更高,且萌發也更迅速。
本研究中,QZ種子萌發最困難,溫度、冷層積和化學試劑處理對QZ種子萌發的影響并不顯著,可能與QZ種子中的內源抑制物含量過高或者基因調控有關[39-41]。因此,QZ種子的休眠和萌發特性以及如何進一步提高QZ種子的發芽率還有待進一步深入研究。
綜上所述,青藏高原莎草科種子在自然條件下的萌發率低,一方面是其生長的地區環境惡劣,植物的適應性生活策略所致[42];另一方面是莎草科種子小、種皮硬實、外覆光滑且較厚的角質層導致種皮對水分和氧氣的不透性,限制種子的萌發。種皮硬實有利于收獲和儲藏、抵抗逆境,但不利于雜草防治和種子繁殖檢驗與加工[43-44]。大量研究表明,牧草種子硬實可以通過物理方法和化學方法有效的破除[45-49],本研究結果表明采用化學試劑浸種+冷層積綜合處理措施比常規化學試劑處理和僅低溫層積處理的種子萌發率更高、萌發更迅速,并結合掃描電鏡發現化學試劑處理對種皮均有不同程度的機械破壞作用,使種皮明顯變薄,說明化學試劑H2SO4,NaOH,NaHClO均能有效破除莎草科種子硬實。這對特殊地域植被種子的萌發具有一定的理論參考價值。
4 結論
同一區域內采集的不同莎草科種子適宜萌發的溫度并不一樣,而且變溫條件下種子的萌發率顯著高于恒溫條件下的萌發率;光照對種子萌發的影響因物種而異,華扁穗草為萌發喜光性種子、帕米爾薹草為萌發光不敏感性種子。一定濃度的硫酸(H2SO4)、氫氧化鈉(NaOH)和次氯酸鈉(NaHClO)溶液通過破壞種皮最外層大型厚壁細胞,增大種皮細胞間隙,促進水分和氧氣進入種子,從而增加萌發率。冷層積處理或化學試劑+冷層積綜合處理能顯著提高華扁穗草、帕米爾薹草和青藏薹草種子的萌發率。所以在促進種子萌發的技術中,應綜合多種技術聯合處理,可以較好地促進種子的萌發。
