若爾蓋高寒沙化地中伴生草種對杯腺柳凋落葉初期分解的影響
2023-01-03 10:18:12周春梅陳冬明鄧東周胡興鵬鄧竹辛周冀瓊孫飛達白彥福韓金鋒鄢武先
草地學報 2022年12期
關鍵詞:影響
李 沛, 周春梅, 陳冬明, 李 其, 鄧東周, 黃 倩, 胡興鵬, 鄧竹辛, 周冀瓊, 孫飛達, 白彥福, 韓金鋒, 孫 庚, 鄢武先*, 劉 琳*
(1.森林和濕地生態恢復四川省重點實驗室, 四川 成都 610081; 2. 四川農業大學草業科學與技術學院, 四川 成都 611130; 3. 中國農業大學草業科學與技術學院, 北京 100193; 4. 四川若爾蓋濕地國家級自然保護區管理局, 四川 阿壩 624500; 5.中國科學院成都生物研究所, 四川 成都 610041)
若爾蓋高原地處青藏高原東南緣高寒半濕潤區,是我國主要牧區之一,也是黃河、長江上游水源涵養的重要生態功能區和生物多樣性重要保護地,在區域經濟社會可持續發展、維護國家生態安全和藏區長治久安中具有重要戰略地位[1-4]。近年來,受自然因素與人為因素的綜合影響,若爾蓋高原沙化面積逐年擴大。2010年,若爾蓋高原已出現3 470 km2沙化草地,占整個區域9.09%的面積,且近30年里,以每年0.007 km2的速率持續擴增[5],生態系統健康和服務功能嚴重下降。種植適生灌木[6-8],利用灌木的“資源島效應”來改善土壤條件和局部小氣候,為此后定居的草本植物提供保育作用,是全球沙化地治理常見方法[9-10]。在灌叢的遮擋和截獲作用下,草本植物種子可以更好地聚集,并利用灌叢冠幅下土壤富集的養分和更溫和的局部小氣候,提高草本植物定居成功率和促進草本層物種多樣性恢復[11]。自上世紀90年代以來,一系列生態治理工程在若爾蓋高原實施,許多鄉土灌木被廣泛應用于高寒沙化草地的生態恢復。若爾蓋高寒草甸區的沙化地植被恢復實踐已證明,杯腺柳(Salixcupularis)因具有很強的適應性[12],可改善高寒沙化土壤特性[13],且與草本配合種植可使沙化地土壤有機質和總氮含量顯著升高[14]。在高寒沙化地中,以杯腺柳為護理植物,選擇適宜的鄉土草本作為其伴生植物,可進一步加快區域沙化地植物物種多樣性和土壤生態功能的恢復。
植物凋落物分解影響著土壤有機質組成、養分濃度和土壤CO2排放[15],是陸地生態系統物質循環過程十分關鍵的環節。一方面,溫度和水分等環境因素對凋落物分解有十分重要的影響[16]。此外,不同植物種類及器官來源的凋落物由于化學組分不同,其分解特性存在一定差異[17],凋落物C、N含量、纖維素含量、木質素含量以及C/N和木質素/N等一系列指標常用于凋落物質量評價。高N凋落物比低N凋落物的分解速率高[18]。凋落物在分解過程中,當凋落物C/N值>25時發生氮固持,進而導致剩余凋落物中氮濃度增加;而當C/N值<25時凋落物氮開始礦化,釋放速率隨之加快[19]。木質素是植物凋落物中相對穩定的成分,在凋落物分解初期其濃度(質量百分率,%)常表現為增加,而到凋落物分解后期緩慢下降,是凋落物分解過程中主要的限速組分[20]。此外,伴生草種凋落物與杯腺柳凋落葉會產生混合凋落物分解的效應。與單種凋落物相比,混合凋落物具有更豐富的結構,凋落物的大小、幾何結構以及持水力之間的差異使分解的異質性增大,這種異質性的增加能夠使分解者具有更豐富的取食選擇。而且,一些土壤動物對多樣性高的凋落物更敏感[21]。此外,由于混合凋落物具有的營養元素更完整,各種營養元素通過淋溶作用在不同凋落物之間進行轉運,使微生物群落能夠更高效的利用碳源底物,從而抵消單種凋落物分解的營養限制[22]。有證據表明,由于凋落物之間的非加性相互作用,觀察到的總凋落物質量損失和營養礦化率很大程度不同于組分中單種凋落物分解總和的預期值[23-25]。而且,植物凋落物的投入數量對C和養分循環有很強的影響[26-28]。
退化生態系統恢復的最終目的,是建成一個新的自我支持生態系統,不需要持續人工輸入物質和能量,使生態系統具有一定的抗干擾能力[29]。因此,選擇適宜的植物及物種搭配,加快凋落物分解及釋放養分,促進土壤特性改善和物質循環的恢復,對于退化生態系統恢復意義重大。我們的前期研究發現,若爾蓋高寒草甸區沙化地中杯腺柳凋落葉分解特征存在著空間異質性,即凋落葉質量損失率在裸地高于在杯腺柳冠幅下[30]。在杯腺柳與草本配合種植的治沙模式中,不同伴生草本對杯腺柳凋落葉提供添加物的數量和質量均不一樣,直接影響著杯腺柳凋落葉的分解;不同伴生草本不同的存活率和地上生物量積累必會引起植被蓋度的差異,造成凋落葉分解微生態環境條件的差異,間接影響著杯腺柳凋落葉的分解。因此,選擇杯腺柳的適宜伴生草本植物,研究不同伴生草種對杯腺柳凋落葉分解的影響,探明這些伴生草種凋落物與杯腺柳凋落葉的混合分解反應和養分釋放規律,找出有利于杯腺柳凋落葉養分回歸的伴生草種,有助于提高高寒沙化地植被恢復的效率,加快高寒沙化地可自我支持的自然生態系統的重建,減少生態治理過程后期物質和能量的投入。
1 材料與方法
1.1 研究區概況
試驗樣地位于四川省阿壩州紅原縣瓦切鄉(33o10′ N,102o37′ E,海拔3 366 m)。該區域為大陸性高原寒溫帶季風氣候,春秋短促、長冬無夏。年均降水量792.0 mm,降雨主要集中在5-10月,年均氣溫為0.9℃,年均積雪期為76 d,無絕對的無霜期。相對濕度71%,晝夜溫差大,冷暖變化劇烈。該地由于沙源、風口、氣候變化、人為因素等導致沙化嚴重,土壤均為沙土且土質疏松,原有植被均已退化。目前所種植的灌木主要是青藏高原特有的杯腺柳等柳屬灌木和沙棘(Hippophaerhamnoides)等,其中杯腺柳等柳屬灌木對沙化地適應良好。
1.2 試驗方法
1.2.1試驗設計 試驗為雙因素設計,以未播種伴生草種的杯腺柳植株為對照,選用3種伴生草本植物分別單播(3水平)和杯腺柳的不同微位置(3水平,即內、中、外圈)為處理因素。隨機選取12株自然生長、大小相近的杯腺柳植株,隨機分為3組,分別播種垂穗披堿草(Elymusnutans)、中華羊茅(Festucasinensis)和老芒麥(Elymussibiricus)三種伴生草本植物,每4株為一組重復。并以每株杯腺柳莖基為中心,根據每株杯腺柳冠幅的大小,分別在莖基周圍、冠幅半徑中點和邊緣處,設置內、中、外3圈。
凋落葉收集與處理:2016年10月,在研究區域人工收集當年新鮮杯腺柳凋落葉,帶回實驗室風干后,65℃烘至恒重,分裝于凋落葉網袋中。按照取樣次數為6次的設計,以杯腺柳植株內圈可滿足放置18個凋落葉網袋為前提,確定最小凋落葉網袋的大小為10 cm×10 cm。然后,為保證單位面積凋落葉初始重量相同及各網袋內凋落葉厚度與幾何大小分布一致,確定中圈和外圈的凋落葉網袋大小,分別為10 cm×20 cm和15 cm×20 cm,并準確稱取烘干重為1.33 g的杯腺柳凋落葉裝入10 cm×10 cm的網袋、2.67 g的杯腺柳凋落葉裝入10 cm×20 cm的網袋、4 g的杯腺柳凋落葉裝入15 cm×20 cm的網袋。為排除小動物活動的影響,凋落葉網袋孔徑均為1 mm×1 mm。
伴生草種的播種方法:2015年5月,根據各株杯腺柳的內、中、外圈的周長,按照相同的播種密度(即,6粒·cm-1),沿各圈環形播種伴生草本的種子。
杯腺柳凋落葉袋的布置:沿每株杯腺柳伴生草本的環形播種槽外緣,分別放置凋落葉網袋,為保證每圈的取樣次數均為6次,每圈均放置凋落葉網袋18袋。同時,以未播種伴生草種的4株杯腺柳(冠幅大小與播種實驗的植株接近)作為對照,同樣的方法放置凋落葉網袋。試驗共16株杯腺柳,冠幅體積平均為(805.799±44.751)dm3,3種規格的凋落葉網袋各54袋,共864袋。在放置凋落葉袋之前,清除試驗樣地的原有凋落物。2016年11月,將處理好的凋落葉網袋放入試驗地,水平鋪于地表,用竹筷和鐵絲固定。放置凋落葉網袋時,盡可能使網袋之間的距離保持均勻一致。
1.2.2樣品收取與分析 在樣品放置后的第4個月(2017年3月)進行第一次取樣,此后每3個月取一次樣,共取6次。為減小空間異質性影響,每次取樣的3個凋落葉網袋與灌木莖基中心相連的半徑之間保持120°夾角。每次取回的3個凋落葉袋去除外來雜物和表面泥沙后,均勻混合凋落葉,待風干后,置于烘箱中,65℃烘至恒重,測定凋落葉的殘留量。隨后使用旋風研磨儀(裝置0.5 mm篩)將樣品研磨成粉末,密封干燥保存,用于凋落葉碳、氮、纖維素、木質素的測量。同時,測定杯腺柳凋落葉的初始化學成分。
1.2.3指標測定 凋落葉質量(含灰分干重):65℃烘至恒重,稱量并記錄。凋落葉總有機碳:采用K2Cr2O7-H2SO4氧化外加熱法測定(GB7657-87)。凋落葉全氮:采用凱氏定氮法測定(LY/T1269-1999)。凋落葉纖維素、木質素:改進的范式酸性洗滌纖維法[31]。
1.2.4數據統計 凋落葉質量損失率、半分解時間和95%分解時間計算公式分別為:
質量損失率Lt(%)=100×(Mt-M0)/M0
50%分解時間t50%(a)=-ln(1-0.50)/k
95%分解時間t95%(a)=ln(1-0.95)/k
式中,Mt為t時刻凋落葉網袋內剩余凋落葉干重量(g),M0為凋落葉網袋內初始凋落葉干重(g);k為凋落葉分解速率,應用Olson指數衰減模型ln(Mt/M0)=-kt擬合求得。
采用SPSS 20.0和Canoco 5.0進行統計分析。采用重復測量方差分析(Repeated-measure ANOVA)檢驗分解時間、伴生草種和微位置對凋落葉分解的影響。方差分析前,對不滿足正態分布的數據進行Box-Cox變換處理。采用單因素方差分析(One-way ANOVA)分別檢驗不同微位置、伴生草種和取樣時間對凋落葉分解的影響,影響顯著的處理因素再采用LSD檢驗法進行多重比較。采用Canoco 5.0對2019年6月凋落葉分解特征數據進行主成分分析及作圖。
2 結果與分析
2.1 伴生草種對凋落葉質量損失率的影響
對照和各草種伴生下,杯腺柳凋落葉質量損失率均隨時間延長顯著升高(P<0.05,表1)且均在582天時質量損失率最高(圖1)。而且,在垂穗披堿草伴生下,內圈和中圈的凋落葉質量損失率均分別比對照、中華羊茅和老芒麥伴生下相應的微位置更高。在第一個秋冬交替期(2017年9月到12月,即308天到398天),三種草本植物伴生下的凋落葉質量損失率出現整個分解試驗中的最大增幅。在214天和582天,在對照和中華羊茅伴生下,中圈和外圈的質量損失率均顯著高于內圈(圖1A,1B,P<0.05)。此外,垂穗披堿草伴生下,各微位置的杯腺柳凋落葉的50%和95%分解時間均最短,而且其伴生下中圈杯腺柳凋落葉分解時間最短(表2)。

圖1 不同伴生草種對凋落葉質量損失率的影響Fig.1 Effect of different companion grass species on the loss rate of litter mass注:*表示在0.05水平上差異顯著Note:* Indicates significant difference at the 0.05 level
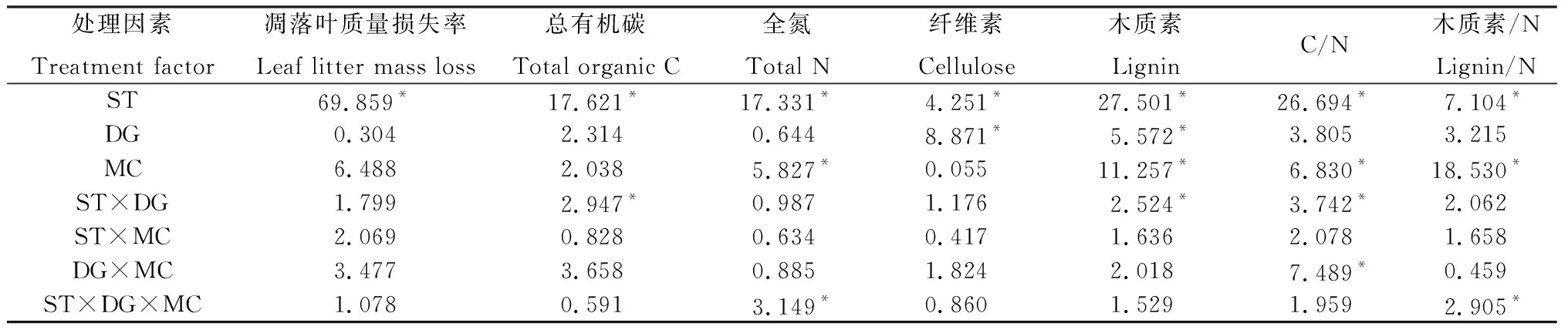
表1 杯腺柳凋落葉質量、養分殘留率和化學計量比的重復測量方差分析結果(F值)Table 1 The results of repeated-measure ANOVA on leaf litter quality,nutrient residual rate and stoichiometric ratio (F value)

表2 不同伴生草種下杯腺柳灌叢不同微位置杯腺柳凋落葉的分解特征Table 2 The decomposition characteristics of leaf litter at different microsites under the canopy of Salix cupularis with different companion grass species
2.2 伴生草種對凋落葉總有機碳和總氮含量變化的影響
分解時間及其與伴生草種交互效應顯著地影響著杯腺柳凋落葉總有機碳含量(P<0.05,表1)。在第398天,在中華羊茅和垂穗披堿草伴生下,內圈的凋落葉總有機碳含量均顯著高于中圈和外圈(P<0.05,圖2B,2C)。在第582天,垂穗披堿草伴生下,各微位置的凋落葉總有機碳含量均比初始含量顯著降低(P<0.05,圖2C),而在對照、中華羊茅和老芒麥伴生下,外圈的凋落葉總有機碳含量均顯著高于中圈和內圈,且均高于其初始含量(P<0.05,圖2B,2C,2D)。

圖2 不同伴生草種對凋落葉碳氮養分濃度變化的影響Fig.2 Effect of different companion grass species on the concentration of carbon and nitrogen nutrients in litter注:*表示在0.05水平上差異顯著Note:* Indicates significant difference at the 0.05 level
杯腺柳凋落葉全氮含量受分解時間、微位置及它們與伴生草種交互效應的顯著影響(P<0.05,表1)。隨著分解時間延長至第582天,各草種伴生下的各微位置凋落葉全氮含量比初始含量顯著上升(P<0.05)。而且,此時,在杯腺柳冠幅下(內圈和中圈),與無伴生草種的對照相比,垂穗披堿草伴生下的凋落葉全氮含量均顯著更高(P<0.05)。從第308天到398天,各草種伴生下的各微位置,凋落葉總有機碳含量和全氮含量幾乎都顯著升高,而且此后凋落葉全氮含量保持較之前更高的水平。
2.3 伴生草種對凋落葉纖維素和木質素分解的影響
伴生草種和分解時間對杯腺柳凋落葉纖維素含量均影響顯著(P<0.05,表1)。在整個分解試驗期間,凋落葉纖維素含量先緩慢上升,然后在第一個生長季(即2017年6月至9月,即第214天到308天)顯著降低,隨后的秋冬交替期間(即2017年9月至12月,即第308天到398天)其含量顯著升高。在第308天,除對照的內圈外,各微位置凋落葉纖維素含量均處于整個試驗期間的最低值,而且在垂穗披堿草伴生的中圈和外圈下,凋落葉纖維素含量均顯著比其它兩種伴生草種和對照低(P<0.05,圖3B,3C,3D)。

圖3 不同伴生草種對凋落葉纖維素和木質素分解的影響。Fig.3 Effect of different companion grass species on the cellulose and lignin decomposition of litter.注:*表示在0.05水平上差異顯著Note:* Indicates significant difference at the 0.05 level
凋落葉木質素含量受分解時間、微位置及它們與伴生草種交互效應的顯著影響(P<0.05,表1)。到第398天,在各草種伴生下的各微位置,凋落葉木質素含量均無顯著變化,此后在2017年12月至2018年3月(即第398天到487天)的冬春交替期,其含量均顯著升高。分解至582天時,凋落葉木質素含量均顯著高于初始濃度(P<0.05),而且垂穗披堿草伴生下,內圈的凋落葉木質素含量高于對照和其它兩種草本伴生下的內圈(圖3F,3G,3H)。
杯腺柳凋落葉的C/N、木質素/N均受分解時間和微位置的影響顯著(P<0.05,表1)。隨分解時間延長,凋落葉的C/N降低(P<0.05),且垂穗披堿草和老芒麥伴生下,凋落葉C/N幾乎一直低于25。分解至582天,三種草本伴生下,除對照、中華羊茅和老芒麥伴生的外圈以外,其余微位置的凋落葉C/N均較初始值顯著下降(P<0.05),且垂穗披堿草伴生下各微位置的C/N無顯著差異,而在對照、中華羊茅和老芒麥伴生下,外圈的凋落葉C/N均顯著高于冠幅下(即內圈和中圈)。分解至308天,在對照和各草種伴生下的各微位置,凋落葉木質素/N均無顯著變化,而在第一個秋冬交替期(即從308天到398天),垂穗披堿草和中華羊茅伴生下,凋落葉木質素/N均顯著降低至整個試驗期間的最低值,且均顯著低于初始濃度(P<0.05),此后冬春交替期(即從398天到487天),各處理下的木質素/N均顯著升高。
2.4 伴生草種對杯腺柳凋落葉分解空間差異的影響
在主成分分析結果中(圖5),其PC 1與PC 2分別解釋了所有變量變異的48.31 %和33.75 %,總解釋率為82.06%。分析結果表明,在分解第582天(即2018年6月),杯腺柳凋落葉質量損失率、總氮和木質素含量兩兩互為正相關,凋落葉總有機碳、C/N和木質素/N也兩兩互為正相關;并且,垂穗披堿草集群在PC1軸左邊,與中華羊茅和老芒麥明顯分離,且凋落葉質量損失率、總氮和木質素含量也在PC1左側。這說明,在垂穗披堿草伴生下,杯腺柳凋落葉質量損失率較高,且凋落葉總氮含量和木質素含量也更高,即與中華羊茅和老芒麥相比,在垂穗披堿草伴生下,杯腺柳凋落葉的分解更快,且在分解的第582天,其凋落葉的質量損失率、總氮和木質素含量均最高。

圖4 不同草種伴生的杯腺柳灌叢下其凋落葉C/N和木質素/N的變化Fig.4 Changes of C/N and lignin/N in litter under the canopy of Salix cupularis associated with different companion grass species注:*表示在0.05水平上差異顯著Note:* Indicates significant difference at the 0.05 level

圖5 不同草種伴生下凋落葉分解各指標的主成分分析Fig.5 Principal-component analysis of all variables associated with different companion grass species注:橫坐標和縱坐標分別代表第一(PCA axis 1)和第二(PCA axis 2)主成分值。DC,凋落葉總有機碳;DN,凋落葉總氮;MLR,凋落葉質量損失率;○表示中華羊茅(FS)伴生,□表示垂穗披堿草(EN)伴生,◇表示老芒麥(ES)伴生;黃色代表灌叢內圈(C),綠色代表灌叢中圈(M),藍色代表灌叢外圈(I)Note:DC,Total organic carbon;TN,Total nitrogen;MLR,Mass loss rate;○ denotes microsites under the S. cupularis associated with F. sinensis,□ denotes microsites under the S. cupularis associated with E. nutans,◇ denotes microsites under the S. cupularis associated with E. sibiricus;Yellow represents canopy center (C),Green represents midpoint of canopy radius (M),Blue represents canopy edge (I)
3 討論
3.1 不同伴生草種對杯腺柳凋落葉分解的影響
3.1.1不同伴生草種與杯腺柳的混合凋落物的分解反應 本研究表明,相對于另外兩種草本植物,垂穗披堿草作為杯腺柳的伴生草種,在混合凋落物分解初期更加有利于其分解。這可能是由于垂穗披堿草凋落物的數量和質量都更優。一方面,我們的前期研究表明,在若爾蓋高寒草甸區以杯腺柳為建群種的沙化地植被中,作為伴生草本植物,垂穗披堿草比中華羊茅和老芒麥有更高的發芽率[32],據此推測垂穗披堿草可形成初級生產量更高的植物群落,進而提供更多的凋落物,因此可促進杯腺柳凋落葉更快分解。另一方面,高質量的凋落物比低質量的凋落物具有更高的N濃度和更低的C/N,其分解速率也相對更大[33]。當質量差異明顯的凋落物混合時,可減少營養元素的淋溶損失[34],促進真菌菌絲體從周圍環境的養分吸收,從而提高養分的固持和減少養分的釋放,使得高質量凋落物組分促進了其它較低質量凋落物組分的分解[35]。本試驗中,來自不同伴生草種的凋落物對混合凋落物分解作用的總體表現為分解和周轉速率加快,而且垂穗披堿草(或老芒麥)混合杯腺柳凋落葉的C/N均一直低于25,以垂穗披堿草伴生對杯腺柳凋落葉分解的促進作用最為明顯。混合凋落物分解產生混合效應與凋落物的物種組合密切相關[36]。張麗莉[37]在對15種常見青藏高原高寒草甸植物凋落物分解的研究中顯示,垂穗披堿草具有較高的初始N濃度與較低的C/N,且與其他高原草種呈現顯著差異,這說明與其他營養成分含量偏低的高寒草甸植物相比,垂穗披堿草凋落物作為養分元素較高的高質量凋落物,可更有效地促進混合凋落物分解。本研究也證明,與中華羊茅和老芒麥相比,垂穗披堿草和杯腺柳的混合凋落物中,N濃度更高,且C/N更低,這可能使杯腺柳凋落葉中的微生物能固定的N源更豐富,可利用養分增加[38-40],從而更好地促進杯腺柳凋落葉初期的分解和養分釋放。
3.1.2伴生草種根系對杯腺柳凋落葉分解的影響 在本研究試驗中,我們觀察到三種伴生草本的根系均在杯腺柳凋落葉網袋中有分布。我們前期的研究表明,在若爾蓋高寒沙化地中,垂穗披堿草比另外兩種草本植物發芽率更高,可推測其根系也會發育得相對更好。根系從凋落物層中獲取養分更有效率,因此凋落物輸入常促進根系從土壤向凋落葉層中轉移[41]。而根系在凋落物層生長會對凋落物分解產生復雜的影響[41-42](圖6-③)。細根通過改變根際微生物的活力[43]和覓食行為[44-45]來對地上凋落葉的產量和分解產生直接或者間接的影響。伴生植物根系的激發效應、對礦質氮的吸收、以及與其共生的真菌,均能加速凋落物的分解[46]。根系激發效應對凋落葉分解的影響體現在兩個方面,一是根系分泌物能夠激活或改變微生物群落的組成[47],從而加速或抑制凋落葉的分解(圖6-②);二是根系死亡后作為高能量碳源,會促進微生物對舊有機物質的分解。根際激發效應對凋落葉分解可能產生正效應,也可能產生負效應。正的根際激發效應可促進有機質礦化增加達3.8倍,而負的根際激發效應會減少50%的有機質礦化[48]。而且,不同植物根系分泌物的化學組成成分常存在明顯差異。已有研究發現,19份不同基因型擬南芥(Arabidopsisthaliana)的根系分泌物化學成分差異明顯[49-50]。由此可以推測,與另外兩種草本植物相比,垂穗披堿草的根系分泌物和根系凋落物的數量和質量可能更有利于混合凋落物分解。此外,根系激發效應與氮的有效性相關[51],而氮的有效性也是影響凋落物分解的重要因素。本研究也表明,分解至2018年6月(即第582天)時,杯腺柳和伴生草本混合凋落物的全氮濃度均較初始增加,且均高于沒有伴生植物影響的單一杯腺柳凋落葉全氮濃度,這可能是由于分解前期,微生物固持氮而使凋落物中氮濃度升高,而且在有草本植物伴生的情況下,草本植物根系進一步促進了凋落物的微生物發育。

圖6 高寒沙化草地中伴生草對灌木凋落葉分解的影響途徑Fig.6 Effects of companion herbs on the decomposition of shrub leaf litter in the desertified alpine meadow注:①伴生草地上凋落物輸入量;②進入灌木凋落物袋的草本根系分泌物;③進入灌木凋落物袋的草本根系凋落物;④伴生草本減弱凋落物袋溫濕度變幅;⑤伴生草本削弱凋落物袋紫外線強度Note:①The leaf litter input of companion herbs;②Root exudates of companion herbs in the shrub litter bag;③Root litter of companion herbs in the shrub litter bag;④Attenuated variations of air temperature and humidity of shrub litter bag;⑤Reduced intensity of UVB exposure to the shrub litter bag
3.2 灌叢下不同微位置凋落物分解的差異
伴生草本植物通過提高植被蓋度,間接影響著杯腺柳凋落葉層的光照、溫度、濕度等微生態環境條件(圖6-④)。在生長季,伴生草種的葉層與杯腺柳灌叢相互遮蓋,阻滯空氣流通,使冠幅下微環境熱量更加不易損失,緩減群落下微環境的溫度波動。同時,通過減緩表層土壤與凋落葉層之間的晝夜溫差,創造更為溫潤的環境條件,影響土壤和凋落葉層的微生物活性,從而對凋落葉的分解產生間接影響。同時,不同伴生草種形成的地上植被蓋度和生物量存在差異,因此對微環境條件波動的緩減效果不同(圖6-①)。我們的前期研究也發現,在此區域,與中華羊茅和老芒麥相比,垂穗披堿草的出苗率顯著更高[32],因此垂穗披堿草形成的伴生草本層蓋度和地上生物量更高,更有效地緩解群落下微環境條件的波動,有利于凋落物層的微生物發育,進而更好地促進杯腺柳凋落葉分解。此外,在青藏高原高寒沙化地中的混合凋落葉分解初期,與生長季相比,非生長季(秋冬交替期和冬春交替期)的凋落葉分解動態也是劇烈和關鍵的。特別是在非生長季,各微位置之間凋落葉特征指標更多地表現出顯著差異。這可能是由于在非生長季,與裸地和杯腺柳灌叢的外圍相比,杯腺柳的莖基周圍常積累更厚的雪層。已有研究表明,雪被可改變土壤的凍融狀態(溫濕度條件)、土壤微生物和土壤動物的區系組成[52-53],進而影響季節性雪被分布區的凋落物分解。
其次,伴生草本的遮擋常減弱凋落葉層接受到的光強,并改變其光質,而紫外線顯著地影響著凋落物分解,其作用機制包括UVB的光降解和UVB導致分解微生物活性降低[54](圖6-⑤)。由于伴隨海拔的升高,空氣逐漸變稀薄,UVB輻射強度一般也逐漸增強。有觀測表明,正午時,與海拔2 300 m的西寧相比,海拔3 200 m的海北UVB輻射強度高10%~15%[55]。本試驗中研究樣地位于青藏高原東部,且海拔在3 300 m以上,同屬于UVB輻射高強度地區,UVB深刻影響著高寒沙化地的凋落物分解。因此,杯腺柳灌叢下不同微位置紫外輻射強度的差異也會影響凋落物分解。本研究也發現,微位置顯著地影響著杯腺柳凋落葉的總氮、木質素含量、C/N和木質素/N。特別是在無伴生草本的情況下,外圈的杯腺柳凋落葉質量損失率顯著高于內圈。說明,外圈更強的UVB輻射更有利于凋落葉的光降解。此外,UVB輻射通過在生長階段改變植物組織的營養組分來影響凋落物分解,例如,增加纖維素、木質素濃度[54],減少N濃度[57]等。李傳龍[58]研究表明,生長在較弱UVB輻射環境下的垂穗披堿草凋落物質量趨于較高水平,即C/N低、木質素/N低。本試驗也觀測到,在垂穗披堿草為伴生草種時,分解第19個月時,在內圈和中圈(即冠幅下)的杯腺柳凋落葉的質量損失率明顯高于外圈,這可能是因為冠幅下較弱的UVB輻射使垂穗披堿草提供了更高質量的凋落物添加,并且這種正效應影響超過了外圈更強烈的UVB光解正效應。
4 結論
綜上所述,在若爾蓋高寒沙化草地中,垂穗披堿草做為杯腺柳的伴生草種,比中華羊茅和老芒麥更有利于杯腺柳凋落葉的分解,更適宜作為高寒沙化地優勢植物——杯腺柳的伴生植物,促進高寒沙化地生物多樣性等生態功能的進一步恢復。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
