不同雜糧多糖的體外抗氧化、益生和抗癌活性比較
2023-01-03 07:10:14楊斯惠馬明芳曹亞楠任遠航彭鐮心
食品工業科技 2023年1期
關鍵詞:苦蕎
楊斯惠,馬明芳,曹亞楠,任遠航,萬 燕,鄒 亮,彭鐮心,
(1.成都大學農業農村部雜糧加工重點實驗室,四川成都 610106;2.四川省雜糧產業化工程技術研究中心,四川成都 610106;3.青海省產品質量檢驗檢測院,青海西寧 810004)
雜糧是指除水稻、小麥、大豆、玉米和薯類作物以外的糧谷類作物[1],具有生育期短、種植面積小、地域性強、種植方法特殊等種植特點[2]。谷類雜糧主要包括大麥、小米、青稞、燕麥、蕎麥等;豆類雜糧主要有綠豆、蕓豆、蠶豆、豌豆等[3]。隨著人們對膳食結構多樣化和飲食均衡化重視的加深,人們逐步發現在調節飲食結構和均衡膳食方面,雜糧是日常飲食中不可或缺的部分。與功能活性含量較低的稻谷、小麥相比,種類繁多的雜糧營養成分含量更高[3],其富含膳食纖維、維生素、礦物質等營養物質[4],具有細糧不可替代的地位。我國作為雜糧生產大國,具有悠久的雜糧飲食文化,可雜糧產品主要集中于原糧,加工產品低端,如何充分利用雜糧生理功能活性物質是雜糧開發的方向之一。多糖物質(如β-葡聚糖、阿拉伯木聚糖、纖維素)作為雜糧營養組成的重要部分,對其功能活性深入研究于雜糧行業發展具有積極的意義。
多糖是由10個或10個以上的單糖經過糖苷鍵聚合、脫水形成的天然高分子化合物[5],主要分布于動物、植物、藻類及微生物中[6]。根據來源不同,多糖可以被分為動物多糖、植物多糖和微生物多糖[7]。作為多糖中的一種,廣泛存在于自然界植物體中的植物多糖具有多種生物活性,如抗氧化、抗腫瘤、抗肝損傷、降血糖、益生作用等[8-9]。多糖作為結構復雜的高分子化合物,其功能特性與結構、物理性質密切相關。因此,對多糖提取、分離技術的探究成為了研究多糖的鋪墊。目前,對多糖提取工藝的研究有熱水浸提、酸提法、堿提法、超聲波提取法等。其中,熱水浸提法是多糖中最常用的提取方法,具有經濟、便捷、能較好地保存分子結構的特點。
近年來,植物多糖在加工、結構和生物活性方面備受關注,成為越來越多科研工作者的研究熱點。其中針對雜糧多糖的研究也逐步增加。Hu等[10]通過DPPH法和ABTS法評估了柱層析純化后藜麥多糖的抗氧化能力,結果顯示,藜麥多糖對DPPH和ABTS自由基表現出顯著的抗氧化活性。Qian等[11]采用超聲輔助法提取大麥多糖,發現大麥多糖在總還原能力、清除DPPH自由基、羥自由基、超氧陰離子自由基等方面具有抗氧化活性。Sargautiene等[12]探索了燕麥非淀粉多糖潛在益生作用,發現地衣芽孢桿菌可以對燕麥非淀粉多糖進行預消化,降低其β-葡聚糖的高粘度,使其他細菌能夠更容易利用半纖維素。Lin等[13]發現不同分子量的青稞β-葡聚糖對結腸癌細胞HCT116的體外生長抑制作用無明顯差異,分子質量較低的青稞β-葡聚糖也具有較強的抗癌活性。雖然雜糧多糖的不同生物活性被逐步揭示,可目前研究更多集中于單種雜糧多糖的提取分離純化、理化特性及某種活性探究,由于提取、分析方法的差異,可比性不足,不利于雜糧的綜合評價。
本研究以燕麥、薏米、藜麥、糙米、黃小米、大麥、青稞、苦蕎、黑麥為原料,小麥作為對照,通過測定、比較10種多糖的抗氧化活性、抑制結腸癌細胞HCT116活性以及對益生菌生長的影響,綜合評價,篩選出活性強的多糖。以多糖為切入點,揭示其在雜糧中的功能作用,明確不同雜糧多糖的功效差異,為雜糧功能性產品的開發和雜糧多糖的合理利用提供理論依據,奠定基礎。
1 材料與方法
1.1 材料與儀器
小麥、燕麥、薏米、藜麥 贛州康瑞農產品有限公司;糙米、黃小米、大麥 福建盛耳食品有限公司;青稞 沈陽信昌糧食貿易有限公司;苦蕎 云南健爽科技有限公司;黑麥 淶水縣金谷糧油食品有限公司;耐高溫α-淀粉酶(活性5萬U/g)、高轉化率糖化酶(活性10萬U/g) 河南萬邦化工科技有限公司;中性蛋白酶(活性10萬U/g) 南寧東恒華道生物科技有限公司;長雙歧桿菌(ATCC 15707)、短雙歧桿菌(ATCC 15700)、青春雙歧桿菌(ATCC 15703)、鼠李糖乳桿菌(ATCC 53103) 明州生物科技有限公司(中國寧波);MRS培養基(不含葡萄糖) 山東拓普生物工程有限公司;TPY液體培養基 青島高科技工業園海博生物技術有限公司;HCT116細胞 中國科學院細胞庫;RPMI1640改性培養基、胎牛血清(FBS,Fetal bovine serum) Gibco公司;NaOH(片狀)、鹽酸、無水葡萄糖、硫酸、苯酚、L(+)-抗壞血酸(VC)、硫酸亞鐵、水楊酸 分析純,成都市科隆化學品有限公司;30% H2O2分析純,成都市科龍化工試劑廠;Trolox標準品(水溶性VE)、ABTS標準品、過硫酸鉀 上海麥克林生化科技有限公司;DPPH標準品 上海源葉生物科技有限公司。
FE28型pH計 梅特勒-托利多國際貿易(上海)有限公司;XMTD-7000恒溫水浴鍋 北京市永光明醫療儀器有限公司;FD-2型真空冷凍干燥機北京博醫康實驗儀器有限公司;Synergy HTX多功能微孔板檢測儀酶標儀 美國伯騰儀器有限公司;UPH-1-10T型超純水制造系統 四川優普超純科技有限公司;LDZX-50KB型高壓蒸汽滅菌鍋 上海中安醫療機構廠;DHP-9160B生化培養箱 上海瑯玕實驗設備有限公司;JJ-CJ-IFD超凈工作臺 蘇州市金凈凈化設備科技有限公司。
1.2 實驗方法
1.2.1 多糖的提取、分離 參考曾海龍[14]、王希[15]等的方法,略作修改。稱取100 g滅酶原料(原料米85 ℃烘1 h滅內源酶活,備用),采用熱水提取法,料液比為1:20(g/mL),在100 ℃浸提2 h,過濾取上清液。調節溶液至pH5.5,在溶液中加入0.5 g耐高溫α-淀粉酶,93 ℃水浴30 min;加入1 g高轉化率糖化酶,60 ℃水浴處理30 min。在水浴溫度55 ℃下,再加入2 g中性蛋白酶,反應30 min,再加熱至100 ℃滅酶10 min,收集酶解液。酶解液以1 mol/L NaOH液調至pH8.0,添加30% H2O2至淺黃色,于50 ℃下水浴加熱2 h。濃縮液體到原體積的1/4,向多糖溶液中逐漸加入4倍體積的無水乙醇,使體系中乙醇最終體積達到80%,4 ℃靜置24 h,離心取沉淀,冷凍干燥得粗多糖樣品。
1.2.2 多糖含量的測定 參考楊雅蛟等[16]的方法,略作修改。取干燥后的多糖樣品用蒸餾水配制成濃度0.2 mg/mL的多糖溶液,備用。以無水葡萄糖為對照品(回歸方程為y=5.6483x+0.0094,R2=0.9993),采用苯酚-硫酸法測定多糖含量。取1 mL多糖溶液,加入質量分數為5%的苯酚溶液1.0 mL,混合均勻后迅速加入濃硫酸5.0 mL,置于40 ℃水浴條件下反應30 min,將反應得到的溶液在波長490 nm處測吸光值。
1.2.3 體外抗氧化活性測定
1.2.3.1 DPPH自由基清除率的測定 取干燥后的多糖樣品用蒸餾水配制成濃度5 mg/mL的多糖溶液。取2 mL 5 mg/mL多糖溶液,加入2 mL DPPH溶液,混勻暗反應30 min,在波長為517 nm處測定吸光度A1;取2 mL 5 mg/mL多糖溶液與2 mL無水乙醇溶液反應測定吸光度A2;2 mL無水乙醇溶液與2 mL DPPH溶液反應測定吸光度A0[17-18]。按下式計算樣品的DPPH自由基清除率。以Trolox標準品為對照品,試驗結果以每1 g干重(DW)樣品中等量Trolox(μmol)表示:μmol Trolox/g DW。
DPPH自由基清除率(%)=[A0-(A1-A2)]/A0×100
1.2.3.2 ABTS自由基清除率的測定 參考李巨秀等[19]的方法,略作修改。取干燥后的多糖樣品用蒸餾水配制成濃度3 mg/mL的多糖溶液。取1 mL 3 mg/mL多糖溶液,加入4 mL ABTS溶液,混勻暗反應10 min,在波長為734 nm處測定吸光度A1;取1 mL 3 mg/mL多糖溶液與4 mL無水乙醇溶液反應測定吸光度A2;1 mL蒸餾水與4 mL ABTS溶液反應測定吸光度A0。按下式計算樣品的ABTS自由基清除率。以Trolox標準品為對照品,試驗結果以每1 g干重(DW)樣品中等量Trolox(μmol)表示:μmol Trolox/g DW。

1.2.3.3 羥自由基(·OH)清除率的測定 取干燥后的多糖樣品用蒸餾水配制成濃度3 mg/mL的多糖溶液。取2 mL H2O2(9 mmol/L)、FeSO4(9 mmol/L)、多糖溶液(3 mg/mL)混勻靜置10 min,加入2 mL水楊酸(9 mmol/L),混勻反應30 min,在波長為510 nm處測定吸光度A1;2 mL多糖溶液(3 mg/mL)與6 mL蒸餾水反應測定吸光度A2;蒸餾水代替多糖溶液測得對應吸光度A0[20]。按下式計算樣品的羥自由基清除率。以VC為對照品,試驗結果以每1 g干重(DW)樣品中等量VC(μmol)表示:μmol VC/g DW。
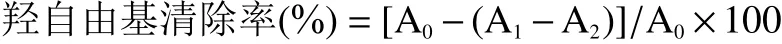
1.2.4 對益生菌生長的影響
1.2.4.1 培養基及培養條件 鼠李糖乳桿菌:MRS培養基,37 ℃恒溫培養過夜。
長雙歧桿菌、短雙歧桿菌、青春雙歧桿菌:TPY培養基,37 ℃恒溫厭氧培養過夜。
1.2.4.2 生長曲線的測定 選擇長雙歧桿菌、短雙歧桿菌、青春雙歧桿菌和鼠李糖乳桿菌,活化后(OD600nm=0.5)以2%(v/v)的比例接種于不添加碳水化合物的液體培養基中,混勻后吸取180 μL于無菌96孔板中,加入20 μL多糖溶液(1%、10%)共培養,每個孔中多糖的最終濃度為0.1%、1%(w/v),每孔200 μL,每組分別設置6個平行,置于37 ℃恒溫培養,每隔4 h檢測發酵液的OD600nm值,根據前期預實驗,選擇連續測定40 h,最后以時間為橫坐標,OD600nm為縱坐標,繪制微生物生長曲線,以無碳水化合物培養基為空白對照,以添加菊糖為唯一碳源的培養基為陽性對照[21-22]。
1.2.5 體外抑制結腸癌細胞HCT116的活性測定采用MTT法測定雜糧多糖對HCT116細胞增殖抑制作用。取對數生長期HCT116細胞,種板到96孔培養板中,每孔100 μL,置于CO2培養箱中培養過夜。吸棄培養基,加入含不同濃度多糖的培養基100 μL/孔,每個濃度設計5個復孔,將96孔板置于CO2培養箱中培養48和72 h[23-25]。對照組(陰性對照)只加細胞和新鮮培養基,空白組不加細胞只加培養液。按下式計算樣品的細胞抑制率。
細胞抑制率(%)=[(對照組細胞OD值-實驗組細胞OD值)/(對照組細胞OD值-空白組OD值)]×100
1.3 數據處理
所有實驗至少重復三次,采用Excel 2010軟件分析整理數據,數據采用平均值±標準差表示。采用SPSS 23.0統計軟件對數據進行統計和分析,P<0.05表示差異顯著;采用Origin 2018軟件作圖。
2 結果與分析
2.1 不同雜糧粗多糖的多糖含量
不同雜糧粗多糖的多糖含量如圖1所示,小麥、黑麥、青稞、黃小米、糙米、薏米、燕麥、大麥、藜麥、苦蕎中的多糖含量分別為37.11%、10.61%、56.74%、40.24%、45.97%、49.89%、45.61%、57.59%、42.83%、36.11%。多糖含量最高的為大麥粗多糖(57.59%),與之前王希[26]報道水提法提取的大麥多糖中多糖含量為64.29%相接近,本研究中過氧化氫脫色步驟可能導致部分多糖的損失[14];多糖含量最低的為黑麥粗多糖(10.61%)。
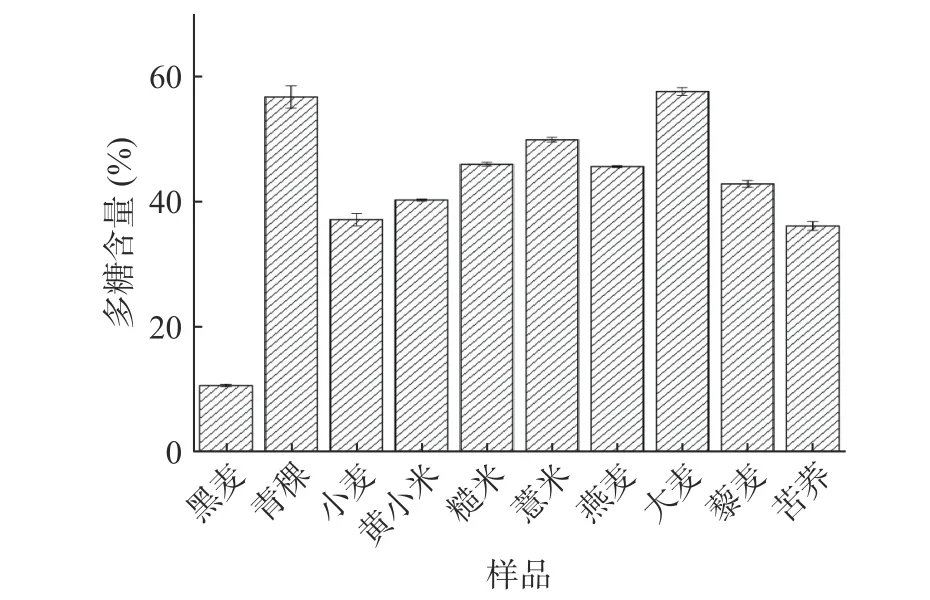
圖 1 不同雜糧粗多糖的多糖含量Fig.1 Polysaccharide content of crude polysaccharide in different coarse cereals
2.2 體外抗氧化活性分析
2.2.1 DPPH自由基清除能力 DPPH·(1,1-二苯基-2-三硝基苯肼)是一種穩定的自由基,呈紫色,其最大吸收波長為517 nm,被廣泛用于評價自由基清除活性[27-28]。如圖2所示,不同多糖對DPPH自由基清除能力具有顯著性差異(P<0.05)。10種雜糧多糖中,苦蕎多糖的DPPH自由基清除能力顯著高于小麥多糖,達12.76 μmol Trolox/g DW;而黃小米、薏米、燕麥等5種雜糧多糖均顯著低于小麥多糖(P<0.05),其中DPPH自由基清除能力最弱的是黃小米多糖。在結構研究方面,王帥等[29]利用最小偏二乘分析初步分析純化后6種多糖的結構、性質對其體外抗氧化活性的影響,提出單糖的組成(葡萄糖醛酸、半乳糖和葡萄糖)和分子量能對多糖抗氧化活性起到影響。已有報道中,苦蕎多糖主要由葡萄糖、半乳糖組成[30],小米多糖則由鼠李糖、阿拉伯糖、木糖、葡萄糖、半乳糖組成[31]。因此,苦蕎多糖和黃小米多糖清除DPPH自由基能力的差異可能是由于其單糖組成不同。除去單獨存在的多糖,多糖還可通過與蛋白質、多肽以及酚類化合物的結合向缺乏電子的自由基提供質子從而增強其抗氧化能[32]。故苦蕎多糖組清除能力強可能與多糖和酚類化合物的結合有關[33-34]。

圖 2 不同雜糧多糖的DPPH自由基清除能力Fig.2 DPPH radical scavenging ability of different coarse cereal polysaccharides

圖 3 不同雜糧多糖的ABTS自由基清除能力Fig.3 ABTS radical scavenging ability of different coarse cereal polysaccharides
2.2.2 ABTS自由基清除能力 在反應體系中,ABTS自由基與抗氧化物質結合使體系褪色,通過測定最大吸收波長734 nm下吸光度的變化評價物質的抗氧化能力[35]。不同雜糧多糖的ABTS自由基清除能力如圖3所示。在ABTS自由基清除能力上,不同多糖間具有顯著性差異(P<0.05)。10種多糖中苦蕎多糖顯著高于小麥多糖(P<0.05),達39.56 μmol Trolox/g DW,而黑麥、黃小米、糙米、薏米和藜麥多糖均顯著低于小麥多糖(P<0.05)。與DPPH自由基清除能力結果不同,ABTS自由基清除活性最弱的是薏米多糖(9.33 μmol Trolox/g DW)。復雜易變的分子結構和物理性質是造成多糖生物活性多變的重要原因。其中,單糖的組成和摩爾比是影響多糖的抗氧化活性的因素之一。孫元彬等[36]對純化得到的苦蕎多糖進行單糖組成分析,發現其只含有葡萄糖和少量的木糖。莊瑋婧[37]發現水提法得到的薏米多糖由L-鼠李糖、L-阿拉伯糖、D-葡萄糖、D-半乳糖、D-木糖、D-甘露糖六種單糖組成。因此,葡萄糖含量高可能是苦蕎多糖ABTS自由基清除能力強的原因。從杏鮑菇菇頭[38]中提取的主要由葡萄糖組成(占84.4%)的多糖對ABTS自由基具有較強的淬滅作用也印證了多糖的單糖組成與抗氧化活性具有相關性這一觀點。同時與DPPH自由基清除能力結果不同,在ABTS反應體系中,10種多糖清除率明顯增大,這可能與ABTS的親水親脂特性及較大空間位阻有關[39]。
2.2.3 羥自由基(·OH)清除能力 羥自由基是活性氧中最具活力的,具有很強的電子氧化能力,是最有毒害的自由基,可對相鄰的生物分子造成損害[40-41],加快生物體衰老,誘發疾病[42]。因此,清除羥自由基是體現多糖抗氧化作用的重要指標之一。不同雜糧多糖的羥自由基清除能力如圖4所示。不同雜糧粗多糖間羥自由基清除能力有顯著差異(P<0.05)。10種多糖中,苦蕎多糖的羥自由基清除能力顯著高于小麥多糖(P<0.05),達1615.32 μmol VC/g DW,而與DPPH、ABTS自由基清除能力結果不同,僅燕麥多糖活性低于小麥多糖。苦蕎多糖比燕麥多糖表現出更高的羥自由基清除能力,這可能是由于其糖醛酸含量較高,糖醛酸含量的增加能增進多糖的還原能力并減少羥自由基的產生[43]。據報道,苦蕎可溶性膳食纖維中含有糖醛酸26.10%[44],燕麥多糖含糖醛酸5.9%[45]。研究表明,糖醛酸含量較高(33.27%±0.10%)的硫酸化涼草粉多糖自由基清除效果更佳[46],這與之前Zhang等[47]的推測相似。同時,比較裙帶菜中不同分子質量巖藻多糖的抗氧化性發現,低分子質量(分子質量小于1×104u)巖藻多糖對羥自由基清除能力更佳[48]。但鑒于本研究中未進行多糖分子量測定,仍需進一步驗證。此外,苦蕎中富含黃酮類物質,李寧等[49]發現通過優化聯合提取工藝即水提醇沉法可以提高苦蕎籽中的黃酮和多糖的提取率。之前的研究證明,苦蕎黃酮粗提物具有清除羥自由基的能力,可隨著對黃酮粗提物的精制,清除羥自由基的效果有所下降[50],故苦蕎多糖表現出具有較強的抗氧化作用可能與黃酮的協同作用有關。
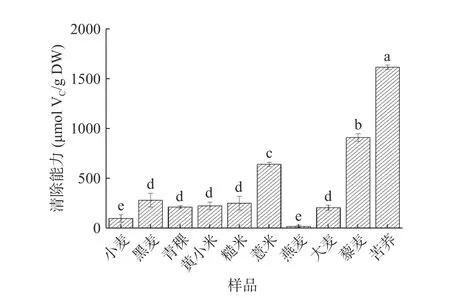
圖 4 不同雜糧多糖的羥自由基清除能力Fig.4 Hydroxyl radical scavenging ability of different coarse cereal polysaccharides
本研究通過三種方法對不同雜糧多糖的抗氧化活性進行測定。總體而言,苦蕎多糖表現出了良好的抗氧化能力。而不同多糖在DPPH、ABTS和羥自由基清除效果具有一定的差異。特別是小麥對照在清除DPPH、ABTS自由基上表現出優于部分雜糧多糖,但對羥自由基清除能力較弱。阿拉伯木聚糖,也稱為戊聚糖,作為一種非淀粉多糖廣泛存在于谷物中,特別在小麥籽粒中尤為豐富[51]。因其分子結構中具有一定的阿魏酸基團,阿拉伯木聚糖具有抗氧化活性[52]。在之前的報道中,具有較高的分子量和酯化阿魏酸的阿拉伯木聚糖被認為是較好的抗氧化劑[52-53]。Chen等[53]發現低取代度有益于阿拉伯木聚糖表現出更強的羥自由基清除活性,與之相反,阿拉伯木聚糖的DPPH自由基清除活性隨取代度的增加而提高,因為取代度越高的阿拉伯木聚糖更易分散到反應混合物中參與氧化還原反應。因此,對照小麥中阿拉伯木聚糖取代度較高可能是清除DPPH、ABTS自由基能力較佳,對羥自由基清除能力較弱的原因之一。
2.3 對益生菌生長的影響
本研究采用體外單菌培養方法對不同雜糧多糖的益生活性進行分析。不同雜糧多糖對益生菌的增殖具有菌株特異性(圖5)。在多糖濃度為0.1%時,除黑麥、青稞多糖外,其余8種多糖均能作為唯一碳源被長雙歧桿菌利用(圖5a);除黑麥、薏米多糖外,短雙歧桿菌能利用其余8種多糖生長增殖(圖5b);10種多糖對青春雙歧桿菌的促生長作用不明顯,均低于菊糖(圖5c);而鼠李糖乳桿菌則能利用10種多糖作為唯一碳源生長增殖(圖5d)。在多糖濃度為1%時,長雙歧桿菌不能利用10種多糖,培養40 h后發酵液OD600nm與陰性對照無糖培養基相近(圖5e),這可能與多糖的濃度具有一定關系,多糖濃度過高可能導致培養基滲透壓增加,從而導致菌體脫水,影響益生菌生長[54-55];黃小米和大麥多糖對短雙歧桿菌的益生活性高于菊糖,燕麥多糖與菊糖對短雙歧桿菌的益生活性相近(圖5f);黃小米和大麥多糖對青春雙歧桿菌的益生活性高于菊糖,燕麥多糖與菊糖對青春雙歧桿菌的益生活性相近(圖5g);鼠李糖乳桿菌能利用黃小米、燕麥、糙米和大麥多糖作為唯一碳源生長增殖(圖5h)。本研究結果顯示,濃度為1%時,黃小米、大麥、燕麥多糖能夠更易被短雙歧桿菌、青春雙歧桿菌和鼠李糖乳桿菌選擇利用,對照小麥多糖則表現出抑制4種益生菌增長的趨勢,同時不同雜糧多糖對益生菌的增殖作用不全是隨著濃度增加而增強。據劉麗莎等[56]報道,高濃度(添加量大于2.0%)的白術多糖可能引起滲透壓、pH改變及代謝物的積累等限制了雙歧桿菌的生長。同樣,張桂蘭等[57]也發現褐藻硫酸多糖濃度為2.0%~2.5%時對雙歧桿菌增殖作用最好;濃度大于5%時,雙歧桿菌數量無明顯增加。而來源不同的多糖對不同的益生菌作用不同,這也可能與其高度復雜的結構、物理特性密切相關[58],例如,水溶性高和黏度低的多糖被認為可以更快、更易被益生菌利用[59],因此還有待進一步研究證實。

圖 5 不同雜糧多糖對益生菌生長的影響Fig.5 The effects of polysaccharides from different coarse cereals on the growth of probiotics
2.4 體外抑制結腸癌細胞HCT116的活性測定
對10種多糖進行體外MTT增殖抑制試驗初篩抗腫瘤活性,其中小麥、黃小米、糙米、薏米和大麥多糖對HCT116細胞有一定抑制作用,其余5種多糖對HCT116細胞沒有明顯抑制作用。根據預實驗結果,選取5個濃度梯度,48、72 h兩個時間梯度,對具有抑制作用的5種多糖進行MTT試驗,如圖6、圖7所示,利用SPSS軟件分析計算得出其對 HCT116細胞的半數致死濃度 IC50,不同多糖之間 IC50具有顯著差異(P<0.05),如圖8所示。隨著對 HCT116細胞作用時間的延長,5種多糖抑制率也相應提高,表現出時間依賴關系。

圖 6 48 h下不同濃度小麥(a)、黃小米(b)、糙米(c)、薏米(d)、大麥(e)多糖對HCT116細胞的抑制活性Fig.6 Inhibitory activity of different concentrations of wheat (a), yellow millet (b), brown rice (c), adlay (d) and barley (e)polysaccharides on HCT116 cells at 48 h

圖 7 72 h下不同濃度小麥(A)、黃小米(B)、糙米(C)、薏米(D)、大麥(E)多糖對HCT116細胞的抑制活性Fig.7 Inhibitory activity of different concentrations of wheat (A), yellow millet (B), brown rice (C), adlay (D) and barley (E)polysaccharides on HCT116 cells at 72 h

圖 8 HCT116細胞上不同雜糧多糖的IC50值Fig.8 IC50 values of polysaccharides from different coarse cereals on the HCT116 cells
在48 h,小麥、黃小米、糙米、薏米和大麥多糖的IC50值分別為1.24、2.90、1.40、1.79、2.82 mg/mL。對比 IC50值大小說明 48 h 下不同多糖抑制 HCT116細胞增殖的活性強弱:小麥>糙米>薏米>大麥>黃小米。在72 h,小麥、黃小米、糙米、薏米和大麥多糖的IC50值分別為0.98、2.55、1.33、1.23、2.82 mg/mL。對比 IC50值大小說明 72 h 下不同多糖抑制 HCT116細胞增殖的活性強弱:小麥>薏米>糙米>黃小米>大麥。李彩嬌[8]曾證明大麥多糖對HT 29結腸癌細胞有抑制作用,且通過線粒體凋亡途徑誘導細胞凋亡,這與本研究中大麥多糖抑制HCT116細胞活性的途徑是否一致,可以作進一步研究。令人意外的是,48、72 h下5種多糖中對照小麥多糖抗結腸癌細胞活性最強。之前的報道中,Murtazina等[60]已證實通過培養小麥細胞提取獲得小麥細胞培養多糖(wheat cell culture polysaccharides,WCCPSs)具有抗HCT 116結腸癌細胞能力并提出從WCCPSs的單糖組成比例來看,葡萄糖的含量和葡萄糖:阿拉伯糖:甘露糖的比例可能是決定抗癌活性的主要因素。但小麥多糖具體如何表現出較高的抑制HCT 116細胞活性的原因和構效關系,有必要進一步深入探究。綜上,明確多糖結構與抗結腸癌細胞活性的構效關系以及多糖抑制結腸癌細胞增殖機制仍是需要廣大研究者們投入研究的關鍵。
3 結論
本研究以9種雜糧多糖為研究對象,小麥多糖為對照,對比了十種多糖的抗氧化活性、體外抑制結腸癌細胞HCT116的活性和對益生菌生長的影響。結果表明,不同雜糧多糖在三種體外活性上存在顯著性差異。十種多糖對DPPH、ABTS和羥自由基均具有清除能力。其中,苦蕎多糖對三種自由基的清除能力最強。在對益生菌生長的影響上,1%濃度的黃小米、大麥、燕麥多糖能夠更易被短雙歧桿菌、青春雙歧桿菌和鼠李糖乳桿菌選擇利用。在48 h下多糖抑制HCT116細胞增殖的活性強弱:小麥>糙米>薏米>大麥>黃小米,在72 h下多糖抑制HCT116細胞增殖的活性強弱:小麥>薏米>糙米>黃小米>大麥。本研究通過測定比較不同雜糧多糖的三種體外活性,明確不同雜糧多糖在體外活性上的差異,為雜糧多糖的開發提供理論依據,為其在食品、生物等不同領域的精準利用提供思路。
本研究僅對9種谷類雜糧進行了研究,然而雜糧種類豐富,品系龐雜,如何綜合評價不同種類雜糧多糖生理活性將是一重大挑戰。此外,本研究采用熱水浸提法提取不同雜糧中的多糖,而熱水浸提得到的大多是分子量較小的中性多糖[61],如采用酸提法、堿提法、酶解法等不同方法提取,得到的多糖結構不同,其在體外活性上的強弱可能會隨結構的改變而產生變化。本文未對各種雜糧多糖進一步的純化和鑒定,存在一定局限,但也為不同雜糧的活性差異提供重要參考。未來,可對活性較強的雜糧多糖進一步分離純化,探討其活性機制。
猜你喜歡
農產品加工(2023年14期)2023-08-17 10:32:28
農業科技與信息(2021年8期)2021-12-06 04:24:43
詩潮(2021年11期)2021-11-24 19:55:50
四川農業與農機(2021年1期)2021-01-05 19:15:18
青年歌聲(2018年5期)2018-10-29 03:18:40
現代食品(2018年21期)2018-02-14 20:51:24
食品安全導刊(2018年30期)2018-01-17 05:47:48
廣東飼料(2016年3期)2016-12-01 03:43:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
林業與生態(2016年2期)2016-02-27 14:24:01
