煙草NtNRAMP3b的克隆及功能分析
2023-01-05 07:48:28尹卓然軒棟棟李晨依李長柴哲王錕瑤趙孟琦彭靖媛董杰賈宏昉
生物技術通報 2022年12期
尹卓然 軒棟棟 李晨依 李長 柴哲,2 王錕瑤 趙孟琦 彭靖媛 董杰 賈宏昉
(1.河南農業大學煙草學院,鄭州 450002;2.陜西中煙工業有限責任公司,西安 710000)
鐵是植物生長發育所必需的一種微量元素,可以通過活化葉綠素合成相關酶、呼吸作用相關酶進而影響植物光合作用、呼吸等生理活動,最終影響植物的生長發育[1]。煙草是一種重要的葉用模式植物和經濟作物。煙草缺鐵時會出現葉片黃化[2],影響煙葉的產量和品質[3-4]。研究煙草對鐵的吸收與轉運可以有針對性的阻止缺鐵環境對煙草生長發育的危害,提高煙葉的產量和品質。
研究表明,廣泛存在于植物中的NRAMP 家族主要參與植物對鐵(Fe)、錳(Mn)和鎘(Cd)等金屬離子的吸收和轉運[5]。擬南芥(Arabidopsis thaliala)中已有6 個NRAMP 家族基因的功能被證實[6-10],其中AtNRAMP1參與Mn 和Fe 的吸收轉運[11],AtNRAMP3和AtNRAMP4參與Mn 和Fe 的再利用[12],且編碼蛋白位于液泡膜上。水稻(Oryza sativa)中有7 種NRAMP 家族基因已被報道[13-17],其中OsNRAMP1負責轉運Cd 和Fe[18],OsNRAMP2參與Fe 從液泡到細胞質的轉運[19],楊猛[20]研究表明OsNRAMP3 主要參與Mn 的再利用,OsNRAMP5是Fe、Mn 和Cd 的轉運蛋白,OsNRAMP6 為Cd 轉運蛋白。
關于煙草NRAMP 家族成員的功能鮮見報道,陳邦蘭等[21]推測有10 個成員屬于NRAMP 家族,其中Tang 等[22]研究表明NtNRAMP5編碼的蛋白定位于質膜,有轉運Mn 和Cd 的能力。Jia 等[23]研究證實NtNRAMP3編碼的蛋白定位于液泡膜,有轉運Cd 的能力,但關于NRAMP 基因家族成員在煙草中對Fe 的功能尚未明晰。
本研究從煙草中克隆獲得一個NRAMP 家族基因,通過鑒定該基因與其他物種基因的同源性,命名為NtNRAMP3b,進一步通過RT-qPCR 和轉基因技術系統探究了NtNRAMP3b在鐵轉運中的功能,為明晰NtNRAMP3b在煙草轉運鐵中的機理提供理論基礎。
1 材料與方法
1.1 材料
普通煙草品種K326(Nicotiana tabacumcv,K326)作為供試材料,將煙草種子置于黑暗培養室(溫度28℃)中生長,培養3 d,然后將發芽的幼苗置于光照培養箱(光16 h 28℃/暗8 h 22℃,光強為300 μmol/(m2·s),相對濕度60%)中。采用霍格蘭營養液水培法,營養液pH 調至7.0[24]。在種子露白后使用1/4 正常營養液培養3 d,之后更換為1/2正常營養液培養3 d,最后更換為正常營養液培養4 d。培養結束后對K326 進行正常處理(CK:霍格蘭營養液)和缺鐵處理(-Fe:0 μmol/L FeSO4·7H2O與Na2-EDTA 螯合物),處理7 d 后取樣將收集的植物用去離子水洗滌,樣品的根、莖、幼葉和老葉(幼葉為完全展開的葉片從上往下數第2 片,老葉為完全展開的葉片從上往下數第7 片)用于測定基因的瞬時表達分析。
1.2 方法
1.2.1 目的基因的克隆 參考煙草在GenBank 收錄的煙草NtNRAMP3b基因組數據庫(GenBank 登錄號:XM_016614857.1)中已發表的序列數據,利用煙草K326 的基因特異性引物(F:5′-ATGCCTCCACACGATGACGA-3′;R:5′-TCAATTCTCTATGCTGGTGA-3′),以煙草cDNA 為模板進行PCR 擴增,擴增條件為94℃ 5 min;94℃ 30 s,58℃ 30 s,72℃2 min,35 個循環;72℃ 5 min。送Invitrogen 生物公司測序驗證,獲得全長cDNA 序列信息。
1.2.2 序列分析方法 利用NCBI 軟件的Open Reading Frame Viewer(ORF)(https://www.ncbi.nlm.nih.gov/orffinder/)進行基因序列的蛋白翻譯,利用Expasy(https://web.expasy.org/protparam/)和PRABI(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopm.html)分析NtNRAMP3b 的蛋白結構,利用在線網站NetPhos-3.1(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)預測NtNRAMP3b 的磷酸化位點,利用TMHMM-2.0(https://services.hea lthtech.dtu.dk/service.php?TMHMM-2.0)預測NtNRAMP3b 蛋白的跨膜結構域,運用DNAMAN 進行蛋白質多序列比對、分析同源性,利用MEGA6.0 制作鄰接樹(neighbor-joining tree,NJ),分析NtNRAMP3b與NRAMP 家族成員的親緣關系,在線網站MEME(https://meme-suite.org/meme/tools/meme)分析序列基序(motif),利用TBtools[25]將NJ-tree 和motif 組合,對NtNRAMP3b 與煙草、擬南芥中已經研究的NRAMP 家族進行序列分析。
1.2.3 啟動子分析 以煙草NtNRAMP3b全長序列為探針,利用Sol Genomics Network 網站進行BLAST搜索基因全長,在ATG 上游取一段長2 000 bp 的啟動子序列,利用PLANTCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在線工具分析啟動子序列的順式元件。
1.2.4 基因表達分析 運用Primer Premier6.0 軟件設計NtNRAMP3b基因擴增引物序列(F:5′-GATGACGAACAGCAACAAT-3′;R:5′-AATGACAACACCAGACCAA-3′),按照Invitrogen 公司的RealMaster-Mix(SYBR Green)試劑盒說明書進行RT-qPCR,試驗數據以煙草組成型表達基因NtL25(GenBank:L18908.1,引物序列為F:5′-CCGTCCAAAAAATCTGACCC-3′;R:5′-TCTTCAAAGTCTTAGGTCGG-3′)為內參基因,3 次重復試驗,采用2-ΔΔCT算法進行分析,ΔΔCT=Treat(樣品-L25)-CK(樣品-L25)[26]。
1.2.5 載體的構建及遺傳轉化 對測序正確的NtNRAMP3bcDNA 進行PCR 擴增得到基因全長序列,并設計引物(NtNRAMP3b-OE)與pCAMBIA1305 載體連接,該載體由花椰菜花葉病毒(CaMV)35S 啟動子和胭脂堿合成酶終止子驅動,轉化大腸桿菌DH5α 感受態細胞,經過菌液 PCR 檢測對比后,得到過表達載體 pCAMBIA1305-NtNRAMP3b。利用電轉化將其轉入農桿菌 EHA105 感受態細胞中,篩選陽性菌液侵染野生型煙草K326[27],室內繁種,收取轉基因植株T0種子,繁種獲得T2純合植株用于接下來的研究。
1.2.6 轉基因煙草株系對鐵的耐受性鑒定
1.2.6.1 觀察煙草表型并記錄生物量 將WT 和NtNRAMP3b-OE株系OE1、OE2 用上述水培法培養約35 d,用缺鐵營養液(-Fe:0 μmol/L)脅迫處理7 d,各材料取3 株長勢一致的幼苗,測定生物量。
1.2.6.2 葉綠素濃度和鐵濃度的測定 取正常處理和缺鐵處理7 d 的WT 和NtNRAMP3b-OE幼苗,利用無水乙醇提取法測定葉片中葉綠素濃度,利用鄰菲啰啉比色法測量各處理中不同材料的Fe 濃度。
1.2.6.3 相關基因瞬時表達的測定 鐵還原酶基因(ferric reduction oxidase,FRO)與鐵調節蛋白(iron-regulated transporter,IRT)已證實參與亞鐵離子的吸收[28],以NtL25為內參基因,利用RT-qPCR測定鐵脅迫處理后NtIRT1(AB263746.F:5′-TGGCTATTGACTCCATGGCT-3′;R:5′-AGTAGTTTCGCGCCATCAAC-3′)和NtFRO2(XM_016644133.F:5′-TATTGGGCTGCCACTCATCA-3′;R:5′-CCAGCATAAGAGACGCCAAC-3′)的相對表達量。
1.2.7 數據分析 使用DPS 7.5 進行單因素完全隨機試驗統計分析,運用LSD 法進行多重比較,分析1%極顯著水平并使用字母標記表示結果,利用Origin2018 和TBtools 作基因相對表達圖。
2 結果
2.1 目的基因克隆及序列分析
克隆獲得NtNRAMP3b,全長為1 530 bp,可編碼509 個氨基酸。編碼蛋白的分子式為C2621H4078N634O698S16,原子總數為8 047,相對分子質量為56.15 kD,理論等電點(pI)為5.77。帶負電荷的殘基的總數為42 個,帶正電荷的殘基有35 個,不穩定性指數為32.06,屬于穩定蛋白。
2.2 蛋白結構分析
NtNRAMP3b 的二級結構中,α 螺旋占比51.47%,β- 轉角占比9.04%,無規卷曲占比19.84%,延伸鏈占比19.65%;含有24 個絲氨酸、10 個蘇氨酸和3 個酪氨酸,推測在這些位點可能發生磷酸化。NtNRAMP3b 具有11 個疏水的跨膜區域(圖1)。
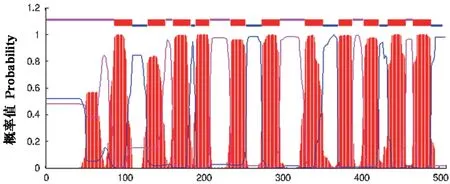
圖1 NtNRAMP3b 跨膜結構域分析Fig.1 Cross-membrane domain analysis of NtNRAMP3b
2.3 同源性與系統進化樹分析
NCBI 中查詢煙草、擬南芥與水稻的NRAMP 家族基因的蛋白質序列,運用DNAMAN 進行蛋白質多序列比對。結果(圖2)表明,NtNRAMP3b 與NtNRAMP2 的同源性為67.28%,與AtNRAMP3 的同源性為74.85%,與水稻OsNRAMP6 的同源性為61.86%,推測NtNRAMP3b 與AtNRAMP3 具有相似的功能。

圖2 同源性分析Fig.2 Homology analysis
運用MEGA6.0 制作鄰接樹,分析NtNRAMP3b與其余家族成員的親緣關系,并驗證NtNRAMP3b的潛在功能。在線網站MEME 分析序列基序(motif,圖3),NtNRAMP3b 與擬南芥AtNRAMP3、AtNRAMP4 的親緣關系較近,同時,NtNRAMP3b 與擬南芥AtNRAMP3 含有相似的motif 類型,表明他們可能具有相似的生物學功能。
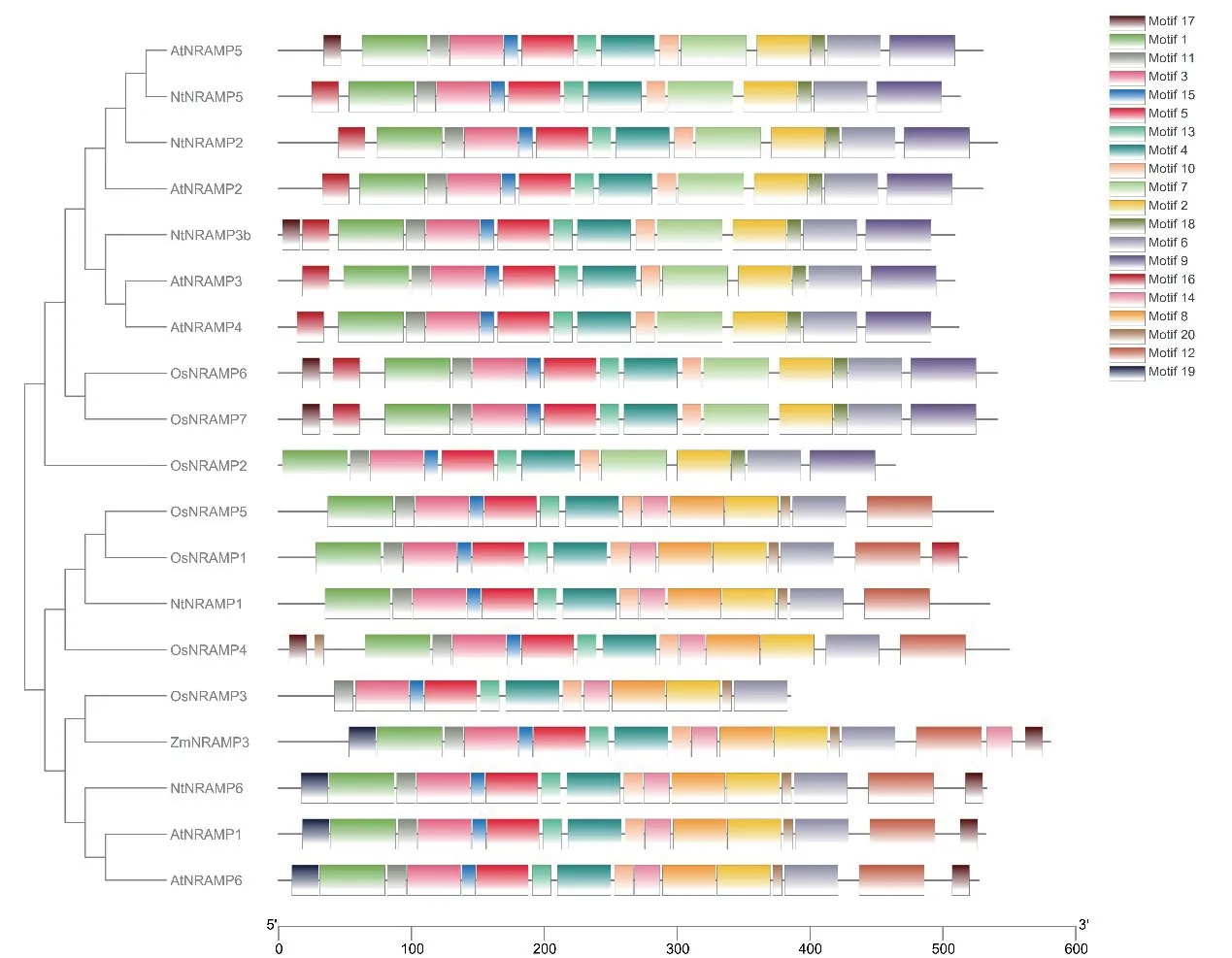
圖3 系統進化樹和motif 分析Fig.3 Phylogenetic tree and motif analysis
2.4 啟動子分析
如圖4所示,啟動子中除含有所必須的TATABOX 和CAAT-BOX 外,還含有植物激素響應元件(如1 個赤霉素反應元件(P-box),2 個脫落酸反應性涉及的順式元件(ABRE)和1 個與水楊酸反應順式作用元件(TCA-element))、組織特異性調控元件(如2 個與分生組織表達相關的順式作用元件(CAT-box))和防御與脅迫響應元件(如2 個低溫響應順式元件(LTR)和5 個厭氧誘導響應元件(ARE))。其中,植物激素響應元件(如ABRE)可以促進或抑制植物激素(如脫落酸)誘導基因的表達,從而表現出加速或減緩植株衰老過程;防御與脅迫響應元件(如LTR)在脅迫(如低溫)下誘導基因表達,從而表現出相關基因的瞬時表達增強。

圖4 啟動子順式元件分析Fig.4 Promoter cis-element analysis
2.5 基因表達模式分析
通過對NtNRAMP3b進行組織特異性表達分析,結果(圖5)顯示,NtNRAMP3b在葉片中表達量極高,其中,老葉是根部的8.07 倍,幼葉是根部的2.28 倍。通過對煙草植株進行缺鐵處理后發現,NtNRAMP3b在各部位的相對表達量增加,其中,老葉是正常處理的3.39 倍。因此,該基因主要在葉片中表達,尤其是老葉,且受Fe 誘導表達。
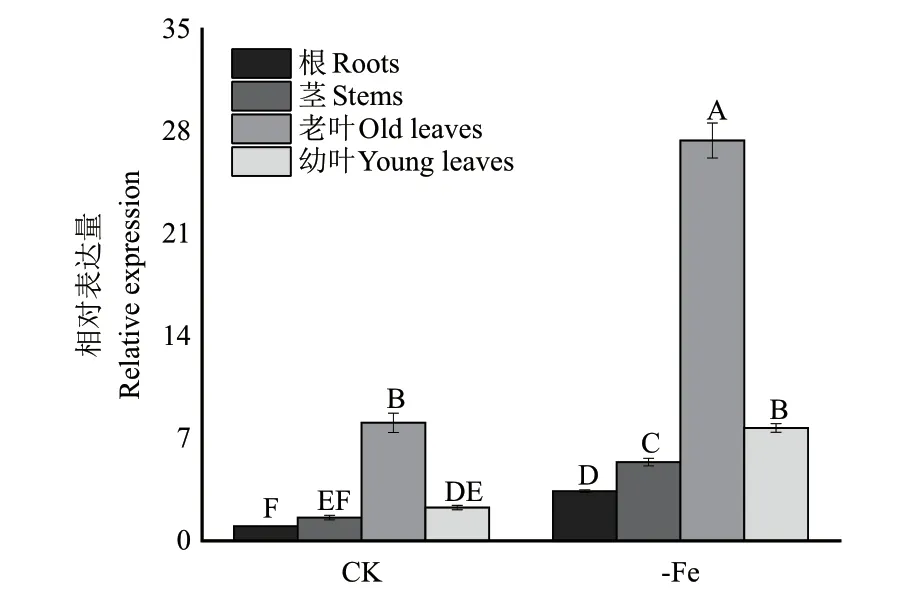
圖5 NtNRAMP3b 相對表達量Fig.5 NtNRAMP3b relative expression
2.6 缺鐵處理對NtNRAMP3b-OE煙草的影響
測定OE1 與OE2 植株NtNRAMP3b的瞬時表達,結果(圖6-A)顯示,OE1 中相對表達水平高。觀察煙株長勢,測量生物量。結果(圖6-B)顯示,正常處理下(CK)生長的煙株生物量不存在差異。在缺鐵處理后,煙株的整體生物量降低,相較之下,NtNRAMP3b-OE較WT 長勢較好,且OE1 的生物量顯著高于WT。葉綠素濃度結果(圖6-C)顯示,NtNRAMP3b-OE在缺鐵處理下葉綠素濃度極顯著的高于WT,表明NtNRAMP3b-OE煙株在缺鐵環境中促進葉綠素合成,從而影響煙株光合作用,提高了煙株的生物量積累。
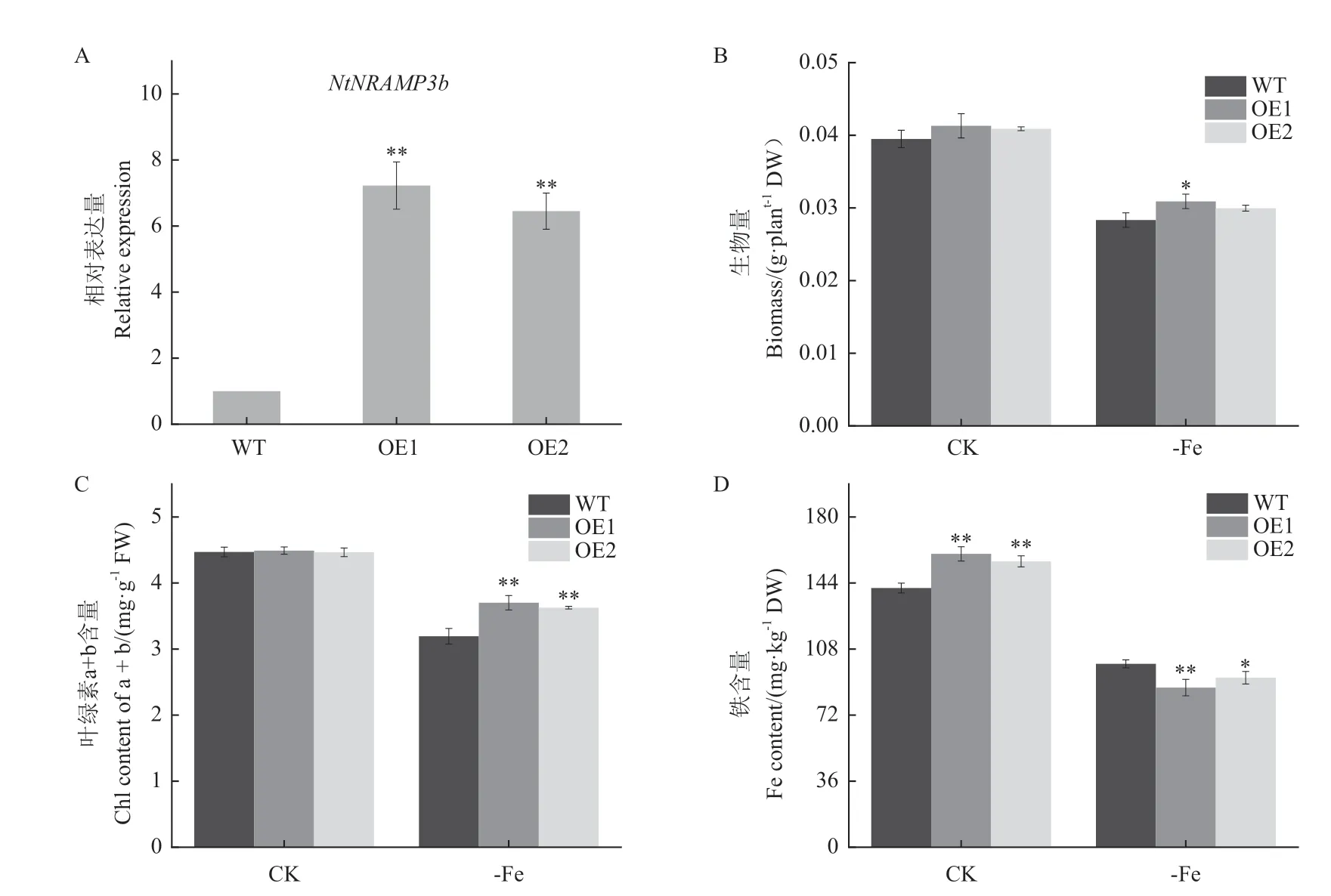
圖6 不同鐵濃度處理下的生物量Fig.6 Biomass under treatment with different cadmium concentrations
鐵濃度測定結果(圖6-D)顯示,正常處理下,NtNRAMP3b-OE植株鐵含量顯著高于WT,可能參與煙草對鐵的吸收。在缺鐵條件下,NtNRAMP3b-OE葉片中的鐵濃度低于WT,且OE1 與WT 間存在顯著差異,說明缺鐵處理后,NtNRAMP3b-OE植株的Fe2+被轉運至液泡外,從而提高葉綠素合成途徑相關酶活性和光敏素的合成,增加了葉綠素濃度和鐵的再利用,通過影響光合作用來影響煙株的生長發育,從而使煙株生物量和葉綠素濃度增加、鐵含量降低。
缺鐵條件下,OE 株系中的NtIRT1與NtFRO2的相對表達量極顯著高于WT(圖7),說明NtNRAMP3b-OE植株在缺鐵條件下通過增強NtIRT1與NtFRO2的表達,促進煙株對鐵的吸收與利用。
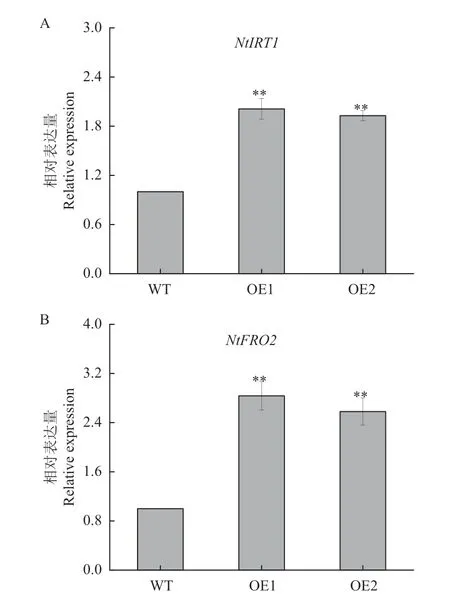
圖7 缺鐵條件下基因的瞬時表達Fig.7 Transient expressions of genes under iron deficiency
3 討論
本研究克隆獲得NtNRAMP3b,全長1 530 bp,編碼509 個氨基酸。其編碼蛋白等電點為5.77,含有51.47%的α 螺旋(alpha helix),不穩定性指數為32.06,屬于穩定蛋白,該基因編碼蛋白的理化性質與余沁等[29]研究結果一致。不同物種中的NRAMP 含有10-12 個跨膜螺旋結構[30],且高度保守。NtNRAMP3b 具有11 個疏水的跨膜區域,符合NRAMP 家族的跨膜結構特點。
Qin 等[31]研究表明NRAMP 可分為2 個亞家族,編碼蛋白分別定位于細胞膜和液泡膜,2 個亞家族成員的功能差異較大[32],其中,AtNRAMP2、AtNRAMP3、AtNRAMP4和AtNRAMP5定位于液泡膜上[8],主要參與液泡中金屬的釋放過程。NtNRAMP3b 的同源性與系統進化樹結果顯示,NtNRAMP3b 與At-NRAMP3 的親緣關系更近,Jia 等[23]研究表明NtNRAMP3 定位于液泡膜,屬于膜轉運蛋白,參與物質的轉運過程。NtNRAMP3b在缺鐵條件下上調表達,在葉片中的瞬時表達極顯著高于正常處理。在缺鐵處理下對鐵濃度分析發現,NtNRAMP3b-OE葉片的鐵含量低于WT,證實該基因在煙草中主要負責葉片中鐵從液泡向外的轉運過程。
Ihnatowicz 等[33]研究表明,NRAMP 家族在環境應激反應和光合作用等方面有調控作用[34]。啟動子中的ABRE 可以促進或抑制植物激素(如脫落酸)誘導基因的表達,從而表現出加速或減緩植株衰老過程。低溫響應順式元件(LTR)在低溫條件下誘導相關基因瞬時表達,從而改變植物的抗寒能力。低溫對類囊體產生氧化損傷[35-36],從而降低了光系統II(PSII)的功能。因此,可以通過RT-qPCR 進一步研究NtNRAMP3b在低溫下的表達情況,驗證NtNRAMP3b在逆境下的調控機制。
NRAMP 家族對Fe、Cd 和Mn 的轉運與吸收可以影響植物的生長發育,主要表現在缺Mn 誘導PSII 光抑制,產生活性氧(reactive oxygen species,ROS),植物膜脂過氧化程度高;過量鎘進入植物體內后影響植株生長發育,并通過食物鏈最終進入人體,危害人體健康[37-38];缺鐵(Fe)影響光系統I(PSI),導致葉綠素合成減少[39]。測定缺鐵環境下生長的WT 和NtNRAMP3b-OE的鐵濃度和葉綠素濃度發現,NtNRAMP3b-OE植株的葉綠素濃度顯著高于WT,而Fe 含量低于WT,說明NtNRAMP3b參與煙草葉片中Fe 的轉運,該基因過表達后,大量Fe 轉運至液泡外,參與PSI,使葉綠素含量增加,降低了煙株中的鐵含量。后期可通過不同Cd、Mn 條件處理煙株,觀察不同處理下煙株表型與長勢,測定葉綠素含量、Cd 濃度及ROS 含量來驗證NtNRAMP3b對光合作用的調控機理和對Cd、Mn 的轉運與吸收的分子機制。
4 結論
在煙草中克隆了天然抗性相關巨噬蛋白基因NtNRAMP3b,NtNRAMP3b主要在葉片中表達且受鐵脅迫誘導并增強表達。初步驗證了NtNRAMP3b主要參與鐵元素的轉運,影響葉片中鐵的吸收與再利用。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
當代化工研究(2016年9期)2016-03-20 16:22:08
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
聲屏世界(2014年6期)2014-02-28 15:18:09
終身教育研究(2014年5期)2014-02-28 01:23:06
中國煙草學報(2012年5期)2012-04-12 06:21:18
