植物SINA E3泛素連接酶功能的研究進(jìn)展
2023-01-05 08:45:26湯曉麗姜福東張洪霞
生物技術(shù)通報(bào) 2022年10期
湯曉麗 姜福東 張洪霞
(1.魯東大學(xué)農(nóng)林工程研究院,煙臺(tái) 264025;2.煙臺(tái)市農(nóng)業(yè)科學(xué)研究院,煙臺(tái) 264000)
泛素-蛋白酶體系統(tǒng)(ubiquitin-proteasome system,UPS)是真核生物細(xì)胞中重要且廣泛存在的蛋白平衡調(diào)節(jié)系統(tǒng),通過(guò)介導(dǎo)細(xì)胞中錯(cuò)誤折疊、受損傷、壽命較短的特別是具有調(diào)節(jié)作用的蛋白質(zhì)(轉(zhuǎn)錄因子和信號(hào)激酶)的泛素化降解,幾乎參與細(xì)胞中所有的生物學(xué)過(guò)程,是細(xì)胞中一種重要的翻譯后調(diào)節(jié)機(jī)制,更是細(xì)胞正常生命活動(dòng)秩序不可或缺的重要調(diào)節(jié)系統(tǒng)[1-4]。UPS由泛素激活酶E1、泛素結(jié)合酶E2、泛素連接酶E3、含76個(gè)氨基酸的泛素分子及26S蛋白酶體構(gòu)成[5]。其中,泛素連接酶E3的種類和數(shù)量復(fù)雜多樣決定UPS所降解靶蛋白的特異性[3]。
SINA(seven in absentia)E3泛素連接酶是一類RING(really interesting new gene)型E3泛素連接酶,蛋白的N端具有負(fù)責(zé)與E2酶結(jié)合的RING結(jié)構(gòu)域,C端是典型的SINA結(jié)構(gòu)域,負(fù)責(zé)蛋白的二聚體化以及與底物蛋白的結(jié)合[6-7]。最早關(guān)于SINA E3泛素連接酶的報(bào)道來(lái)自果蠅。在果蠅眼睛R7視光細(xì)胞的分化發(fā)育過(guò)程中,果蠅SINA發(fā)揮重要作用[8]。哺乳動(dòng)物小鼠的SINA在脊椎動(dòng)物的發(fā)育衍化過(guò)程中具有重要作用[9]。果蠅SINA編碼的E3泛素連接酶與轉(zhuǎn)錄抑制子Tramtrack(TTK)互作,介導(dǎo)TTK泛素化降解,影響果蠅眼睛R7細(xì)胞向視光細(xì)胞的分化[10]。同年,在人類中也發(fā)現(xiàn)了SINA的同源基因,隨著研究的不斷深入,越來(lái)越多的研究表明SINA編碼的E3泛素連接酶在生物體中的作用重要且廣泛[11-16]。然而,與動(dòng)物相比,SINA E3泛素連接酶在植物中的研究起步較晚(表1)。本研究對(duì)目前SINA在植物中的功能研究進(jìn)展進(jìn)行總結(jié)概述。
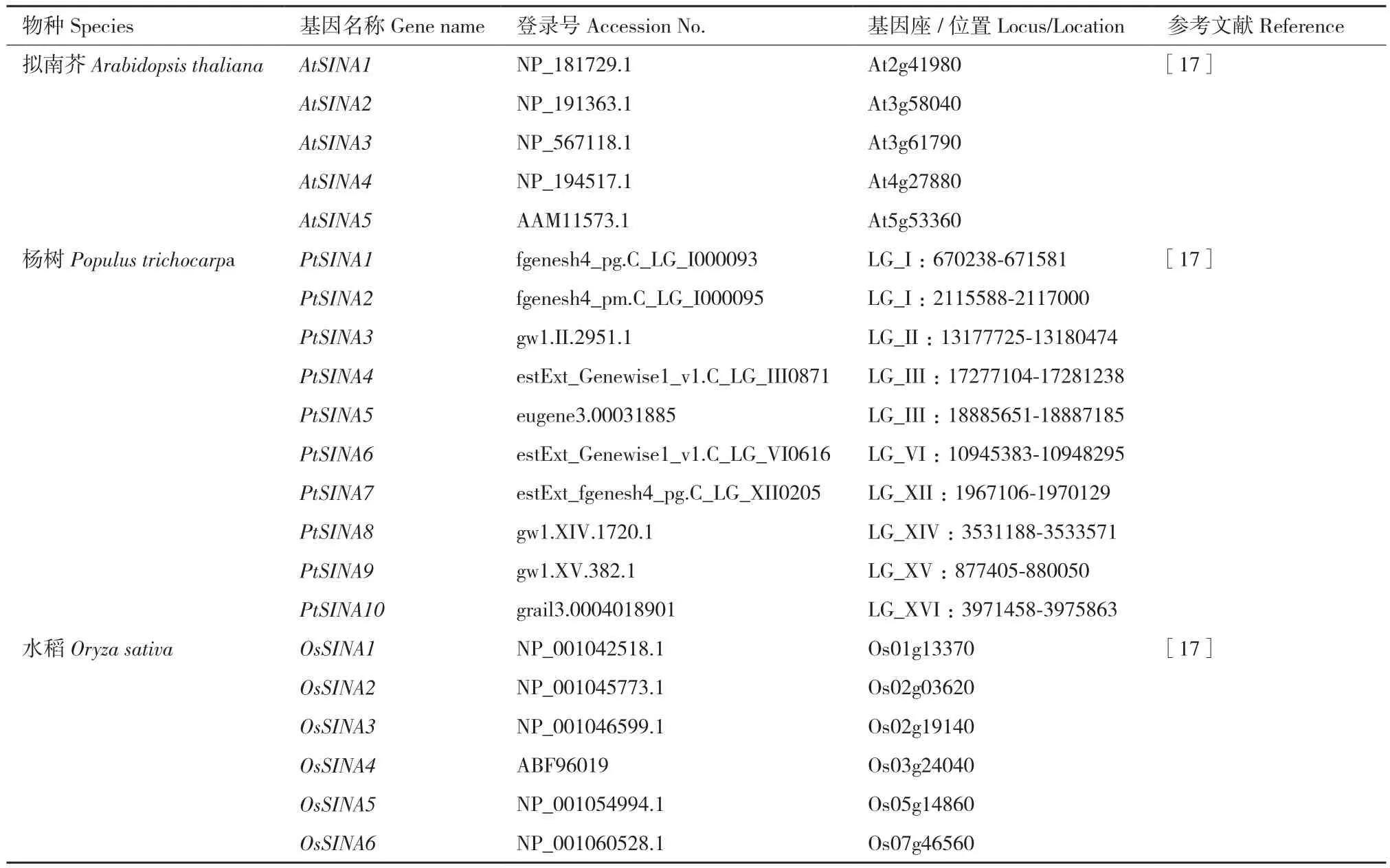
表1 目前研究報(bào)道的植物體中的SINATable 1 Reported SINA genes in plant at present
1 SINA E3泛素連接酶在植物生長(zhǎng)發(fā)育中的功能
擬南芥(Arabidopsis thaliana)SINAT5是植物體中第一個(gè)被發(fā)現(xiàn)的SINA E3泛素連接酶。2002年,Xie等[23]以AtNAC1為誘餌蛋白,尋找互作蛋白時(shí)發(fā)現(xiàn)了SINAT5。進(jìn)一步研究發(fā)現(xiàn),SINAT5與AtNAC1互作,并將其泛素化,進(jìn)而通過(guò)泛素介導(dǎo)的蛋白酶體途徑將其降解,終止植物體由AtNAC1傳導(dǎo)的生長(zhǎng)素信號(hào),實(shí)現(xiàn)了對(duì)植物側(cè)根發(fā)育的調(diào)控。雖然生長(zhǎng)素信號(hào)可以同時(shí)誘導(dǎo)SINAT5和AtNAC1(2個(gè)在生長(zhǎng)素信號(hào)傳導(dǎo)中具有拮抗作用的基因)的表達(dá),但是,在生長(zhǎng)素信號(hào)的傳導(dǎo)中,SINAT5對(duì)AtNAC1的翻譯后調(diào)節(jié)發(fā)揮了最終的決定性作用,調(diào)
節(jié)了植物的生長(zhǎng)發(fā)育。另外,置換SINAT5的RING結(jié)構(gòu)域酶活位點(diǎn)的關(guān)鍵氨基酸,使其喪失泛素連接酶活性,其對(duì)應(yīng)的超表達(dá)株系則表現(xiàn)出側(cè)根數(shù)量增加。SINAT5以二聚體形式發(fā)揮作用,雖然突變的SINAT5仍可以參與二聚體化,但不具有連接酶的活性,因此,突變的SINAT5競(jìng)爭(zhēng)性抑制了活性二聚體的產(chǎn)生,發(fā)揮了相反的生物學(xué)作用[23]。2008年,den Herder等[18]在苜蓿中進(jìn)行了SINAT5異源表達(dá),得到了相同的結(jié)果。超表達(dá)SINAT5轉(zhuǎn)基因苜蓿側(cè)根數(shù)量明顯減少,而SINAT5功能缺失突變體(SINAT5DN)的側(cè)根數(shù)量明顯增加[18]。超表達(dá)SINAT5DN轉(zhuǎn)基因植株的地上部長(zhǎng)勢(shì)和葉片大小明顯優(yōu)于野生型[18]。綜上所述,SINA E3泛素連接酶對(duì)植物正常生長(zhǎng)發(fā)育具有重要作用。
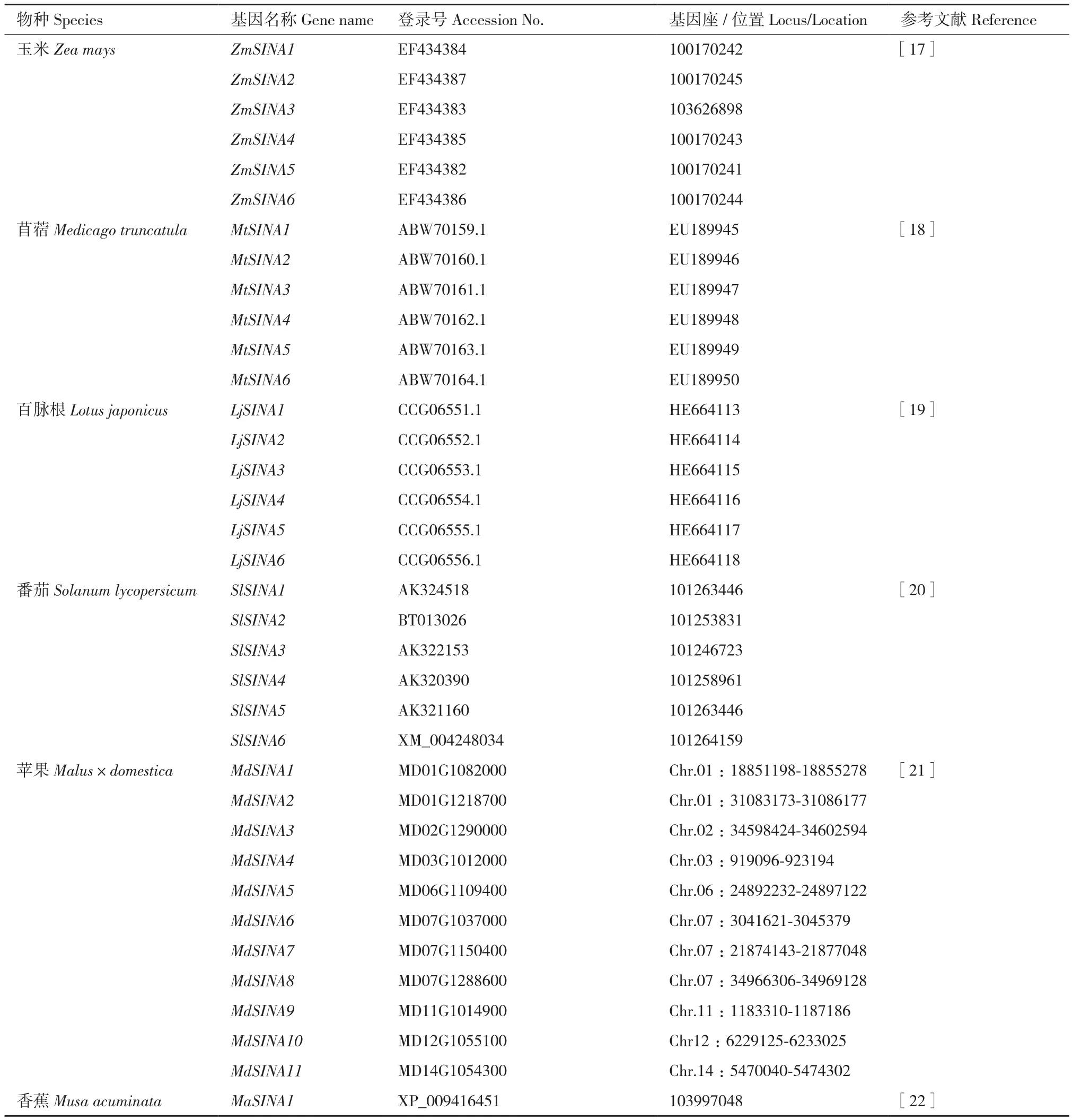
續(xù)表Continued
Park等[24]發(fā)現(xiàn)在擬南芥由營(yíng)養(yǎng)生長(zhǎng)向生殖生長(zhǎng)轉(zhuǎn)變的過(guò)程中,SINAT5參與調(diào)控了擬南芥的開(kāi)花。FLC(flowering locus C)是一個(gè)MADS-box轉(zhuǎn)錄因子,與SINAT5互作,通過(guò)抑制成花誘導(dǎo)過(guò)程中的一系列基因(如FT、SOC1等),進(jìn)而抑制植物的開(kāi)花過(guò)程[24-26]。LHY蛋白是擬南芥生物周期節(jié)律的一個(gè)關(guān)鍵組分,能夠根據(jù)外界光照和溫度的變化調(diào)節(jié)細(xì)胞的生物學(xué)活動(dòng)和植物體的生理適應(yīng)。SINAT5介導(dǎo)LHY(late elongated hypocotyl)蛋白的泛素化降解,進(jìn)而調(diào)節(jié)擬南芥的開(kāi)花時(shí)間[27-29]。研究還發(fā)現(xiàn),光信號(hào)的傳導(dǎo)蛋白DET1(DE-ETIOLATED1)可以抑制SINAT5對(duì)LHY的泛素化降解。因此,SINAT5通過(guò)對(duì)靶蛋白的泛素化降解建立了光周期與植物開(kāi)花時(shí)間的聯(lián)系,對(duì)植物的生長(zhǎng)發(fā)育特別是由營(yíng)養(yǎng)生長(zhǎng)向生殖生長(zhǎng)的轉(zhuǎn)變發(fā)揮重要作用。
擬南芥SINA E3泛素連接酶基因家族有5個(gè)成員,SINAT2是擬南芥中第二個(gè)被發(fā)現(xiàn)的SINA E3泛素連接酶基因,也參與了擬南芥正常生長(zhǎng)發(fā)育的調(diào)節(jié)過(guò)程。Welsch等[30]研究發(fā)現(xiàn),SINAT2通過(guò)與RAP2.2(乙烯響應(yīng)元件結(jié)合蛋白轉(zhuǎn)錄因子)互作,影響擬南芥類胡蘿卜素的合成。油菜素內(nèi)酯(BRs)是一類新型的廣譜植物生長(zhǎng)調(diào)節(jié)劑,參與調(diào)節(jié)植物的營(yíng)養(yǎng)生長(zhǎng)、生殖生長(zhǎng)、種子發(fā)芽以及衰老等一系列的生長(zhǎng)發(fā)育過(guò)程[31]。BRI1-EMS-SUPPRE SSOR 1(BES1)是BR信號(hào)通路中一個(gè)重要的轉(zhuǎn)錄因子,調(diào)節(jié)BR信號(hào)下游基因的表達(dá)[32]。擬南芥5個(gè)SINA E3均能與BES1互作,促進(jìn)光下BES1的降解,抑制其下游基因的表達(dá),從而光下植株下胚軸的伸長(zhǎng)生長(zhǎng)被抑制[33]。相反,在黑暗條件下,SINAs發(fā)生自泛素化降解,BES1蛋白正常發(fā)揮生物學(xué)功能,促進(jìn)植株下胚軸的伸長(zhǎng)生長(zhǎng)。同時(shí),該研究還發(fā)現(xiàn),SINAs光下積累量增加,反之減少,BES1含量也隨之發(fā)生變化[33]。因此,表明SINA E3通過(guò)介導(dǎo)BES1的降解,連通了光信號(hào)和BR信號(hào),調(diào)節(jié)了植物的光形態(tài)建成。Wang等[20]研究發(fā)現(xiàn)超表達(dá)SlSINA2轉(zhuǎn)基因番茄的葉片顏色變淺,葉綠素含量降低,植株生長(zhǎng)發(fā)育遲緩,嚴(yán)重影響了番茄正常的生長(zhǎng)發(fā)育,還發(fā)現(xiàn)超表達(dá)SlSINA5轉(zhuǎn)基因番茄的花發(fā)育出現(xiàn)異常,花萼、花瓣、雄蕊以及整朵花均不同程度地變小,但具體的作用機(jī)制還有待進(jìn)一步探究。因此,不同的SINA E3通過(guò)靶向介導(dǎo)各種各樣靶蛋白的降解,調(diào)節(jié)植物復(fù)雜多樣的生長(zhǎng)發(fā)育過(guò)程。
2 SINA E3泛素連接酶在植物響應(yīng)逆境脅迫中的功能
隨著研究的不斷深入,越來(lái)越多的研究發(fā)現(xiàn),SINA E3除了調(diào)節(jié)植物生長(zhǎng)發(fā)育外,還參與了植物對(duì)多種逆境脅迫的響應(yīng)。植物中第一個(gè)被發(fā)現(xiàn)參與逆境脅迫響應(yīng)的SINA E3是水稻(Oryza sativa)的OsDIS1(Oryza sativa drought-induced SINA protein 1)。2011年Ning等[34]篩選獲得受干旱誘導(dǎo)表達(dá)的OsDIS1,OsDIS1抑制了水稻的抗旱性。隨后,Ning等[35]進(jìn)一步研究發(fā)現(xiàn)OsDIS1通過(guò)與OsSKIPa(干旱和鹽脅迫的正調(diào)因子)互作,影響了下游脅迫應(yīng)答響應(yīng)基因的表達(dá),進(jìn)而調(diào)節(jié)水稻的抗旱性。
與水稻SINA E3功能恰恰相反,AtSINA2提高了擬南芥的干旱脅迫抗性,超表達(dá)AtSINA2后,轉(zhuǎn)基因擬南芥的抗旱性明顯增強(qiáng)。進(jìn)一步研究發(fā)現(xiàn),干旱條件下AtSINA2促進(jìn)ABA介導(dǎo)的植物葉片氣孔的閉合,降低水分散失,進(jìn)而提高了轉(zhuǎn)基因植株的抗旱性[36]。與擬南芥 5 個(gè) SINA E3(SINAT1-SINAT5)相比,AtSINA2(也被稱為SINAT6)的N端含有一個(gè)不完整的RING結(jié)構(gòu)域,認(rèn)為與SINAT5DN一樣,AtSINA2沒(méi)有E3泛素連接酶的活性,競(jìng)爭(zhēng)性抑制了活性二聚體的產(chǎn)生,因此,發(fā)揮了與OsDIS1截然相反的作用[36]。然而,2018年Chen等[37]研究發(fā)現(xiàn),AtSINA2蛋白N端的B-box結(jié)構(gòu)域彌補(bǔ)了不完整的RING結(jié)構(gòu)域,使其仍具有E3泛素連接酶的活性。同時(shí),該研究還發(fā)現(xiàn)AtSINA2可以與CDKG1(細(xì)胞周期蛋白依賴的蛋白激酶G1)互作,而CDKG1對(duì)AtSINA2的磷酸化直接影響了AtSINA2的穩(wěn)定性,進(jìn)而調(diào)節(jié)了植物體對(duì)ABA和滲透脅迫的響應(yīng)[37],表明SINA E3調(diào)節(jié)其靶蛋白穩(wěn)定性的同時(shí),其自身的活性功能同樣受蛋白的磷酸化和去磷酸化調(diào)節(jié),因此,SINA E3發(fā)揮功能的機(jī)制復(fù)雜而多樣。ICE(INDUCER OF CBF EXPRESSION)是一類MYC型bHLH轉(zhuǎn)錄因子,目前,ICE-CBF-COR轉(zhuǎn)錄級(jí)聯(lián)途徑是已知植物體中重要的冷脅迫響應(yīng)信號(hào)途徑[38]。2017年,F(xiàn)an等[22]研究發(fā)現(xiàn)香蕉SINAE3還參與了植物對(duì)冷脅迫的響應(yīng),發(fā)現(xiàn)MaSINA1通過(guò)泛素化降解轉(zhuǎn)錄因子MaICE1,進(jìn)而抑制MaICE1下游冷脅迫重要調(diào)節(jié)因子MaNAC1的表達(dá),調(diào)節(jié)植物對(duì)冷脅迫的應(yīng)答響應(yīng)。
ABA是所有植物激素中最重要也是作用最廣泛的一種激素,其處于植物對(duì)非生物脅迫應(yīng)答調(diào)節(jié)的核心,被稱為“植物逆境激素”。AtSINA2通過(guò)調(diào)節(jié)由ABA介導(dǎo)的氣孔閉合而提高植物的抗旱性[36]。2020 年 Li等[21]發(fā)現(xiàn)蘋(píng)果 MdSINA2 可以被ABA誘導(dǎo)表達(dá),超表達(dá)MdSINA2轉(zhuǎn)基因蘋(píng)果愈傷和擬南芥均表現(xiàn)出ABA敏感,說(shuō)明與AtSINA2一樣,MdSINA2參與了ABA信號(hào)調(diào)節(jié)植物對(duì)干旱等多種逆境脅迫的應(yīng)答響應(yīng)[21]。同時(shí),蘋(píng)果SINA E3還調(diào)節(jié)植物對(duì)營(yíng)養(yǎng)元素匱乏的響應(yīng)。2020年,Xia等[39]發(fā)現(xiàn)超表達(dá)SINAT1、SINAT2、SINAT3和SINAT4的轉(zhuǎn)基因植株對(duì)ABA的敏感性增加,SINATs通過(guò)泛素化降解FREE1和VPS23A(蛋白運(yùn)輸?shù)陌麅?nèi)分揀復(fù)合體的關(guān)鍵組分),解除它們對(duì)ABA受體PYR和PYL4的抑制作用,進(jìn)而參與植物的ABA信號(hào)傳導(dǎo)。當(dāng)植物體中ABA含量升高,SINATs受誘導(dǎo)含量升高,與FREE1和VPS23A發(fā)生互作,將其降解,此時(shí)ABA的受體被釋放發(fā)揮作用,啟動(dòng)ABA的傳導(dǎo)及響應(yīng)基因的表達(dá),植物體表現(xiàn)出對(duì)ABA的應(yīng)答響應(yīng)。
3 SINA E3泛素連接酶在植物與其他生物互作中的功能
植物固著生長(zhǎng)的特性使植物被動(dòng)遭受外界環(huán)境的各種有利因素和不利因素的影響,SINA E3不僅調(diào)節(jié)植物在適宜條件下的生長(zhǎng)發(fā)育,同時(shí)也參與植物對(duì)各種不利環(huán)境條件的應(yīng)答。當(dāng)外界環(huán)境營(yíng)養(yǎng)條件匱乏時(shí),植物一方面通過(guò)改變根系的構(gòu)型,盡可能多地從外界土壤中吸收養(yǎng)分;另一方面,植物利用根系與土壤中各類生物建立共生關(guān)系來(lái)吸收養(yǎng)分[40]。在根系的構(gòu)型方面,SINAT5通過(guò)影響根毛的多少來(lái)發(fā)揮作用。SINA E3在植物根系與土壤中各類生物相互作用中也發(fā)揮著重要作用。
den Herder等[18]發(fā) 現(xiàn) 異 源 表 達(dá) SINAT5和SINAT5DN的苜蓿,不僅影響了它的正常生長(zhǎng)發(fā)育,同時(shí)也影響了根毛與根瘤菌之間的相互作用,抑制了根瘤菌與根毛形成穩(wěn)定的共生關(guān)系和共生結(jié)構(gòu)。共生類受體蛋白激酶(symbiosis receptor-like kinase,SYMRK)是微生物信號(hào)分子刺激植物根細(xì)胞后,植物體進(jìn)行共生信號(hào)傳導(dǎo)的關(guān)鍵組分[41]。2012年,den Herder等[19]發(fā)現(xiàn)百脈根中的LjSINA4 E3可以與SYMRK互作,并且超表達(dá)LjSINA4后,SYMRK蛋白含量降低,抑制了與微生物進(jìn)行互利共生信號(hào)的傳導(dǎo),進(jìn)而抑制了百脈根與根瘤菌的相互作用。
SINA E3除了參與植物與共生微生物之間的相互作用外,還參與了植物與病原微生物之間的相互作用。2016年Miao等[42]發(fā)現(xiàn)番茄SlSINA3可以將轉(zhuǎn)錄因子SlNAC1泛素化降解。SlNAC1是番茄脅迫相關(guān)NAC轉(zhuǎn)錄因子,可以被病原微生物假單胞菌(Pseudomonas)誘導(dǎo)表達(dá),是植物防御病原微生物侵染的有效策略。當(dāng)煙草SlNAC1同源基因被抑制表達(dá)后,煙草對(duì)假單胞菌的敏感性明顯增強(qiáng)[43]。超表達(dá)SlSINA3后,抑制了阻力蛋白R(shí)(resistance)蛋白誘導(dǎo)的超敏反應(yīng)(hypersensitive response,HR)。因此,SlSINA3通過(guò)翻譯后水平的調(diào)節(jié)作用,抑制了番茄對(duì)病原微生物的防御反應(yīng)。除了SlSINA3以外,番茄其他SINA E3也參與了番茄對(duì)病原微生物的防御響應(yīng)。其中,SlSINA4可以激活細(xì)胞中防御相關(guān)信號(hào)的傳導(dǎo),促進(jìn)植物體對(duì)病原微生物侵染的應(yīng)答響應(yīng)。與SlSINA4不同,SlSINA1、SlSINA2、SlSINA5和SlSINA6則是通過(guò)降解R蛋白下游的相關(guān)組分,發(fā)揮與SlSINA3相同的功能,抑制植物對(duì)病原微生物的防御響應(yīng)[43]。因此,面對(duì)外界病原微生物的入侵,植物體中不同SINA E3發(fā)揮著各自不同的作用,有的相互協(xié)同發(fā)揮作用,有的相互拮抗發(fā)揮作用,共同調(diào)節(jié)植物體對(duì)病原微生物的響應(yīng)。
4 SINA E3泛素連接酶在植物細(xì)胞自噬中的功能
與UPS對(duì)細(xì)胞中蛋白平衡調(diào)節(jié)作用一樣,細(xì)胞自噬(autophagy)是細(xì)胞中另一個(gè)重要的蛋白平衡調(diào)節(jié)系統(tǒng)。其中,UPS是對(duì)壽命較短、錯(cuò)誤折疊以及受損傷蛋白的主要降解途徑,細(xì)胞自噬主要負(fù)責(zé)清除大的胞質(zhì)組分,包括聚集的蛋白、喪失功能的細(xì)胞器和侵入的病原菌[39]。兩個(gè)蛋白平衡調(diào)節(jié)系統(tǒng)分別發(fā)揮不同的作用,又保持著千絲萬(wàn)縷的聯(lián)系。如前所述,SINA E3通過(guò)泛素化調(diào)節(jié)植物BR信號(hào)途徑BES1轉(zhuǎn)錄因子的穩(wěn)定性,進(jìn)而參與植物的光形態(tài)建成[33]。同時(shí)還發(fā)現(xiàn)SINAT2可以與泛素結(jié)合受體蛋白DOMINANT SUPPRESSOR OF KAR 2(DSK2)直接互作,參與BES1-DSK2-ATG8復(fù)合體的構(gòu)成,介導(dǎo)BES1經(jīng)細(xì)胞自噬途徑的降解,表明植物在逆境脅迫和N或C元素缺乏的條件下,SINA E3可以通過(guò)同時(shí)激活泛素蛋白酶途徑和細(xì)胞自噬途徑對(duì)BES1的降解[33]。因此,SINA E3在細(xì)胞自噬過(guò)程中具有抑制植物在逆境下生長(zhǎng)發(fā)育,提高植物抗逆性的作用,維持了植物在逆境條件下生長(zhǎng)和抗性能力的平衡。
2017年,Qi等[44]研究發(fā)現(xiàn)SINAT1、SINAT2和SINA2均可直接與細(xì)胞自噬相關(guān)蛋白ATGs發(fā)生互作,參與細(xì)胞自噬過(guò)程的調(diào)節(jié)。SINAT1/SINAT2和SINA2在調(diào)節(jié)細(xì)胞自噬方面發(fā)揮著相反作用,當(dāng)植物體的營(yíng)養(yǎng)條件良好時(shí),SINAT1和SINAT2促進(jìn)ATG6蛋白的泛素化降解,抑制細(xì)胞自噬;當(dāng)營(yíng)養(yǎng)條件匱乏時(shí),SINA2被誘導(dǎo)表達(dá),發(fā)揮相反作用,抑制ATG6的泛素化降解,促進(jìn)細(xì)胞自噬[44]。同時(shí)研究還指出,SINAT1和SINAT2在調(diào)節(jié)ATG6方面具有一定程度的功能冗余。因此該研究表明,植物體中SINA E3對(duì)細(xì)胞生命活動(dòng)的調(diào)節(jié)精準(zhǔn)而多樣,不僅可以通過(guò)UPS途徑直接發(fā)揮作用,同時(shí)也可以通過(guò)間接影響與UPS平行的其他調(diào)節(jié)途徑來(lái)發(fā)揮作用,實(shí)現(xiàn)了功能的進(jìn)一步強(qiáng)化。而對(duì)于每一個(gè)SINA E3及其家族成員具體的生物學(xué)功能還需要更深入細(xì)致的研究和探索。2020年,Qi等[45]再次發(fā)現(xiàn)SINAT1、SINAT2和SINA2還可以與自噬相關(guān)蛋白ATG13互作。ATG13通過(guò)與ATG1互作形成蛋白激酶復(fù)合體調(diào)節(jié)自噬小體的形成,進(jìn)而調(diào)節(jié)植物細(xì)胞自噬的過(guò)程[46]。植物體在不同的營(yíng)養(yǎng)條件下,就是通過(guò)SINAT1、SINAT2和SINA2調(diào)節(jié)ATG1和ATG13復(fù)合體的穩(wěn)定性,調(diào)節(jié)了細(xì)胞自噬過(guò)程,進(jìn)而決定植物在不同營(yíng)養(yǎng)條件下的應(yīng)答響應(yīng)。綜上所述,ATGs是細(xì)胞自噬的關(guān)鍵組分,在細(xì)胞自噬中發(fā)揮重要作用。SINA E3通過(guò)調(diào)節(jié)ATGs的穩(wěn)定性,間接參與了細(xì)胞自噬[47-49]。因此,UPS和細(xì)胞自噬是細(xì)胞中兩個(gè)非常重要的蛋白平衡調(diào)節(jié)系統(tǒng),兩者彼此獨(dú)立同時(shí)又相互依存,SINA E3的存在介導(dǎo)了兩個(gè)系統(tǒng)之間千絲萬(wàn)縷的聯(lián)系,UPS通過(guò)SINA E3實(shí)現(xiàn)了對(duì)細(xì)胞自噬的調(diào)節(jié)和影響,同時(shí)細(xì)胞自噬也通過(guò)SINA E3實(shí)現(xiàn)了自身功能的完善和強(qiáng)大。
5 總結(jié)與展望
SINA E3是一類重要的E3泛素連接酶,通過(guò)調(diào)節(jié)細(xì)胞中不同信號(hào)通路上游的轉(zhuǎn)錄因子或蛋白激酶等調(diào)節(jié)蛋白發(fā)揮生物學(xué)功能,作用廣泛而深入。他的調(diào)節(jié)作用賦予了植物面對(duì)外界復(fù)雜環(huán)境條件“文武之道,一張一弛”的智慧。條件適宜時(shí),植物正常生長(zhǎng)發(fā)育;環(huán)境惡劣時(shí),通過(guò)犧牲生長(zhǎng)速度換取繼續(xù)活下去的能力;當(dāng)有益微生物來(lái)臨時(shí),調(diào)節(jié)微生物與植物的和諧相處進(jìn)行互利共生;當(dāng)有害微生物侵入時(shí),將其限定在一定范圍內(nèi),防止其對(duì)植物進(jìn)一步侵害。當(dāng)然,植物中SINA E3的功能研究起步較晚,還有很多問(wèn)題有待未來(lái)發(fā)現(xiàn)和探索。包括:(1)AtSINA2的E3泛素連接酶活性受其蛋白磷酸化狀態(tài)的影響,其他的SINA E3活性是否也同樣存在蛋白的翻譯后調(diào)節(jié)?(2)植物中SINA E3以同源或異源二聚體發(fā)揮作用的具體機(jī)制是什么?(3)動(dòng)物中研究發(fā)現(xiàn)有的SINA E3可以以多聚體的形式發(fā)揮作用,植物中是否也存在相同的作用機(jī)制?(4)是否有更好的挖掘SINA E3互作蛋白的研究方法,進(jìn)而更高效的研究SINA E3在植物體中重要的生物學(xué)功能。(5)為什么植物中大多數(shù)目前研究已知的SINA E3均發(fā)揮負(fù)調(diào)作用?不論是SINA E3蛋白自身的磷酸化,還是同源和異源二聚體的形成機(jī)制以及SINA E3通過(guò)蛋白與蛋白的互作靶向降解不同的靶蛋白(調(diào)節(jié)蛋白或者功能蛋白),均涉及到蛋白與蛋白的互作,因此在未來(lái)SINA E3功能的研究中需綜合利用多種蛋白互作的研究技術(shù)解決目前SINA E3功能研究中遇到的諸多問(wèn)題,進(jìn)而對(duì)植物中SINA E3功能進(jìn)行進(jìn)一步的挖掘和探索。SINA E3多樣化的功能也讓我們初步領(lǐng)略了植物體龐大復(fù)雜的調(diào)控網(wǎng)絡(luò),為未來(lái)探索植物體中更多、更復(fù)雜的調(diào)節(jié)機(jī)制和功能途徑奠定基礎(chǔ)。
猜你喜歡
中華詩(shī)詞(2022年6期)2022-12-31 06:41:24
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
電子制作(2018年11期)2018-08-04 03:25:42
中國(guó)科技論壇(2017年7期)2017-07-25 08:49:53
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
