植物中C-糖基轉移酶的結構與應用
2023-01-05 08:45:26趙玉雪王蕓余璐瑤劉京晶斯金平張新鳳張磊
生物技術通報 2022年10期
關鍵詞:植物
趙玉雪 王蕓 余璐瑤 劉京晶 斯金平 張新鳳 張磊,,
(1.浙江農林大學省部共建亞熱帶森林培育國家重點實驗室,杭州 311300;2.上海大學醫學院生物醫藥創新研發中心,上海 200444;3.海軍軍醫大學藥學院藥用植物學教研室,上海 200433)
糖基化是植物次生代謝中較為普遍的修飾反應[1],催化該反應的后修飾酶是糖基轉移酶(glycosyltransferases,GT;EC 2.4.x.y),幾乎存在于所有植物中[2],植物糖基轉移酶參與不同生命過程,包括特化代謝、植物激素修飾、外源性生物解毒以及植物與病原體相互作用等[1,3-7]。例如蘋果O-糖基轉移酶參與的根皮苷合成是高品質蘋果培育的重要調控靶點[8];茶葉中的糖基轉移酶廣泛參與多酚類物質的合成,直接影響茶飲的口感[9];茶樹中的橙花叔醇糖基化參與調控冷脅迫響應[10]。
糖基化反應機制一直是植物代謝研究中的重要領域,在植物的糖基化反應中,常見的糖基供體為UDP-葡萄糖、UDP-半乳糖、UDP-木糖和UDP-鼠李糖等[11-15];而糖基受體比較多樣,不僅包括脂類、氨基酸、多糖類初生代謝產物,也包括木脂素、萜類、生物堿、黃酮類等次生代謝產物。糖基轉移酶能夠將糖基從活化的供體分子轉移到受體分子上,形成糖苷鍵,從而直接參與單糖苷、二糖、低聚糖、聚糖苷和多糖的生物合成[16-17],提高了植物天然產物的復雜性和多樣性[18-19]。
根據催化受體分子的糖基化位點的差異,糖苷產物分為O-型糖苷、C-型糖苷、N-型糖苷和S-型糖苷,對應的催化酶分為O-、C-、N-、S-四種糖基轉移酶。O-糖基轉移酶(O-glucosyltransferases,OGTs)的研究最為廣泛、深入,普遍參與植物次生代謝產物和激素的糖基化修飾,催化底物呈現出較為明顯的特征[20]。其催化產物對植物生長發育環境抗逆性等方面具有重要意義[21-22]。N-糖基化和S-糖基化是蛋白翻譯后修飾的重要方法,但植物中N-糖基轉移酶(N-glucosyltransferases,NGTs)和S-糖基轉移酶(S-glucosyltransferases,SGTs)的相關研究較少。番茄中O-糖基轉移酶對其莖尖的長度有著調控作用[23],穿心蓮中二萜糖基轉移酶ApUGT參與活性成分穿心蓮內酯的合成[24];模式植物擬南芥中發現的N-糖基轉移酶UGT76C1、UGT76C2可以催化玉米素、激動素、6-芐基氨基嘌呤等細胞分裂素[25],UGT76C4和 UGT76C5可以催化煙酸 N-糖基化[26];銀杏中的N-糖基轉移酶GbNGT1完善了生長素的代謝通道[27]。通過探究UGT74B1參與擬南芥硫代葡萄糖苷的生物合成,以及突變體的表型觀察,初步探索了S-糖基轉移酶的生物學功能[28]。
隨著糖基轉移酶研究的深入以及結構生物學的發展,C-糖基轉移酶(C-glucosyltransferases,CGTs)開始進入研究者的視野,有些C-糖苷類化合物具有抗氧化、抗糖尿病等多種生物活性和較高的藥用價值[29],如葛根素(大豆苷元8-葡萄糖苷)廣泛用于治療心血管疾病,芹菜苷元糖基化后能表現出明顯的抗病毒、抗糖尿病、抗阿爾茨海默病及抗炎等作用。
C-糖基轉移酶是天然和非天然產物C-糖基化的有力工具,但糖苷類化合物的化學合成困難,來源、結構多樣性及藥物研究受到一定程度的制約[30-34]。C-糖基化反應中,糖基供體的含量、C-糖基轉移酶的活性、蛋白表達量、底物的濃度等與碳苷化合物的產量密切相關[29]。目前對植物體內的碳苷類化合物的生物合成途徑和CGT的調控機制還有待深入研究。為了發現植物中的CGT,現階段最常用的方法是構建系統進化樹,將已表征的這些CGT以最大似然法構建系統進化樹(圖1),發現它們是一個同源性極高的基因家族,可以通過同源比對的方法對CGT進行挖掘和篩選。對糖基轉移酶及其基因家族的研究不僅有助于解析糖苷類化合物生物合成與調控的機制,還可以為研究合成有生物活性的糖苷類化合物提供新的路徑。本文綜述了植物中C-糖基轉移酶分類、空間結構與生物技術應用的研究進展,以期此類酶為今后的研究提供可借鑒的思路。
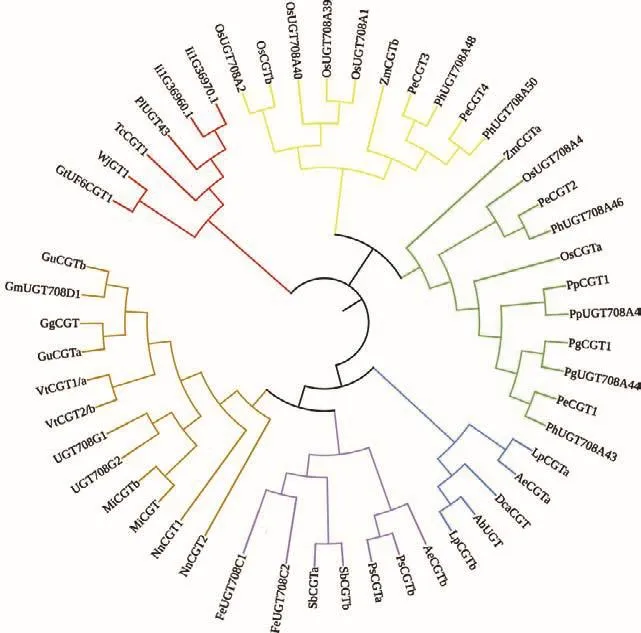
圖1 已有功能驗證C-糖基轉移酶的系統進化樹Fig.1 Phylogenetic tree of functionally validated C-glycosyltransferases
1 植物C-糖基轉移酶
C-糖基轉移酶催化糖基供體和受體形成C-C鍵,合成C-糖苷類化合物,此過程具有高效性和區域特異性。目前已報道的C-糖基轉移酶介導糖基化的底物主要以黃酮類化合物為主,也有部分苯丙素類和酚類化合物(圖2,相關CGTs見表1)。
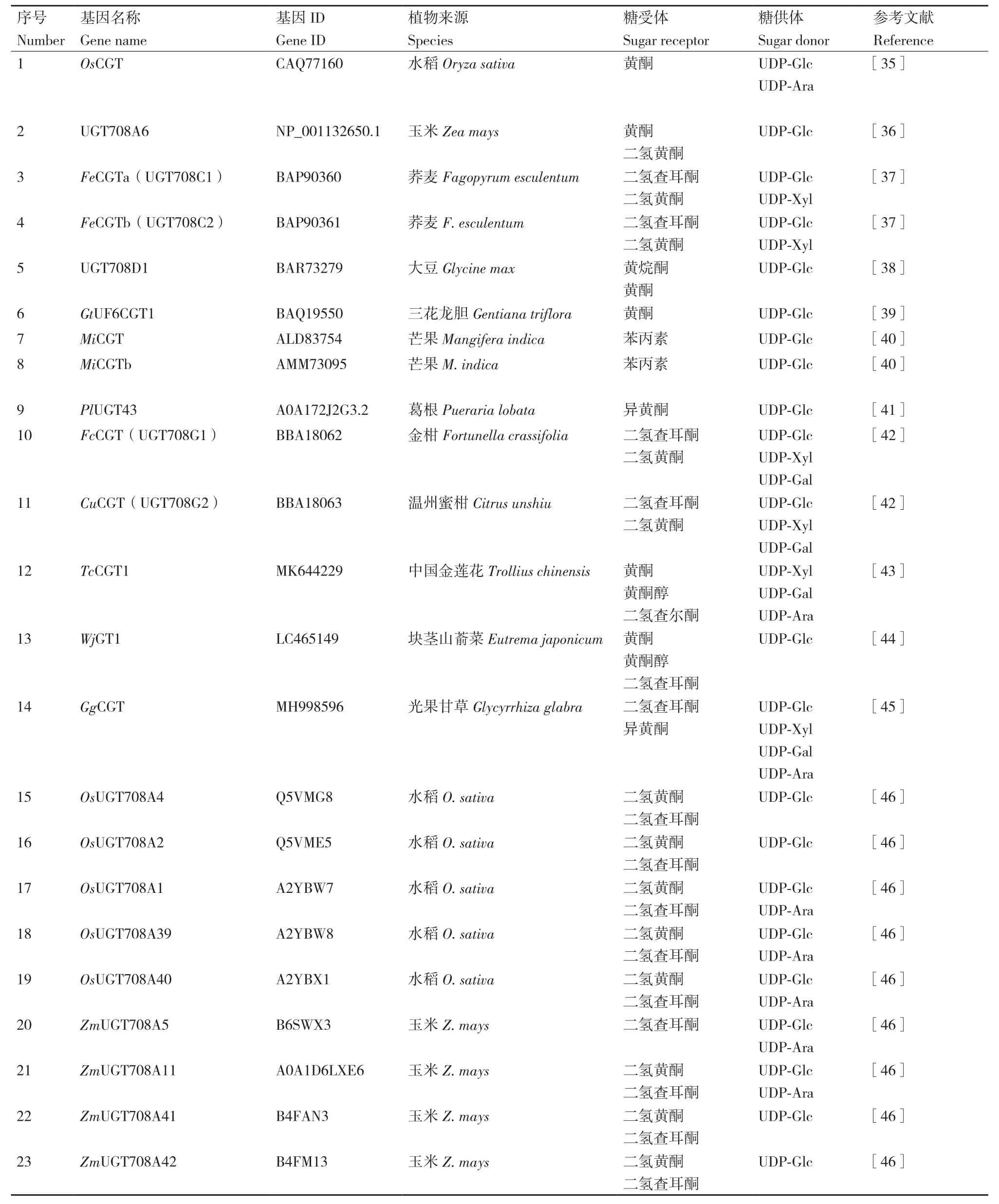
表1 已有報道的CGT基因研究匯總Table 1 Summary of reported CGT gene studies
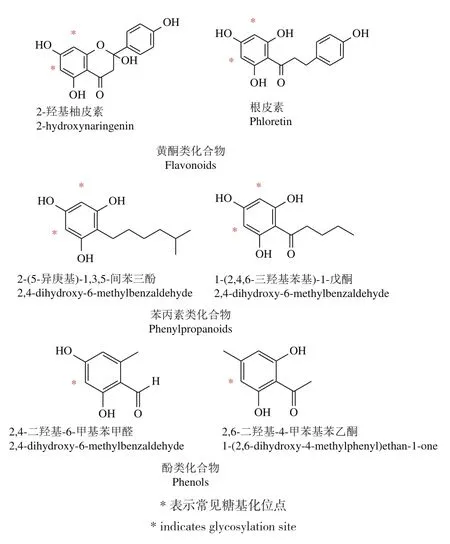
圖2 CGT介導糖基化的主要底物結構示例Fig.2 Example structures of major substrates for CGT-mediated glycosylation
2 C-糖基轉移酶的結構
2.1 CGT結構解析
對C-糖基轉移酶晶體結構的探索能夠為理解酶-底物的相互作用機制提供科學依據,同時也奠定了蛋白質改造工程的基礎[51]。2019年中國金蓮花TcCGT1晶體結構的解析,開啟了探索植物功能酶CGT晶體結構的大門[43]。截至目前,已有6種植物共15個CGT的實際晶體結構經過X-射線衍射解析(表2、圖3)。
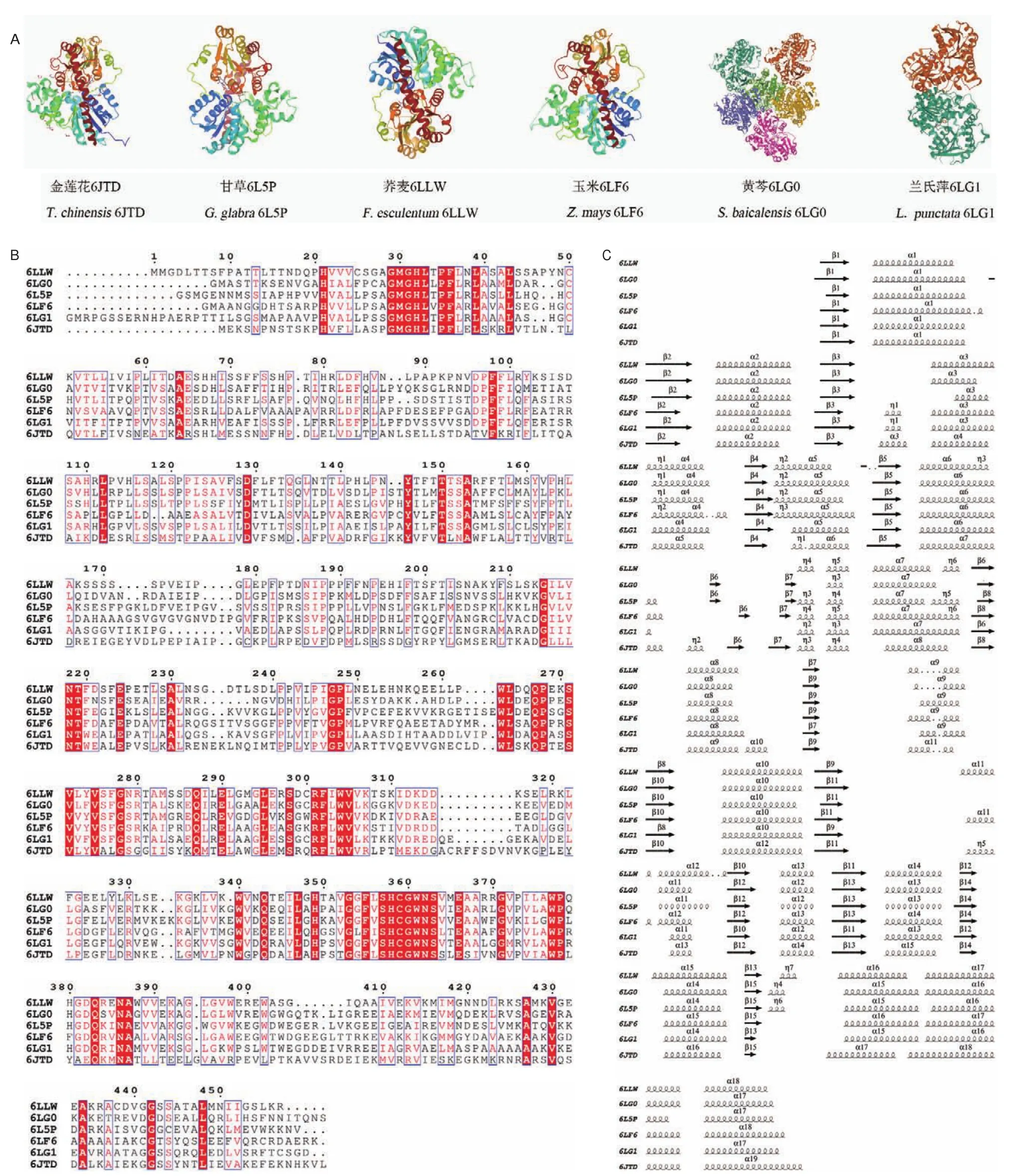
圖3 已有結構解析的植物CGT的蛋白結構Fig.3 Protein crystal structure of structurally resolved plant CGT

表2 已有解析晶體結構的CGT蛋白Table 2 Resolved crystal structures of available CGT proteins
CGT作為催化形成C-糖苷鍵的生物合成關鍵酶,相比于其他類型植物糖基轉移酶發現較少,其蛋白質三維結構與UGT大致相似,具有UGT普遍的結構特征,以GT-B折疊方式呈現:在單分子蛋白的狀態下,其蛋白結構含有兩個Rossmann折疊的β/α/β結構域,分為N-端結構(NTD)和C-端結構域(CTD),它們之間的聯系并非十分緊密,以一個面對的姿勢呈現,從而形成一個能夠與底物結合的細長裂縫。其中N-端結構域其保守性稍差,負責與糖基受體的結合;C-端結構域中相對保守的PSPG(putative secondary plant glycosyltransferase)區域則由44個氨基酸組合而成,糖基供體通過與PSPG區域的氨基酸殘基之間的多個氫鍵相互作用從而與糖基轉移酶穩定地結合[53-58]。
2.2 CGT底物雜泛性的結構基礎
現有研究發現,目前植物CGT的催化過程表現出底物雜泛性的特點,N-端結構域和C-端結構域之間所形成的底物結合口袋,是決定CGT糖受體和糖基供體結合性質的結構基礎[43]。底物在糖基轉移酶活性口袋中的立體結合構像,會影響雙功能糖基轉移酶催化性質的偏好性,即傾向于生成C-苷或O-苷。在中國金蓮花TcCGT1的催化研究中發現,寬敞的糖受體結合口袋有利于TcCGT1高效催化36種碳苷類化合物,基于晶體結構解析結果進行關鍵氨基酸殘基的定點突變,在具體催化時,對其寬敞的糖受體結合口袋周圍的殘基進行定點誘變,改變了立體構象,口袋寬敞時,更易接納較長基團,TcCGT1表現C-活性,口袋較小時迫使與較短的基團結合,故TcCGT1催化出O-苷產物。由底物的自發去質子化,可使得TcCGT1實現由C-糖基化向O-糖基化的功能轉變。即在糖基化底物相同的情況下,口袋寬敞時,更易接納較長基團生成C-苷產物,口袋較小時則被迫與較短的基團結合,催化生成O-苷產物[43]。
同時,對光果甘草中的雙C-糖基轉移酶GgCGT進行研究,采用X-射線晶體學方法對其進行結構解析(圖4),證明His27作為堿性氨基酸輔助底物脫質子,從而起到催化作用,確定了GgCGT催化雙C-糖基化反應的最佳結構單元為弗洛丙酮,其口袋的立體構象發生的位置偏轉導致糖基部分與周圍氨基酸形成氫鍵作用的強弱發生變化,影響酶與糖供體的結合作用強弱,進而決定糖基供體的偏好性順序[45]。
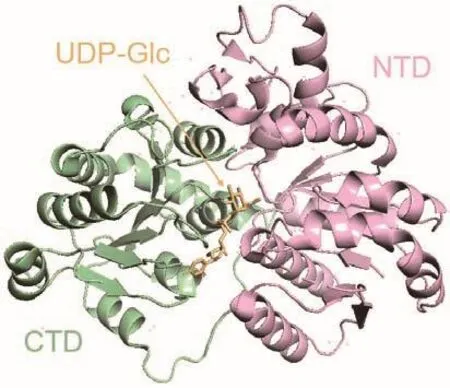
圖4 光果甘草GgCGT的晶體結構Fig.4 Crystal structure of GgCGT of Glycyrrhiza glabra
3 C-糖基轉移酶的應用
糖基化反應機制一直是植物代謝研究中的重要領域,2009年水稻中首個植物C-糖基轉移酶的發現拉開了植物C-糖基轉移酶研究的序章[35]。數十年來,對植物體內催化各種活性成分物質合成的糖基轉移酶進行了解析,其中催化黃酮類化合物的CGT為主,例如水稻 OsCGT[35]、蕎麥 FeCGTa 和 FeCGTb[37]、日本三花龍膽GtUF6CGT1[39]等。黃酮類化合物作為底物時,多進行6-C-糖基化,鮮見8-C-糖基化產物,而雙C-糖基轉移酶可以將這兩個位點同時或分步催化[59],例如柑橘類植物金桔和溫州蜜桔的FcCGT 和 CuCGT[42],鐵皮石斛的 DcaCGT[47],荷花的NnCGT1和NnCGT2[48]。有些CGT是既產生C-糖苷類,又可以產生O-糖苷類物質的雙功能糖基轉移酶,如玉米UGT708A6[36]。開展C-糖基轉移酶的應用研究,可以調控重要活性成分生物合成途徑,同時也是生物催化劑定向改造的基礎。
續表Continued
3.1 藥物發現與異源合成
隨著對糖基轉移酶的挖掘和認識的深入,其催化途徑逐漸明確,基于糖基化的藥物設計和開發日趨成熟。竹子和谷類作物的葉片中含有一類具有止痛功能的C-糖基化黃酮(C-glycosylated flavones)CGFs[60-63],優于阿司匹林等臨床藥物,對其合成過程中C-糖基轉移酶的解析[46],可為新藥發現鑒定基礎。蘆薈中新穎的C-糖基轉移酶AbCGT[50]能將缺乏乙酰基團的酚類化合物進行C-糖基化,且AbCGT具有底物雜泛性,可以通過化學酶法先合成苷元再合成C-糖苷的方式,實現多種新穎C-糖苷的化學-酶法合成,此結果對抗糖尿病創新藥物的發現及綠色制造提供了新的思路。葛根中主要藥用活性物質葛根素合成的關鍵酶PlUGT43是一類C-糖基轉移酶[41,64],該發現有利于實現葛根素的異源合成。
3.2 晶體解析與定向改造
通過在蛋白晶體結構中找到糖供體及受體結合所需的氨基酸殘基位點以及蛋白空間結構發生的變化闡明糖基轉移酶的催化機制,為設計有效的糖基化生物催化劑并定向改造提供基礎,定向改造不僅能夠提高其對底物的雜泛性,同時也能夠提高其催化活性[65]。
通過同源模擬和分子對接預測活性位點,再對其突變加以驗證,是判斷關鍵氨基酸位點的常用方法之一。大豆UGT708D1[27]驗證的3個殘基位點是預測的活性位點,取代關鍵殘基位點可破壞其催化活性,也可阻斷C-糖基轉移酶的催化活性,增強O-糖基轉移酶的催化的專屬性,能夠達到定向調控。芒果中發現的兩步催化香豆素MiCGTb[40]也驗證了關鍵氨基酸殘基位點,通過對MiCGTb進行突變,其糖受體槲皮素的糖基化活性大幅提升,且糖基化選擇過程更為嚴格[66]。
糖基轉移酶的綠色、高效,能夠為藥物的靶向合成提供新的方向,實際晶體結構的解析對明確催化機制有著重要的作用,進一步闡明糖基化活性和糖供體選擇性的分子催化機理,推動了C-糖苷類化合物的生物合成和以此為基礎的藥物研發進程。金蓮花 TcCGT1[43]、甘草 GgCGT[45]以及蕎麥CGT708C1[52]等晶體結構都已解析并應用。學者們還鑒定了玉米、蘭氏萍和黃芩等6種植物夏佛塔苷合成途徑中的12個關鍵CGT,并對部分酶的晶體結構進行了解析[49],為闡明植物糖基化次生代謝的進化機制提供了重要理論基礎。
4 展望
糖基化反應是天然產物生物合成的重要后修飾方式,一些植物次生代謝產物的糖基化反應由于其對生理的重大影響也備受關注,無論是在天然或非天然領域都發揮著不可或缺的作用,同時,一些重要的植物激素的糖苷化修飾在生理上也發揮著重要作用。因此解析植物糖苷類物質的生物合成途徑的關鍵酶功能結構,是推動這些化合物應用的基礎。
MiCGT進行定向進化來催化黃酮類化合物的糖基化,闡明了底物特異性和區域選擇性的起源[66]。此類糖基轉移酶的研究,對以改良植株和提高產量為目的定向改造糖基轉移酶提供依據,可以結合基因工程和蛋白質工程的技術手段,對糖基轉移酶進行基于活性氨基酸位點的挖掘與改造的理性設計,可以實現目標活性成分的定向合成和積累,以實現糖苷類產物的定向精準調控。
隨著C-糖基轉移酶的實際晶體結構被不斷解析,為設計高效、高特異性的生物催化劑和生物活性藥物提供科學依據,為進一步開發利用糖苷類化合物創造條件,為合成生物學提供最適元件,為天然產物異源合成奠定基礎。就目前植物C-糖基轉移酶的研究進展而言,研究最透徹的是黃酮類化合物的相關CGT,大量研究闡明了以黃酮、二氫黃酮為底物的催化過程,對苯丙素類、酚類化合物的研究還有待深入。
植物糖基轉移酶在改善天然產物活性方面有很大的應用潛力,以酶促催化合成為基礎的新型糖苷類產物的研究與開發為人們所期待,C-糖基轉移酶催化底物雜泛性的特點使糖苷類產物更具多樣化,能為人工模擬生物合成領域拓展新思路。隨著多種新型糖基轉移酶的發現和商業化生產使這些多樣化逐漸成為可能,更多優質新型糖苷類產物將會被挖掘并研制,對植物優良種質培育及為糖苷類化合物的異源高效合成有重要意義。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
