植物miR396-GRF模塊的生物學功能及其潛在應用價值
2023-01-05 08:45:28山琦賈惠舒姚文博劉偉燦李海燕
生物技術通報 2022年10期
山琦 賈惠舒 姚文博 劉偉燦 李海燕
(1.吉林農業大學生命科學學院,長春 130118;2.海南大學熱帶作物學院,海口 570228)
植物miRNA(microRNA)是一類具有調控作用的小分子RNA,其本身并不編碼功能蛋白,而是通過負調控靶mRNA行使功能。miR396是植物中保守存在的miRNA家族之一,并且植物中保守的生長調節因子(GRF)基因家族的多數成員已被證實是其作用的靶基因[1]。近年來的研究發現,miR396通過負調控生長調節因子GRF家族基因,可以影響植物葉片及花器官的形態建成、根和莖組織的伸展、種子的大小、數量等表型變化[2],進而影響植物的生物量及作物產量[3],并可改善植物吸收氮和磷肥的能力及植物耐受逆境脅迫的響應能力[4-5]。此外,最新的研究結果證明,GRF基因在基因工程技術的遺傳轉化過程中,不僅可以顯著提高外植體的再生效率,還可拓展受體植物的基因型[6]。因此,miR396介導的GRF調控(miR396-GRF)模塊在改良植物生物量、作物產量、抗逆能力及提高植物遺傳轉化效率方面,已展現出了潛在的應用價值。本文介紹了miR396-GRF模塊的作用模式,綜述了近年來miR396-GRF模塊的生物學功能及分子機制研究進展。為進一步深入研究miR396-GRF模塊及其開發應用提供思路和參考。
1 植物中的miR396與GRF基因及其互作模式
目前,在miRNA的序列數據庫(miRBase)中,共收錄了植物47個物種中的138個前體miR396(pre-miR396)和175個成熟體miR396(mat-miR396)序列(包括132個miR396-5p和43個miR396-3p序列)。序列比對分析發現,不同物種中的mat-miR396序列具有高度的同源性(圖1)。并且,大多數matmiR396序列與擬南芥中的ath-miR396a-5p和athmiR396b-5p兩種成熟序列形式相同,其二者在序列長度上一致,但末位具有一個不同的核苷酸堿基(圖1)。并且,在目前已有的報道中,這兩種matmiR396序列也已被證實相比miR396家族的其它成員,具有更顯著的表征作用。

圖1 不同植物物種中miR396基因家族成熟區序列比對分析Fig.1 Alignment analysis of mature region sequences of miR396 gene family in different plant species
生長調節因子GRF是一類保守存在于植物中的轉錄因子,在調控植物生長發育等過程中起重要作用。GRF蛋白的N端普遍含有QLQ和WRC兩個保守功能結構域(圖2)。在QLQ結構域中,除保守的Q/Gln-L/Leu-Q/GLn氨基酸外,還含有疏水性的酸性氨基酸殘基 F/Phe、Y/Tyr、L/Leu、E/Glu和P/Pro等,或其他理化性質類似的氨基酸(圖2-A)。這些氨基酸殘基可能在GRF蛋白與靶蛋白互作中起關鍵作用。在WRC結構域中,含有保守的W/Trp-R/Arg-C/Cys氨基酸殘基,以及由3個C/Cys和一個H/His組成的C3H motif,這些高度保守的氨基酸位點可能在決定GRF蛋白核定位和結合下游基因順式作用元件過程中起重要作用(圖2-B)[7]。值得注意的是,緊隨C3H motif區域之后,有一個保守的“RSRK-VE區”(圖2-B),該區域對應的核苷酸編碼序列大多高度保守,且與成熟的miR396序列近乎互補。目前已證實,miR396在此保守位點的“CGUUCAAGAA”和“AGCNUGUGGAA”之間剪切GRF基因的mRNA序列(圖3),進而負調控GRF基因的表達[1]。但是,也有少數GRF成員的“RSRK-VE”區對應的核苷酸序列不能與成熟miR396序列互補配對,因此不被miR396剪切調控。例如,在擬南芥9個AtGRF家族成員中,AtGRF5和AtGRF6雖含有保守的“RSRKVE”,但其核苷酸編碼序列卻不能與成熟miR396序列互補,因此不受miR396調控[8]。此外,植物中GRF蛋白家族的C端氨基酸序列高度可變,推斷其可能與GRF蛋白功能的多樣性或轉錄激活有關[9]。
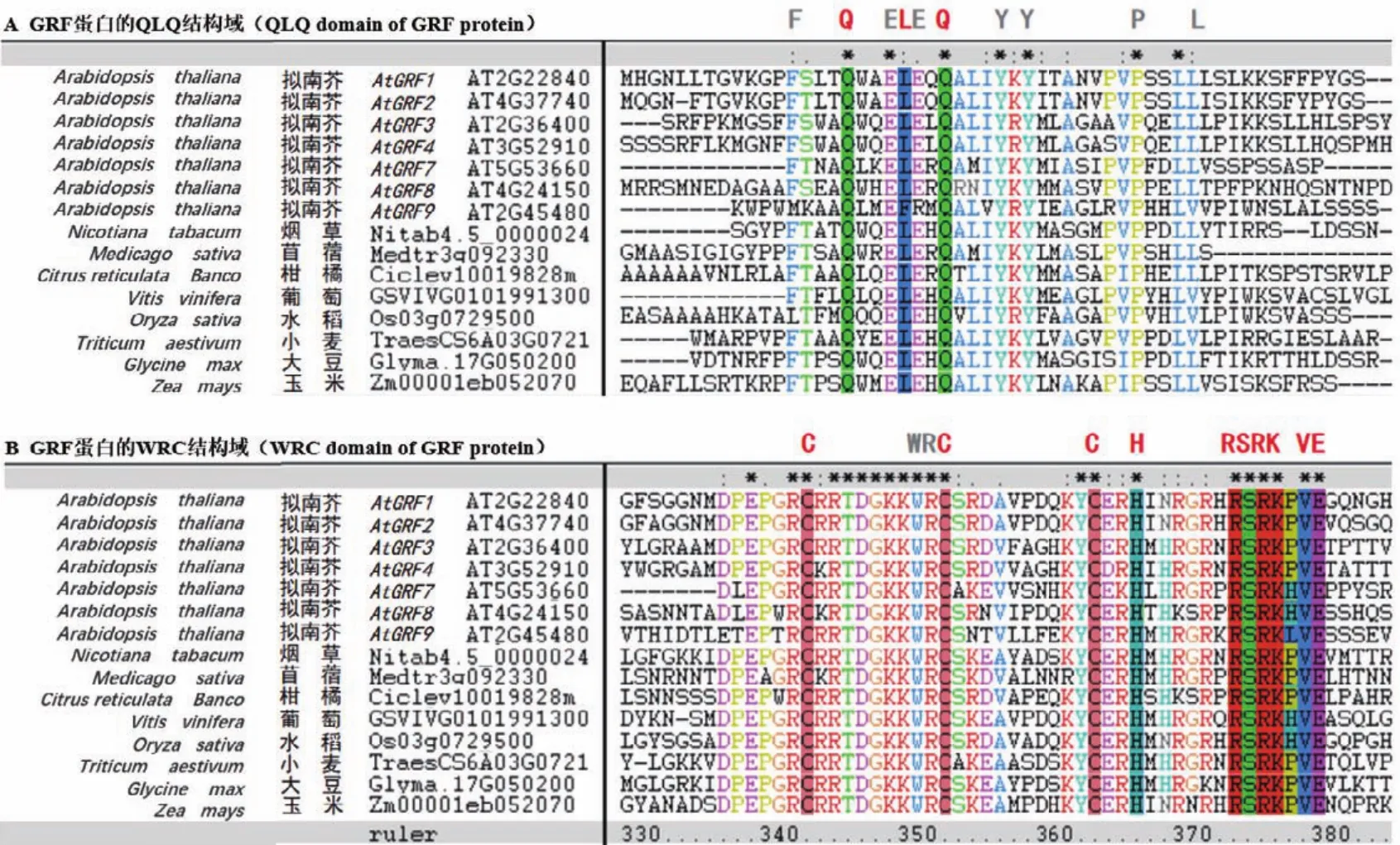
圖2 不同植物物種中GRF蛋白家族氨基酸序列的保守結構域比對分析Fig.2 Alignment analysis of the conserved domains of GRF protein family amino acid sequences in different plant species
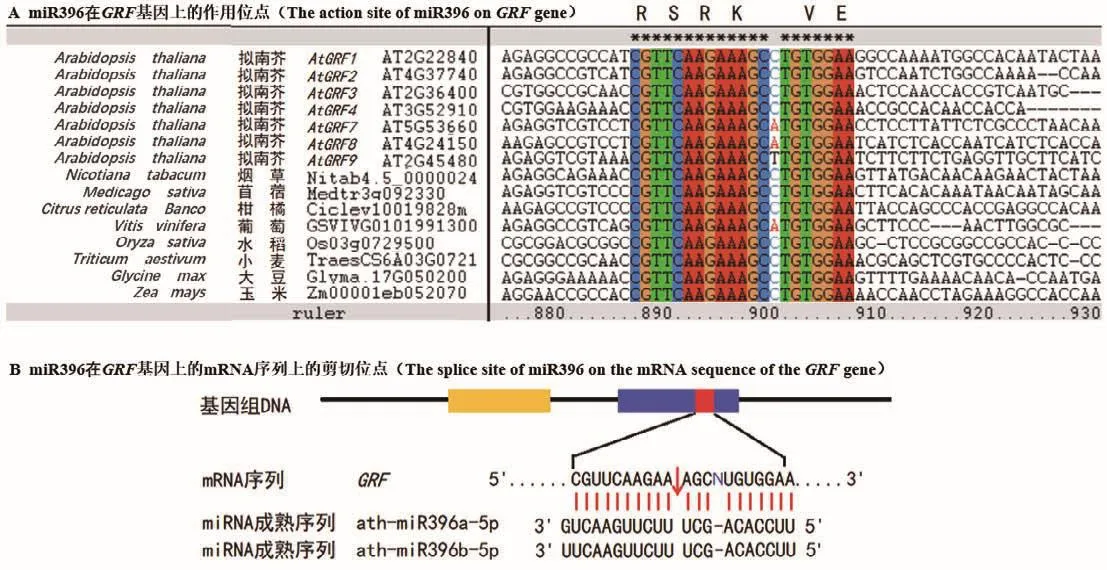
圖3 不同植物物種中miR396在GRF基因序列上的作用位點及剪切位點Fig.3 Action sites and splicing sites of miR396 on GRF gene sequences in different plant species
2 miR396-GRF模塊的生物學功能及應用研究進展
2.1 miR396-GRF模塊對植物生長發育及生物量的影響
自miR396-GRF模塊被發現以來,越來越多的研究證實了miR396-GRF模塊具有影響植物組織器官形態建成過程的調控功能。當突變體中的miR396基因被插入失活,或是miR396基因被敲除表達,以及在超表達GRF基因的植物材料中,miR396-GRF模塊通常表現出促進植物組織生長發育過程的趨勢。例如,在擬南芥中過表達內源的AtGRF3基因[10]、異源的煙草 NtGRF8[11]、白菜 RbGRF8[12],或是在水稻中過表達rOsGRF1基因(氨基酸同義突變,但核酸序列上消除了miR396靶位點)等[13],都促使轉基因植株變高,葉片變大,根系粗壯,種子及花器官發育延緩等形態變化[14-16]。并且,同義突變處理后的rGRF基因促進植物增長的效果更加顯著[10]。相反,當在植物中超表達miR396基因,或是突變體中的GRF基因被插入失活,以及GRF基因在植物材料中被抑制表達,miR396-GRF模塊通常會表現出抑制植物組織生長發育過程的趨勢。例如,在擬南芥[17]和水稻[18]等植物中過表達同源或異源miR396的轉基因株系,都表現出生長遲緩,植株矮小,根系變短,葉片也變得窄小的形態變化,這些表型與grf突變體相似[19]。因此,miR396與GRF基因在調控植物組織器官生長發育的過程中呈負相關關系,二者之間的靶向作用平衡制約著植物的形態表征。
此外,miR396-GRF模塊在一些經濟類植物中的表達調控,也表現出了一些有益的表型變化。例如,Wang等[20]在我國木材主要來源的楊樹(Populus alba × P.glandulosa)中,證實了PagGRF12a基因的表達調控作用可增加楊樹木質部的寬度,促進木材的建成。Liu等[21]在可用于生物燃料生產的植物柳枝稷(Panicum virgatum L.)中,證實了rPVGRF9基因的過表達可改變其生物量及原料質量。此外,在生菜(Lactuca sativa)中過表達LsaGRF5基因后,生菜葉片明顯變大,可顯著提升蔬菜的生物量產出[22]。以及在可用作牧飼材料的苜蓿[23]中,利用Target mimicry技術敲降表達miR396后的MIM396苜蓿材料,葉片的生物量也顯著增加,并且根系粗壯發達。綜上所述,miR396-GRF模塊的調控作用在提升植物生物量方面展現出了潛在的應用價值。
2.2 miR396-GRF模塊對產量等農藝性狀的影響
基于對植物生長發育過程的影響作用,miR396-GRF模塊也展現出了對作物產量等農藝性狀的影響能力。例如,miR396沉默體 MIM396水稻[14]、mir396ef突變體水稻[24]和GRF基因過表達后的水稻和小麥植株[25-27],都表現出了穗數的顯著增加,以及粒長和粒寬的增長現象,并且它們相對野生型能獲得更高的糧食產量。在果蔬植物番茄中,miR396通過調控不同的GRFs靶基因,也影響著番茄果實的多方面變化,過表達rGRF4和rGRF5的轉基因番茄可通過增加果皮厚度和果實心室數使果實重量明顯增加[28]。在油菜中,本源表達BnGRF2基因后的轉基因油菜的種子千粒重和含油量也得到顯著提升[29]。因此,miR396-GRF模塊的調控作用在提升作物產量及品質方面也展現出了潛在的應用價值。
另外,目前的研究報道顯示,miR396-GRF模塊還具有提升植物氮肥和磷肥利用率的能力。在缺氮條件下,過表達ZmGRF1基因的玉米可正向調控玉米氮素相關途徑基因的表達,促進氮肥吸收[30]。OsGRF4蛋白在水稻中的高水平積累也可以提高光合作用和氮肥利用效率[5]。此外,在高供磷條件下,過表達SlGRF9的轉基因番茄對土壤中磷的利用率也顯著提高。所以,miR396-GRF模塊在植物調控肥料吸收能力方面也有益于農藝生產。
2.3 miR396-GRF模塊在逆境脅迫響應過程中的調控作用
在農業生產中,植物體對逆境環境的適應能力也是影響田間生產的重要因素。近年來的研究表明,miR396-GRF模塊在調控植物響應逆境脅迫的過程中發揮著積極的調控作用。例如,MIM396番茄植株中由于miR396的下調表達,導致其在缺水脅迫下的蒸騰速率(E)降低,水分利用效率(WUE)顯著提高[31]。并且,在番茄中過表達AtGRF9基因,可提升根系蔗糖和干物質的含量,增強根系分泌質子的能力。所以,miR396-GRF模塊的調節作用對于番茄響應干旱等多種逆境脅迫過程都具有積極的意義[32]。在草坪草匍匐剪股穎(Agrostis stolonifera)中,超表達Osa-miR396c的轉基因植株在高鹽度環境中保水性增強、葉綠素含量相對較高、且細胞膜具有較好的完整性和鈉離子排除能力,由此表明其具有較強的鹽脅迫適應能力[33]。此外,OsGRF4基因突變體水稻在苗期的耐寒性增強,冷脅迫處理后幼苗的存活率顯著升高[3]。擬南芥經UV-B輻射后,miR396抑制AtGRF1/2/3表達,降低細胞增殖速率的同時抑制葉片生長,起到耐紫外線的作用。因此,miR396-GRF模塊在調控植物響應多種非生物脅迫的過程中具有積極意義。
在植物響應生物脅迫的過程中,miR396-GRF模塊也同樣具有調控功能。在大豆中,通過過表達或敲除實驗干擾miR396-GmGRF調控途徑,會抑制大豆孢囊線蟲幼體向成體的發育[34]。此外,稻瘟病毒嚴重影響著水稻的生長和產出。研究發現,OsamiR396d抑制OsGRF6/7/8/9表達,延緩水稻生長發育,減少病毒向新生組織擴散,從而提高了抵抗稻瘟病的能力[35]。因此,miR396-GRF模塊在調控植物響應生物脅迫的過程中也展現出了積極作用。
2.4 GRF提高植物再生效率的研究
轉基因受體材料遺傳轉化效率低及基因型的限制(即小麥等物種中僅有少數基因型的品種材料才可以被成功遺傳轉化)是基因功能研究及基于基因組編輯育種技術的主要瓶頸。2020年Kong等[6]研究發現,將AtGRF5及其同源基因導入甜菜、油菜、大豆和向日葵中均可有效提高轉化效率并產生完全可育的轉基因植株。緊隨其后,Pan等[36]進一步證實了在AtGRF5基因的輔助轉化作用下,西瓜外植體材料的轉化效率提高了40倍。同期,Debernardi等[37]研究發現小麥TaGRF4-TaGIF1嵌合體的轉基因表達顯著提高了4個品種小麥的再生效率,并且外植體再生過程周期短,不依賴激素調節,且轉化體沒有負面表型影響,從而拓展了小麥被轉化受體材料的基因型。此后,高彩霞團隊通過共轉化瞬時表達GRF-GIF嵌合體蛋白也證實了其有效性,并且小麥TaGRF4-TaGIF1嵌合體與基因編輯體系共表達也被證實可以高效地獲得編輯成功的突變體材料[38]。目前已被證實,GRF基因及其嵌合體相比以往使用的提高遺傳轉化效率的生長調節基因如BBM、WUS、IPT等更加有效[39]。顯然,GRF基因在改善雙子葉和單子葉植物外植體再生效率,及提高基因編輯效率方面都展現出了誘人的應用前景。
3 植物miR396-GRF模塊作用的分子機制研究
3.1 GIF作為輔激活因子可增強GRF蛋白的活性及功能
目前,在多篇研究報道中,都證實了不同物種中的 GRF 和 GIF 之間存在著蛋白互作關系[7,20,40],GIF蛋白也由此而得名為GRF蛋白相互作用因子(GRF-interacting factor,GIF)。Lu 等[13]研究發現,過表達OsGIF1基因的水稻沒有顯著的表型變化,而OsGIF1基因敲除后的水稻表現為植株矮小等表型形態,與OsGRF1基因敲除后的水稻形態相似。并且,mOsGRF1基因過表達和OsGIF1基因敲除的雜交系水稻植株也呈現矮小狀態。因此推斷,GIF蛋白雖然不能獨立行使功能,但確是生長調節因子GRF調控植物生長發育過程中不可或缺的因素。
進一步的深入研究表明,GRF和GIF蛋白首先在細胞核內形成功能復合物,然后GIF蛋白負責招募SWI/SNF染色質重塑復合物,借助染色質重塑酶BRM,利用ATP水解產生的能量,破壞組蛋白與DNA的相互作用,使DNA和組蛋白之間變得松散,以便作為轉錄因子的GRF蛋白與順勢作用元件結合并激活或抑制下游目的基因的轉錄[9,39],進而發揮各種生物學功能。因此,GIF作為GRF蛋白的輔激活因子,具有增強GRF蛋白活性的功能。
此外,植物細胞的去分化和再生能力,直接影響著植物外植體材料的遺傳轉化效率。一般來說,分化細胞具有緊密的染色質狀態,而去分化過程使染色質變為開放松弛狀態[39]。由于GRF-GIF復合體可以使染色質重塑而變得松散,因此可以有助于提升植物外植體材料的再生能力和遺傳轉化效率。
3.2 miR396-GRF模塊調控細胞增殖的機制研究
目前已有的研究結果顯示,在植物成熟的葉片、根和莖等組織中,miR396的表達量相對較高,GRF基因的表達量相對較低;而在根尖、花芽和幼嫩的葉片等初生組織中,miR396的表達量相對較低,GRF基因的表達量卻相對較高[41]。
其中,KNOX蛋白是促進莖尖組織分生的關鍵因子。Kuijt等[42]證實,水稻OsGRF3和OsGRF10基因通過與KNOX家族基因啟動子區的相互結合,從而抑制OsKN2基因的表達,并影響植物分生組織的發育。
另外,在植物中過表達miR396或敲除GRF基因可觀測到植株矮小,器官發育延緩等現象,但這種變化可以被GRF基因的表達所恢復[14-16]。細胞水平的檢測結果證實,上述轉基因材料葉片單位面積內的表皮細胞數量顯著減少,細胞分裂增殖受到了抑制[1,19]。轉錄組測序和熒光定量PCR分析結果顯示,miR396-GRF模塊的遺傳轉化材料中,一些與DNA 復制、細胞分裂和細胞周期調節等有關的基因具有顯著表達差異,如MAPK、CYC家族基因等[29,33]。因此說明,miR396-GRF 模塊通過控制植物組織細胞的分裂增殖過程[19,33],進而影響著植物組織的生長發育過程。
3.3 miR396-GRF模塊的激素調節機制研究
植物激素是植物細胞接受環境信號誘導產生的活性物質。在細胞分裂分化、開花結實、成熟衰老等多個方面,植物激素對植物的生長發育過程起著重要的調控作用[43-44]。已有的研究報道表明,miR396-GRF模塊調控植物生長發育的過程與赤霉素(GA)、吲哚乙酸(IAA)等激素調節及合成過程有著密切關系(圖4)。Zhang等[24]對水稻雙突變體osmir396ef的葉片進行了液相質譜色譜聯用及代謝組分析,發現GA3,GA4,GA7水平以及GA合成前體甲羥戊酸(mevalonic acid,MVA)水平均明顯提高,由此說明,miR396-GRF模塊間接影響著IAA的生物合成。此外,郭涇磊等[45]分析發現,在小麥TaGRF基因的啟動子區存在大量IAA的響應元件,在外源施加IAA處理后,可顯著上調TaGRF基因的表達,進而影響小麥分蘗。此外,在水稻和擬南芥中,都證實了AtGRF基因的下調表達,會進一步調控IAA生物合成途徑的關鍵基因,如YUC和ARF基因等,進而影響IAA的生物合成。因此,GRF基因的表達受到了IAA的誘導調控,GRF基因的表達又影響IAA的生物合成。綜上,miR396-GRF模塊參與了植物激素的調節過程,但大多缺少直接聯系的作用證據,還需進一步開展深入的研究。
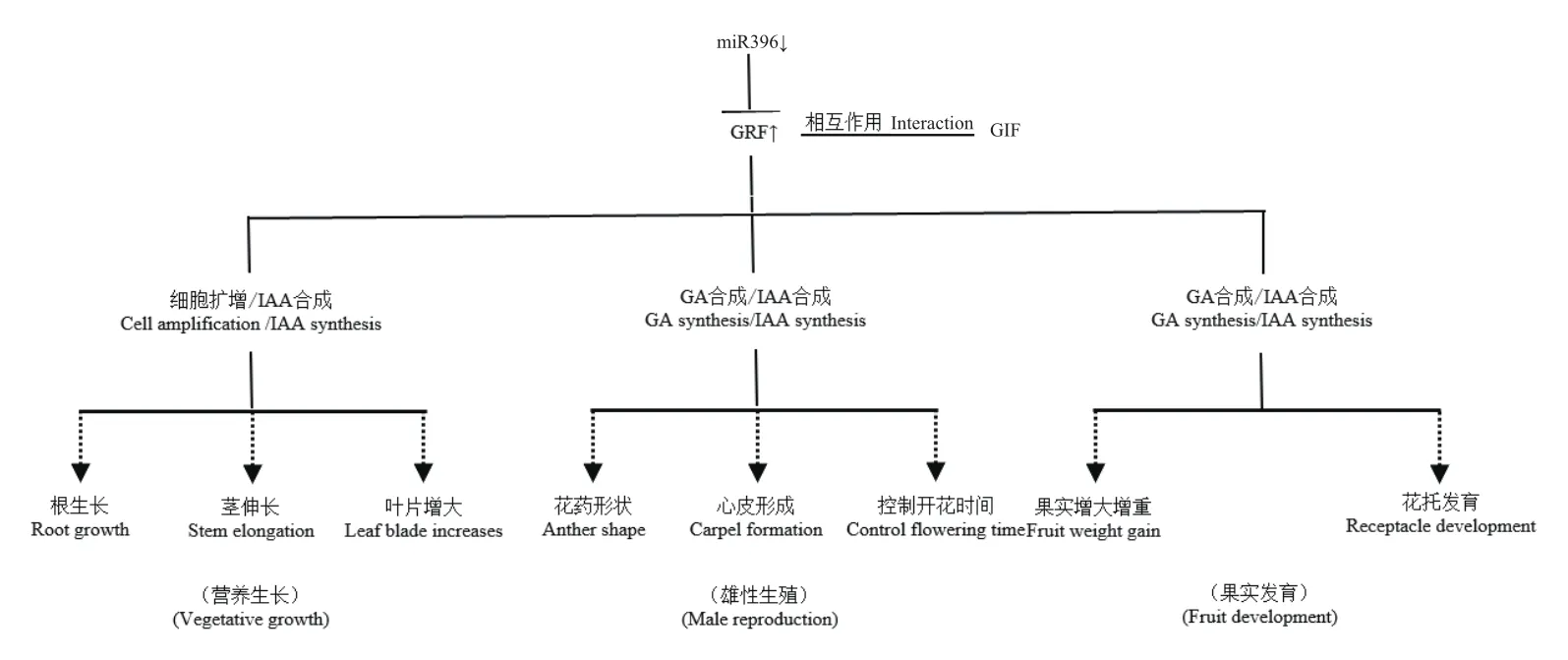
圖4 miR396-GRF模塊調控植物生長發育的過程Fig.4 Process of miR396-GRF module regulating plant growth and development
3.4 miR396-GRF模塊調控植物逆境脅迫響應的機制研究
植物由于其固著的生活方式而不得不連續暴露于可能存在各種逆境脅迫的環境之中。為應對環境脅迫,植物進化出了各種復雜的防御機制,包括信號傳感和轉導途徑、調控下游脅迫反應相關功能基因的表達等,從而通過氣孔關閉、細胞生長、光合作用抑制、可溶性物質積累等各種生物過程,保護植物免受損害并增加生存機會。有關miR396-GRF模塊遺傳轉化材料的轉錄組及Go富集分析發現,除了與細胞分裂有關的基因外,還存在很多與表達調控、激素合成、抗氧化還原反應等各種與逆境脅迫響應有關基因的差異表達[33]。因此說明,miR396-GRF模塊在植物響應逆境脅迫的過程中,也發揮著重要的調控作用。
根據目前研究獲得的數據表明,miR396-GRF模塊在植物中響應逆境脅迫的潛在信號通路大致為:在逆境脅迫下,miR396-GRF模塊被誘導轉錄后,首先控制蛋白激酶和轉錄因子等調節蛋白;然后這些調節蛋白激活或抑制下游逆境響應相關的功能基因,進而響應逆境脅迫過程(圖5)。例如,Yuan等[33]通過RT-PCR分析發現,在miR396的轉基因匍匐翦股穎中,WRKY、MADS和AP2等調節作用基因被下調表達,進而下游Na+逆轉運蛋白AsSOS1的作用被激活,從而影響了植株的耐鹽脅迫響應能力。Vall等[46]研究發現,GRFs的下調表達將導致控制活性氧(ROS)產生的乙烯反應因子ERF1基因的上調,從而阻斷乙烯反應,降低抗氧化酶活性。Kim等[47]的研究揭示,擬南芥AtGRF7基因可直接作用到與脫水反應有關的DREB2A基因的啟動子區,并抑制其表達,從而導致atgrf7突變體在高鹽與干旱條件下的耐受性增強。Fracasso等[31]研究結果顯示,MIM396番茄植株中,GRF基因的上調表達可刺激表皮細胞增殖,導致氣孔密度增加,氣孔變小;并可促進茉莉酸(JA)和γ-氨基丁酸(4-aminobutyric acid,GABA)的積累,從而調節氣孔關閉和維系光合作用過程,改變植物的逆境適應能力。總之,關于miR396-GRF模塊的研究有很多報道,但是其在植物中響應脅迫的調控網絡在很大程度上仍然是未知的,也需進一步開展深入的研究工作。
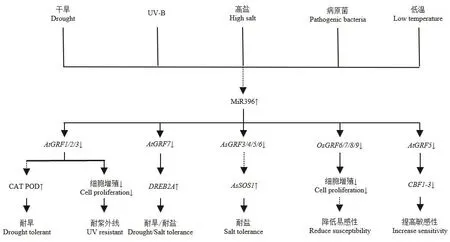
圖5 miR396-GRF調控模塊在脅迫響應過程中的可能調控途徑Fig.5 Feasible regulation approach of miR396-GRF module in the process of stress response
4 結語
目前已有的研究結果表明,miR396-GRF模塊不僅可以調控水稻及小麥等的株高、分蘗、穗數、籽粒大小及灌漿過程而影響作物產量[25-27]、提高油菜籽的出油率[29];還可改善番茄的抗旱能力[31],草坪草的耐鹽能力[33]及水稻的耐寒能力[3]等,并影響大豆感染孢囊線蟲[34]及水稻感染稻瘟病毒的過接證據。第四,miR396-GRF模塊在影響植物生長發育方面的研究報道進展較快,但是關于其影響植物逆境脅迫響應的研究相對薄弱。第五,miR396-GRF模塊調控作用的分子機制還很不清楚,缺乏調控途徑之間的直接關聯證據。第六,在大豆、玉米等重要物種中及同一物種中的不同基因型品種中,GRF基因可提升遺傳轉化效率的有效性正亟待實驗證實,與基因編輯模塊的聯合應用更有待開發。
綜上,miR396-GRF作為植物分子育種領域的候選基因,進行實際應用之前還需要很多的研究準備工作。建議加強miR396-GRF模塊在標的物種中的全方面調控作用研究,深入闡明其調控作用的分子機制;并大力開發高效有益的miR396-GRF基因用于植物品種改良,快速推進GRF基因在遺傳轉化和基因編輯系統中的有效運用。在此,本文僅為進一步深入研究miR396-GRF途徑及其開發應用整理思路和提供參考。程[35]。此外,GRF基因被證實能有效提升小麥等植物基因轉化及外植體再生效率[39]。因此,miR396-GRF模塊在提高作物產量、改良植物抗逆能力及提升植物遺傳轉化效率方面都展現出了誘人的應用前景。
然而,近年來miR396-GRF模塊雖然得到了研究者們的更多關注,有關miR396-GRF模塊生物學功能的研究報道也層出不窮,但在其研究發展的道路上和真正被開發應用之前,還存在很多值得注意的地方和需要解決的問題。例如,第一,miR396-GRF模塊調控途徑在植物界的不同物種中保守存在,在每個植物物種中,miR396和GRF家族基因的成員數都普遍較多,存在功能冗余情況。第二,miR396-GRF模塊中的同一基因在不同物種中,或同一物種的不同家族基因成員之間,可能存在著顯著的功能差異,甚至相反的影響。例如,TaGRF4 與ZmGRF10因缺少C端轉錄激活結構域,由此它們都抑制葉片的伸長生長[45,48];miR396轉基因煙草抗旱能力增強[49],而miR396轉基因擬南芥抗旱能力降低[50],這可能與物種之間的差異有關。第三,目前miR396-GRF模塊基因的功能更多的是在模式植物中異源表達被證實,大多缺乏在標的物種中的直
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
