嘌呤合成途徑基因VAL1超表達水稻光合調控生理機制研究
2023-01-16 02:41:04李玲依焦穎瑞胡健楊仕會曹紅宇張紅梅王兵兵馮萍凌英華張婷何光華姚賀盛
西南大學學報(自然科學版) 2023年1期
李玲依, 焦穎瑞, 胡健, 楊仕會, 曹紅宇, 張紅梅, 王兵兵, 馮萍, 凌英華, 張婷, 何光華, 姚賀盛
西南大學 農學與生物科技學院,重慶 400715
光合作用是產量形成的基礎,從能量轉化角度講,影響水稻產量潛力的主要因素包括:光吸收量、光能利用效率和收獲指數[1],而當前水稻收獲指數已接近理論上限[2],難以依靠優化收獲指數來大幅提高水稻的產量.因此,提高光合適應能力,促進生育前期獲得較高光合物質基礎,在維持較高收獲指數基礎上進一步提高光能利用效率以形成較高的生物量是實現水稻高產的重要途徑[3-5].
西南地區是我國重要的水稻栽培區,但生育前期長期的陰雨寡日照、自然光照弱等客觀因素致使冠層內葉片光合速率低,難以在生育前期獲得較好的光合物質基礎.提高低光光適應能力進而提升低光光能利用效率成為該區域水稻光合適應性研究的重要目標.水稻低光光能利用效率具有高度遺傳特性,是影響水稻生物量形成的關鍵因素之一,且育成水稻品種之間低光光能利用效率存在很大變異,尚未在人工馴化過程中受到強烈選擇[6].因此,低光光適應能力強的基因及資源材料挖掘成為進一步提高水稻產量的重要研究方向.
光合有效輻射(PPFD)不僅影響植物葉片的形態學特征變化(如光合色素質量分數)[7-8],還會對葉片的光合生理特征(如光合電子傳遞效率、光合碳同化效率)產生顯著的調節作用[9-10].這些生理特征均會影響葉片的光合能力和光能利用效率.除單葉水平光合效率外,保持合理的單株葉片面積也是提高作物群體光合和光能利用效率的重要基礎,適宜的單株葉片面積可以顯著改善群體光合物質的產出,進而促進作物產量的形成[11].
前期研究發現,嘌呤合成途徑基因(Virescent-Albino Leaf 1,VAL1)編碼蛋白VAL1,是嘌呤生物合成途徑中的關鍵基因,參與調節水稻葉片發育過程中的葉綠體發育、葉綠素代謝和細胞分裂.通過將VAL1在野生型材料中超表達,獲得了光合速率顯著提高且表型穩定的水稻株系(VAL1-OE)[12].本研究測定結果顯示,VAL1-OE水稻具備同時適應低光和高光環境的典型生理特性,這表明VAL1-OE水稻同時擁有高效利用低光和高光的能力.在自然界中,很少發現同時具備適應低光和高光能力的天然植物[13],這一特性使VAL1-OE水稻成為開展光合適應性研究的優勢材料.本文擬通過研究VAL1-OE與野生型材料在光合色素合成、光能吸收及光合碳同化效率等生理途徑的差異,揭示VAL1-OE水稻增強光合適應能力及提高光合碳同化速率的調控機制,為挖掘寡日照地區水稻增產潛力,篩選光照廣適稻種資源提供參考.
1 材料和方法
1.1 試驗概況
試驗于2020年4月至8月在西南大學水稻研究所歇馬(北緯N29°46′4.20″,東經E106°21′53.07″)試驗基地進行.試驗選用縉恢10號野生型(WT)和VAL1超表達水稻(VAL1-OE)為試驗材料,在抽穗期進行株型特征、葉片表型、光合作用及光能利用效率的測定與分析.試驗選取2個水稻材料,于4月育秧,5月6日移至田間插秧,栽插規格為株行距16.67 cm×26.66 cm,每個試驗小區10行×10列,共100株,管理措施同大田.試驗區3 m高處運用防鳥網完全覆蓋,以防止鳥類采食影響試驗結果.
1.2 樣品采集及處理
大田試驗中,選擇分蘗期水稻分別進行光合測定、色素質量分數測定和干物質分配測定.
水稻成熟后單獨收種考種,水稻考種包括穗長、穗實粒數、穗空殼數等基本產量構成指標.
1.3 測定項目及方法
1.3.1 株型指標測定
在抽穗期對水稻進行取樣,每個試驗小區取代表性植株3株,洗凈根部泥土,然后測量相關指標.測量指標包括株高、分孽數量、比葉質量(LMA)、單株葉面積(S)、葉片干質量(W)以及葉綠素質量分數,其中,單株葉面積(S)計算公式為
S=W/LMA
1.3.2 葉綠素質量分數的測定
光合色素質量分數測定方法:用切割法切下部分葉片(用游標卡尺量葉片的長度并計算面積),然后加入10 mL葉綠素提取液,黑暗環境浸提48 h,每隔12 h混勻1次,最后用紫外分光光度測量吸光度.測定時提前20 min將紫外分光光度預熱,待預熱結束后,以空白的葉綠素提取液作為對照,分別在645 nm,663 nm兩個波長下測定提取液的吸光值(OD)[14].
Chla= 12.2OD663-2.81OD645
Chlb= 20.13OD645-5.03OD663
Chl(a+b)=17.32OD645+7.18OD663
OD645,OD663分別表示在波長645 nm,663 nm下測得的吸光值,Chla,Chlb,Chl(a+b)分別表示葉綠素a,葉綠素b和總葉綠素的濃度.
單位面積葉綠素質量分數(Cc,mg/m2)
Cc=C×V×10/S
式中,C為相應葉綠素的濃度(mg/mL),V為提取液體積(mL),S為測試樣品的面積(m2).
1.3.3 比葉質量的測定
用切割法切下部分葉片(用游標卡尺量葉片的長度并計算面積),先在烘箱中105 ℃殺青30 min,再在65 ℃烘至恒質量,最后用百分之一天平稱質量.
1.3.4 光響應曲線(A-PPFD)的測定
水稻葉片A-PPFD的測定運用GFS-3000和雙通道熒光儀(Chlorophyll Fluorescence & P700 Measuring System,DUAL-PAM-100,Walz,Germany)聯用系統進行測定.
選取冠層上部倒1葉片為測試樣本,樣本葉片進行暗適應30 min后進行測定.測定時,溫度為27 ℃,相對濕度為70%,CO2濃度為400 μmol/mol,流速為400 μmol/s,光合有效輻射梯度設置為1 895,1 523,1 218,980,621,390,221,116,85,55,30,17,7和0 μmol/(m2·s).選取1 523,980和390 μmol/(m2·s) 3個光強的點進行比較.
在測定A-PPFD時,葉綠素熒光和P700進行同步測定,光系統I和光系統II的電子傳遞速率ETRI和ETRII分別由試驗儀器直接測定.
1.3.5 干物質、產量及其構成因素的測定
干物質的測定:將水稻各器官分別裝于紙質信封,置于烘箱中105 ℃殺青30 min,然后在65 ℃烘干至恒質量后,用百分之一天平稱質量.
產量及其構成因素的測定:水稻成熟前,調查每個小區的單株有效分蘗數,每個小區取10株進行考種,考種指標包括穗長、一次枝梗、二次枝梗、每穗實粒數和癟粒數,計算結實率和單株產量.
1.4 數據分析
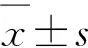
2 結果與分析
2.1 基因表達和植型表型分析
通過將VAL1基因在野生型材料縉恢10號(WT)中進行超表達獲得了表型穩定的水稻株系VAL1-OE(圖1a),在分蘗期對野生型和VAL1-OE水稻葉片葉綠體發育和光合相關基因進行了qRT-PCR測定分析(圖1b).試驗結果表明,在光能吸收方面,VAL1-OE水稻葉片中編碼捕光復合體II葉綠素a/b結合蛋白基因(LhcpII)的轉錄水平較野生型提高了75%; 在電子傳遞方面,VAL1-OE水稻葉片中編碼PS I P700葉綠素a脫輔基蛋白A1基因(psaA),PS II D1蛋白基因(psbA),細胞色素f脫輔基蛋白基因(petA)和細胞色素b6-f復合體小亞基基因(petG)的轉錄水平分別較野生型提高228%,355%,151%和258%; 在光合羧化固定方面,VAL1-OE水稻葉片中編碼核酮糖-1,5-二磷酸羧化酶/加氧酶大亞基基因(rbcL)、核酮糖-1,5-二磷酸羧化酶/加氧酶小亞基基因(RbcS)和葉綠體ATP合成酶α亞基基因(atpA)的轉錄水平分別較野生型提高了370%,131%和375%.由此可知,VAL1-OE水稻葉片中調控光吸收、傳遞與轉化過程主要基因的表達均顯著上調(p<0.01).
具有良好的株型特征是作物實現高光效的重要前提,分別對野生型(WT)和VAL1-OE水稻材料的株高、分蘗數量、比葉質量和單株葉面積等性狀進行了差異顯著性分析(圖2).結果表明:VAL1-OE水稻材料的比葉質量(LMA)顯著高于野生型,較野生型提高了36.95%(p<0.01,圖2c);VAL1-OE水稻材料的株高、分蘗數量和單株葉面積顯著低于野生型(p<0.05),較野生型分別降低了6.75%(圖2a),14.29%(圖2b),和31.13%(圖2d).
2.2 光合表型分析
VAL1超表達材料(VAL1-OE)與野生型(WT)之間的色素質量分數差異存在統計學意義(圖3).試驗結果表明,VAL1-OE水稻材料單位面積葉綠素a,葉綠素b和葉綠素(a+b)質量分數均顯著高于野生型(p<0.05),分別較野生型提高了20.36%,8.09%和17.86%.
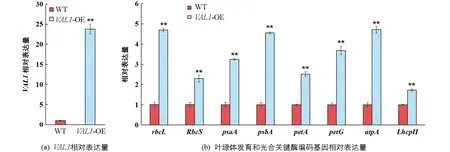
*表示p<0.05,**表示p<0.01,差異有統計學意義.圖1 相關基因差異表達分析
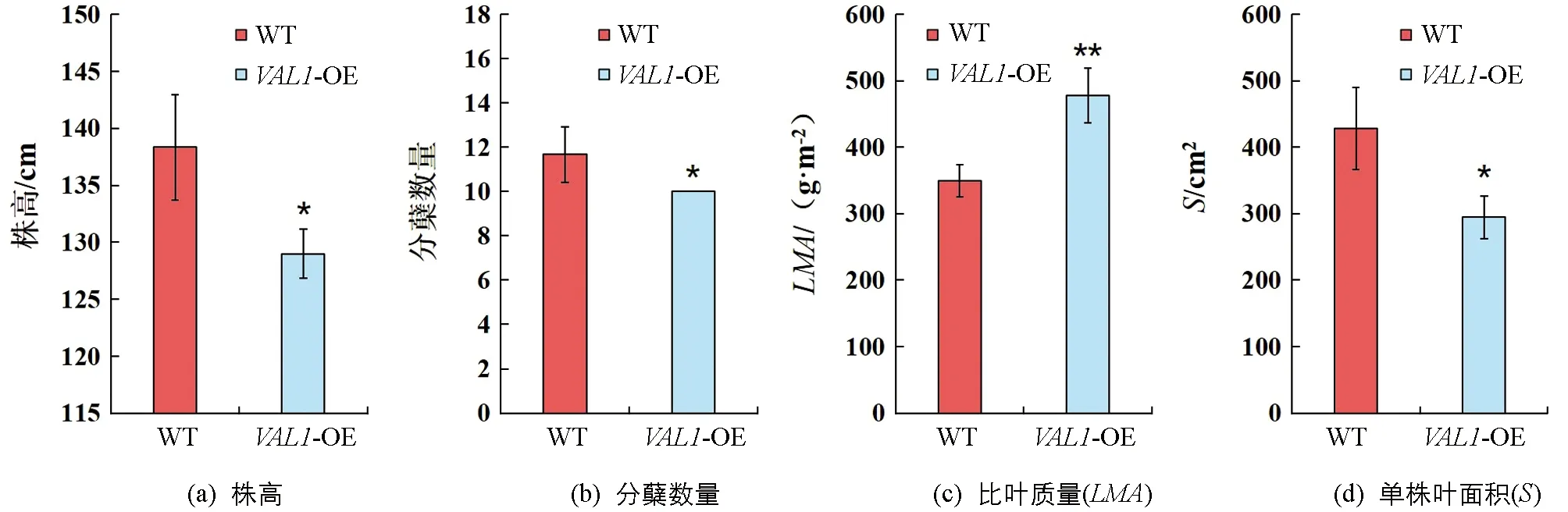
*表示p<0.05,**表示p<0.01,差異有統計學意義.圖2 株型表型差異分析

*表示p<0.05,**表示p<0.01,差異有統計學意義.圖3 光合色素質量分數差異分析
電子傳遞是光合作用進程中的重要環節,光合電子傳遞速率的不同也可能會對最終的光合值以及產量產生影響.為探究VAL1-OE水稻材料與野生型(WT)之間光合電子傳遞的差異,分別在390,980和1 523 μmol/(m2·s)的光照強度下對野生型和VAL1-OE水稻材料的光系統II和光系統I電子傳遞效率進行差異顯著性分析.試驗結果表明,在不同光照強度下,VAL1-OE水稻材料光系統II和光系統I電子傳遞效率均顯著高于野生型(圖4).VAL1-OE水稻材料在390,980和1 523 μmol/(m2·s)的光照強度下光系統II電子傳遞效率分別較野生型提高了18.06%,30.76%和33.09%(p<0.05,圖4a); 光系統I電子傳遞效率分別較野生型提高了16.14%,26.58%和30.28%(p<0.05,圖4b).
光合碳同化是植物干物質累積的基礎,對植物生長發育具有重要的調控作用.VAL1-OE水稻材料與野生型(WT)在不同光強下的光合碳同化效率差異有統計學意義.試驗結果表明,在不同光照強度下,VAL1-OE水稻材料的凈光合速率均顯著高于野生型(p<0.01).在390,980和1 523 μmol/(m2·s)的光照強度下分別提高了82.47%,100.43%和97.01%(圖5).

*表示p<0.05,**表示p<0.01,差異有統計學意義.圖4 光合電子傳遞差異分析
2.3 光能利用效率和物質累積及產量構成分析
VAL1-OE水稻材料與野生型(WT)在不同光強下光能利用效率差異有統計學意義(圖6).數據結果表明,在不同光照強度下,VAL1-OE水稻材料的光能利用效率均顯著高于野生型(p<0.01),在390,980和1 523 μmol/(m2·s)的光照強度下的光能利用效率分別較野生型提高了82.47%,100.43%和97.01%.VAL1-OE水稻材料與野生型的考種數據結果表明,VAL1-OE水稻材料的抽穗期干物質量、穗長、實粒數、癟粒數、穗數、千粒質量和總產量與野生型差異無統計學意義(表1).
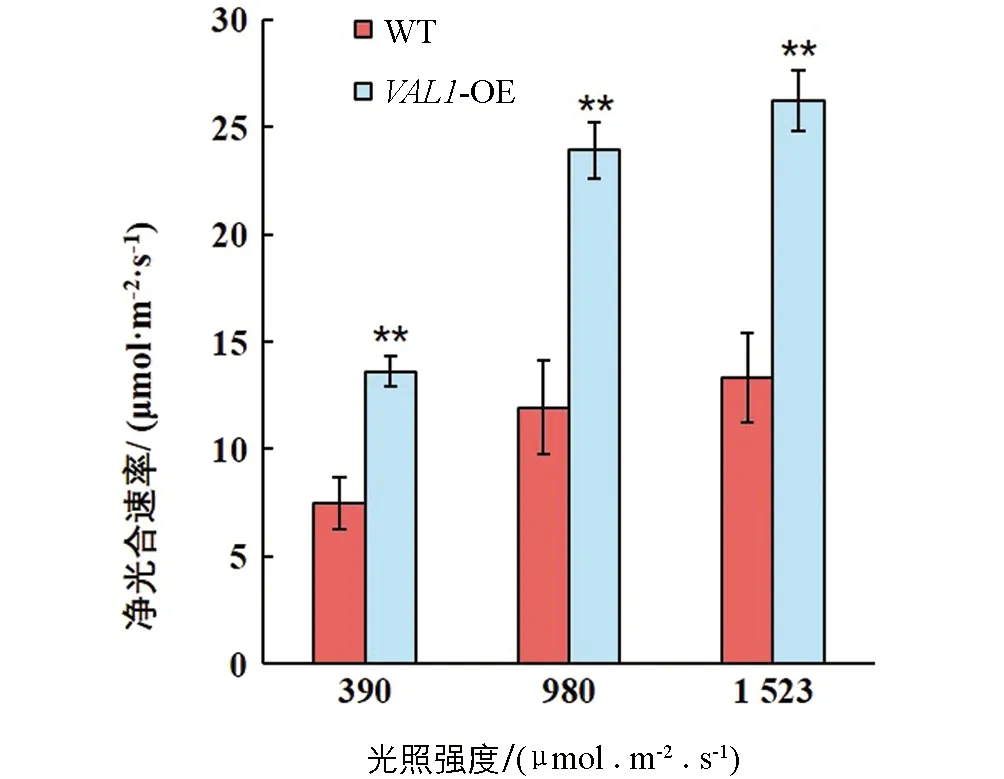
*表示p<0.05,**表示p<0.01,差異有統計學意義.圖5 不同光強下光合碳同化效率差異分析

*表示p<0.05,**表示p<0.01,差異有統計學意義.圖6 不同光強下光能利用效率(LUE)差異分析

表1 物質累積和產量構成因素
3 討論
3.1 超表達VAL1水稻優化葉片光能吸收、電子傳遞和碳同化是提高光合作用和光能利用效率的關鍵
光吸收能力的提高是優化光合作用的前提.葉綠素a/b結合蛋白圍繞光系統II(PS II)的反應中心作為主要的外部天線[15],能把接受的光能量快速傳導至反應中心,參與光能轉化與傳遞,以及對各種環境的適應等過程,對植物的光合作用起到關鍵作用[16].研究表明,在VAL1-OE超表達水稻葉片中,捕光復合體II葉綠素a/b結合蛋白基因(LhcpII),編碼PS I P700葉綠素a脫輔基蛋白A1基因(psaA),PS II D1蛋白基因(psbA),細胞色素f脫輔基蛋白基因(petA)和細胞色素b6-f復合體小亞基基因(petG)的轉錄水平均顯著提高(圖1),這些基因轉錄水平的提高有利于提升水稻葉片葉綠素a和葉綠素b的質量分數(圖3),使其在低光條件下能捕獲更多的光能,促進光能的轉化和傳遞(圖4),進而提高凈光合速率和光能利用效率(圖5,圖6).
通常情況下,比葉質量較大的葉片也會具有較大的光合潛力.研究結果表明,VAL1-OE超表達水稻葉片比葉質量顯著高于野生型(圖2).這有利于使葉片在單位面積上獲得更多的氮素分配和光合蛋白的分布,進而提高VAL1-OE超表達水稻葉片在強光下的電子傳遞效率(圖4)、光合碳同化效率(圖5)和光能利用效率(圖6).研究結果還表明,在強光條件下,VAL1-OE超表達水稻葉片ETRI顯著高于ETRII(圖4),這主要是因為強光下水稻通過PS I的電子傳遞速率顯著增強,進而導致葉片環式電子傳遞顯著提高,而環式電子傳遞增強有利于高光強下實現對光合機構的保護作用[17].綜上可知,VAL1-OE超表達水稻葉片關鍵光合酶編碼基因轉錄水平的上調,顯著提高了光合捕光復合蛋白的質量分數,促使無論是低光還是高光條件下,VAL1-OE超表達水稻葉片較野生型均具有顯著的光合能力和光能利用效率.
3.2 在高光合基礎上培育高葉面積表型材料,進一步提高光合作用面積是高光效水稻育種的突破口
具有良好的株型特征是作物實現高光效的基礎.張耀文等[18]在研究作物的高光效試驗中認為,葉面積系數較大,株型緊湊且適于密植,莖稈粗壯抗倒伏能力強,冠層結構較良好,群體內光照分布合理、群體光合速率較高.本研究結果表明,在抽穗期時VAL1-OE超表達水稻與野生型干物質累積量之間差異無統計學意義(表1),但株高和分蘗數量顯著低于野生型(圖2).這可能是因為VAL1-OE超表達水稻葉片具有較強的光合能力,較多的干物質累積分配到各個分蘗當中,單分蘗獲得了更多的干物質分配,致使其干物質總量未出現較大差異.此外,在收獲期時,VAL1-OE超表達水稻與野生型實際收獲植株的分蘗數量差異無統計學意義,可能是由于進入抽穗期后,VAL1-OE超表達水稻重新獲得一些新的有效分蘗增加了最終的有效穗數.
提高作物光能物質生產能力的途徑有3個:提高光合能力、增大光合面積和延長光合作用的時間.在本研究中,雖然VAL1-OE超表達水稻具有顯著高的葉綠素質量分數、比葉質量和凈光合速率(圖2,圖3,圖5),但VAL1-OE超表達水稻單株面積顯著低于野生型(圖2).光合面積較小成為制約VAL1-OE超表達水稻獲得更多干物質累積和產量的主要因素.以VAL1-OE超表達水稻為基礎,在實現高光合能力的同時,培育高葉面積表型材料,提高光合作用面積是未來產量進一步提升的突破口.
4 結論
超表達VAL1-OE水稻優化葉片光能吸收、電子傳遞和碳同化是提高光合作用和光能利用效率的關鍵.VAL1-OE超表達水稻葉片關鍵光合酶編碼基因轉錄水平的上調,顯著提高了光合捕光復合蛋白的質量分數,促使無論是低光還是高光條件下,VAL1-OE超表達水稻葉片較野生型均具有顯著增強的電子傳遞速率和凈光合速率,致使其光能利用效率顯著提高.與此同時,光合面積較小成為制約VAL1-OE超表達水稻獲得更多干物質累積和產量的主要因素.以VAL1-OE超表達水稻為基礎,在實現高光合能力的同時,培育高葉面積表型材料,提高光合作用面積是進一步提高該水稻材料干物質累積量和產量的關鍵.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
軍事文摘(2021年22期)2021-11-26 00:43:51
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
