COPD患者血清microRNA表達譜及相關生物信息學分析*
2023-01-31 14:04:14胡雪茹劉軍輝申永春秦江月文富強
西部醫學 2023年1期
胡雪茹 劉軍輝 申永春 秦江月 文富強
(四川大學華西醫院呼吸與危重癥醫學科, 四川 成都 610041)
慢性阻塞性肺疾病(Chronic obstructive pulmonary disease,COPD)又稱慢阻肺,是一種常見的可預防和治療的慢性氣道疾病,特征是持續存在的氣流受限和相應的呼吸系統癥狀[1]。2021GOLD報道,全球范圍到2060年,每年可能有540萬以上的人死于慢阻肺和其相關疾病[2]。國內的流行病學研究顯示,我國 20歲及以上成人慢阻肺患病率為8.6%,40歲以上的人群將近1億人,其已成為我國乃至世界范圍內的重大慢性非傳染性疾病,造成了巨大的公共衛生負擔[3]。早期明確慢阻肺的診斷對后續的治療有重要的臨床意義,目前肺功能檢查仍是確診慢阻肺的必備條件,但當前基于肺功能的慢阻肺診斷在我國的臨床實踐中仍存在較大的挑戰[4],尋找到簡單易行的慢阻肺診斷標志物具有重要的臨床意義。microRNA(miRNA)是一組小的非編碼RNA(長度大約為22個核苷酸),通過mRNA 降解、抑制蛋白質翻譯或通過這兩種機制的組合,在轉錄后負調控基因表達,是細胞增殖和分化、發育和凋亡的多個生物途徑中的關鍵調節劑[5]。miRNA已成為包括神經膠質瘤、結直腸癌、肺癌、心力衰竭在內的多種疾病的生物標志物[6],在疾病的診斷、分期中具備廣闊的臨床運用前景。近年來研究發現,miRNA與慢阻肺的關系密切,有可能被用作潛在慢阻肺的生物標志物。目前研究發現miRNA可能主要通過調控MAPK、Wnt、NF-κB、JAK-STAT等信號通路參與慢阻肺的發病機制[7],但其在慢阻肺診斷和致病的詳細機制中尚未完全被闡釋。因此本研究旨在探索慢阻肺患者血清miRNA差異性表達譜,并通過生物信息學預測差異性miRNA的靶基因并進一步分析其靶基因在調控生物學過程功能分析、信號通路、蛋白質互作中的機制,探索miRNA對慢阻肺的的臨床診斷意義及潛在相關的分子生物學機制。
1 資料與方法
1.1 數據的提取及分析 GSE70080的miRNA表達譜數據集從GEO(http://www.ncbi.nlm.nih.gov/geo/)下載[8], GSE70080位于平臺GPL20591中,其中包括16個慢阻肺患者和16個正常血清樣本。從GSE70080數據集下載微陣列數據的歸一化表達矩陣。然后用數據集中的注釋文件對探針進行注釋。R軟件的“limma”包用于識別差異表達的miRNA。P<0.05基因被認為是差異表達的基因。受試者曲線及火山圖、熱圖聚類圖使用R軟件的“ggplot2”包進行。受試者工作特征(Receiver operating characteristic, ROC)曲線法評估差異性表達miRNA對慢阻肺的診斷價值。
1.2 靶基因的預測 運用靶基因預測軟件TargetScan、PicTar2和 miRanda分別預測差異miRNA的靶基因[9],再分別取各自在3個軟件下預測所得靶基因的交集,然后利用miTarbase數據庫獲取經過至少3種實驗驗證并且靶向關系類型為具有功能的miRNA-靶基因相互作的靶基因,將軟件預測所得結果與靶向關系預測結果取并集作為所納入每個miRNA的靶基因集合,繪制miRNA-mRNA網絡圖。
1.3 靶基因生物信息分析 基因本體(Gene Ontology, GO)分析和京都基因和基因組百科全書(Kyoto Encyclopedia of Genes and Genomes, KEGG)信號通路富集分析在R軟件中使用“GO plot”包進行。GO分析包括細胞成分(CC)、生物過程(BP)和分子功能(MF)[10]。
1.4 miRNA靶基因的蛋白互作分析 使用STRING數據庫(https://string-db.org/)和Cytoscape軟件對差異性表達miRNA的靶基因進行蛋白互作網絡(Protein protein interaction network, PPI network)分析[11]。
2 結果
2.1 差異性miRNA表達分析 對于慢阻肺組和對照組的高通量測序結果,見圖1。根據P<0.05篩選出8個上調和54個下調差異miRNA,與慢阻肺患者相比,miR-655和miR-337在慢阻肺樣本中的表達增加了近兩倍,miR-645在慢阻肺組織中的表達平均降低了4倍,見圖2。
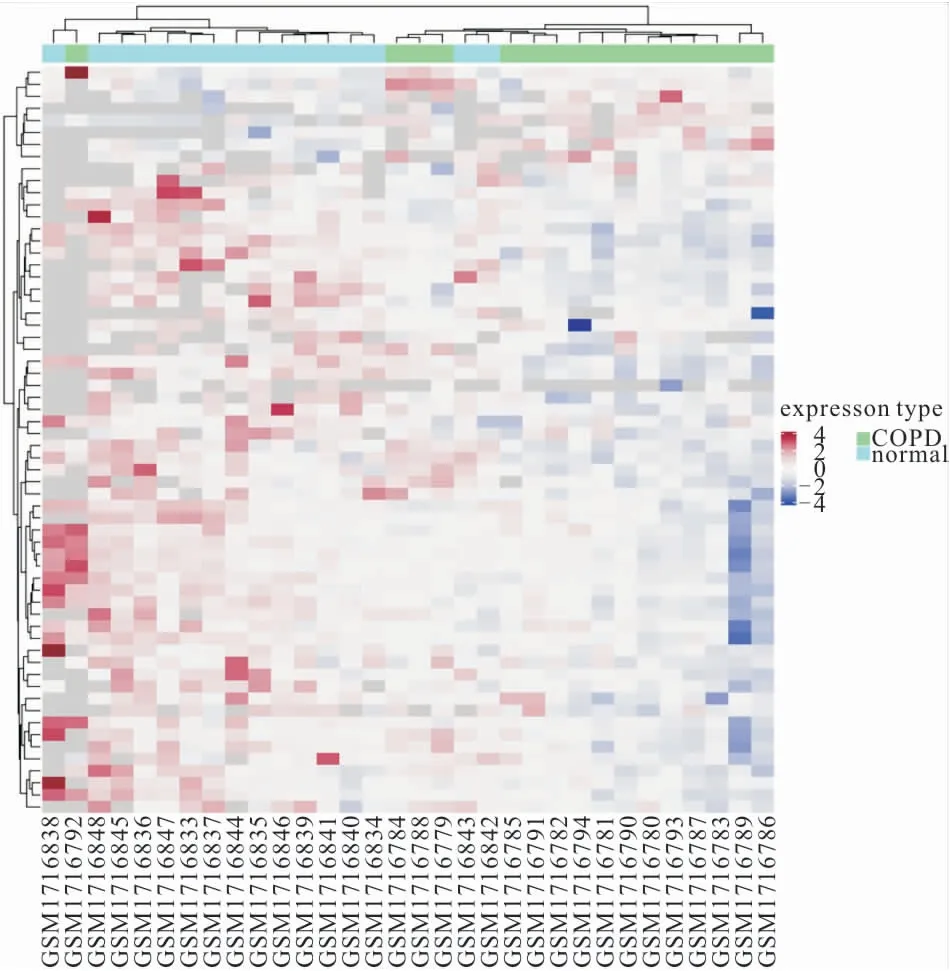
圖1 慢阻肺患者與健康對照者血清miRNA差異性表達熱圖

圖2 慢阻肺患者與健康對照者血清miRNA差異性表達火山圖
2.2 差異性表達miRNA對慢阻肺的診斷價值 ROC曲線分析顯示下調的54個miRNA診斷慢阻肺的ROC曲線下面積為0.969,上調的8個miRNA診斷慢阻肺ROC曲線下面積是0.938,見圖3。
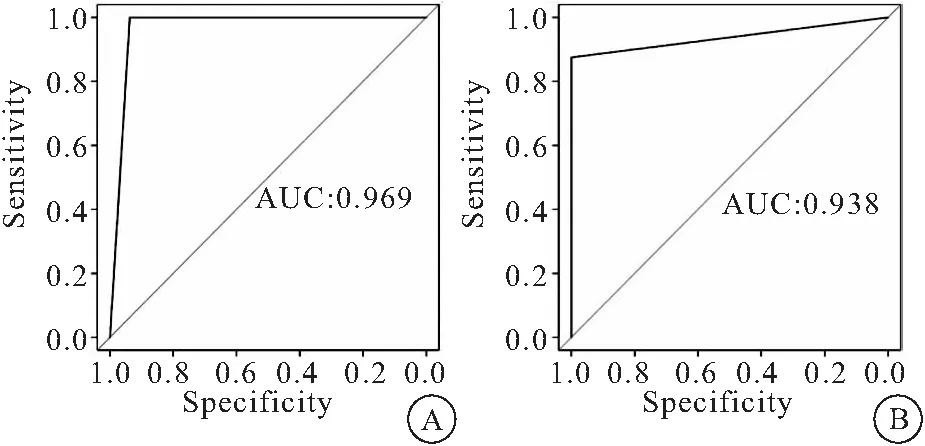
圖3 差異性表達miRNA診斷慢阻肺的價值
2.3 靶基因的預測與生物富集分析 通過3個靶基因預測軟件TargetScan、PicTar2和 miRanda預測靶基因,結合miTarbase數據庫中經過驗證,共15099個靶基因。GO富集結果見表1,其中下調差異miRNA的靶基因GO富集的主要生物過程涉及Wnt、MAPK、NF-KB信號通路;上調miRNA的靶基因集合生物過程主要涉及Wnt信號通路;在KEGG富集分析中,下調差異miRNA富集于MAPK信號通路;上調差異miRNA富集于Wnt、MAPK信號通路。見圖4。
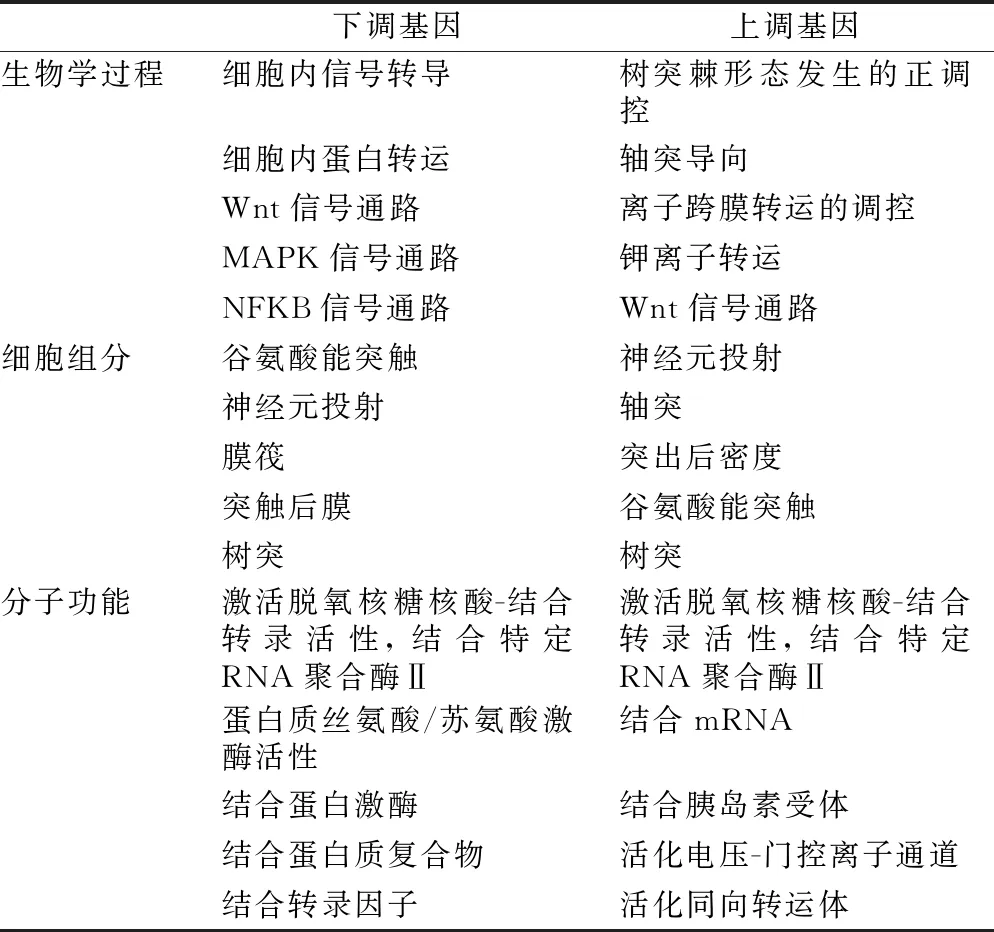
表1 差異性表達miRNA靶基因GO分析結果
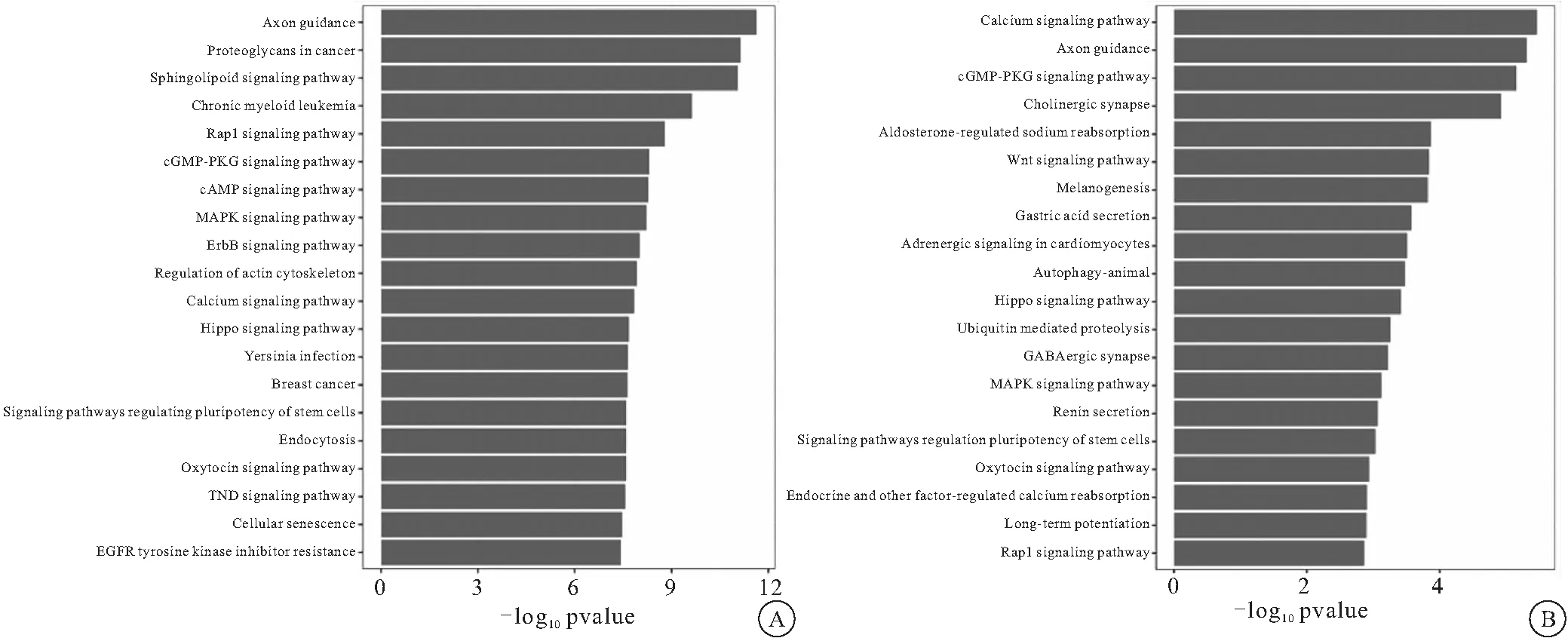
圖4 差異性表達miRNA靶基因KEGG分析結果
2.4 miRNA與靶基因網絡分析 經過實驗驗證并且靶向關系類型為具有功能的miRNA-mRNA網絡圖,見圖5。其中13種miRNA(hsa-miR-140-3p、hsa-miR-206、hsa-miR-337-5p、hsa-miR-574-3p 、hsa-miR-645、hsa-miR-769-5p、hsa-miR-107、hsa-miR-324-5p、hsa-miR-532-3p、hsa-miR-628-3p、hsa-miR-375 和hsa-miR-543),其中12種下調,1種 miRNA(hsa-miR-337-5p)在上調。4個核心miRNA為:miR-574-3p、hsa-miR-628-3p、hsa-miR-107、hsa-miR-769-5p;多個miRNA與其靶基因相互作用,其中SLC4A8受7個miRNA的調節。

圖5 miRNA與靶基因網絡分析圖
2.5 miRNA靶基因PPI分析 將miRNA的靶基因合并,導入STRING數據庫,選擇高可信度的互作關系,刪除沒有互作關系的靶基因點后蛋白互作網絡,見圖6。根據該網絡結構中結點的連通性即按Degree對靶基因進行排序,其中前5位分別是ACVR1B、ACVRL1、KRT2、KRT74、KRT82。
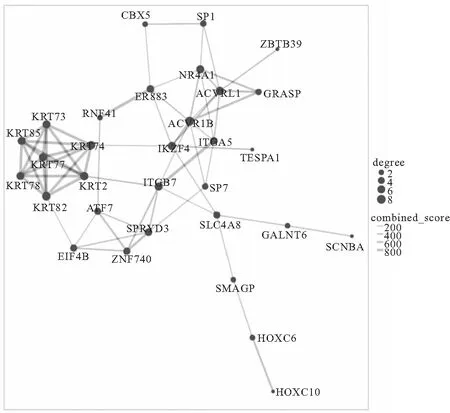
圖6 miRNA靶基因編碼蛋白質互作網絡分析圖
3 討論
慢阻肺的早期診斷與發病機制的探索對其患者的診治具有重要的臨床意義。本研究通過現代生物信息學分析方法研究慢阻肺患者血清樣本中的差異性miRNA,尋找潛在的miRNA生物標志物,并通過生物信息分析基于miRNA的慢阻肺發病機制。
慢阻肺差異miRNA的研究在多種樣本中得到證實,包括血清、肺組織、痰液、肺泡灌洗液,在臨床研究過程中發現異常表達的miRNA與體外研究中的表達趨勢一致,并且觀察到失調的miRNA與慢阻肺臨床嚴重程度相關,并且會增加穩定的慢阻肺患者急性加重的風險。本次研究發現慢阻肺組和正常對照組血清中miRNA表達存在差異,上調基因有8個,下調基因54個,根據差異miRNA繪制ROC曲線,曲線下面積均大于0.9(P<0.05),提示miRNA對慢阻肺有一定的診斷價值。其中血清中miR-320表達增加,這與以往研究一致,研究報道miR-320表達增加與生存率正相關,可作為慢阻肺的預后標志物[12]。有研究發現暴露于煙霧主要影響miRNA的下調[13],在本研究中54個基因表達下調,其中miR-106b被報道可作為反映慢阻肺患者持續或者全身變化的指標[14]。miR-23a、miR-25、miR-145 和 miR-224,與慢阻肺全球倡議 (GOLD) 階段相關。miR-23a和miR-145在慢阻肺的非頻繁和頻繁加重者之間有顯著差異,miR-23a可能是區分頻繁加重和非頻繁加重的潛在且有希望的生物標志物[15]。6個差異miRNA(let-7c、miR-125b、miR-324、miR-642、miR-340、miR-532)表達下調,與誘導痰中的表達一致,其中let-7c降低顯著。研究發現let-7c的主要靶點是腫瘤壞死因子受體Ⅱ(TNFR-Ⅱ),且痰中let-7c水平與一秒用力呼氣容積(FEV1)呈正相關, 與痰腫瘤壞死因子受體Ⅱ(sTNFR-Ⅱ)的蛋白水平呈顯著負相關[16],在慢阻肺小鼠模型中進一步證明,TNFR-Ⅱ基因敲除小鼠受到保護,免受香煙煙霧引起的炎癥和肺氣腫[17]。通過文獻學習,本研究發現下調的miRNA-206在肺組織中表達上調,且與重度慢阻肺患者的循環炎癥細胞因子有關[18],如TNF-α、IL-2和IL-5,可通過調節人肺成纖維細胞MRC-5細胞中的IRAK1促進脂多糖誘導炎癥損傷[19],其靶基因Notch3和VEGFA被證明在慢阻肺中可增強誘導細胞凋亡作用[20]。因此miRNA在不同樣本之間可能存在表達趨勢的差異,還需要進一步探索相關miRNA在慢阻肺中的確切作用。
有研究報道miRNA具有高穩定性、強特異性、高靈敏度和易于在血液中檢測,因此可用于臨床診斷及預后評估[21]。如miR-3620-3p可以區分慢阻肺和哮喘[22]。Leidinger等[23]表明特定的外周miRNA譜可以區分肺癌和慢阻肺。miR-146a的表達與慢阻肺急性加重期的嚴重程度呈負相關;miR-218-5p 與嚴重慢阻肺有相關性,但與吸煙狀況無關[24]。miR-146a 和 miR-146b 可以預測慢阻肺患者慢阻肺急性加重期的風險[25]。miR-1273-3p、miR-126、miR-503、miR-34b/c、miR-342-3p、miR-30e-3p、miR-125a-5p與 FEV1% 呈正相關,miR-106b-5p、miR-15b、miR-195、miR-24-3p、miR-320a、miR-320b、miR-34a 和 miR-199a-5p 的水平與 FEV1 %負相關[26]。綜上,慢阻肺患者血清中存在顯著的miRNA差異性表達,并且對慢阻肺具有一定的診斷及預后價值。鑒于其樣本可及性,患者配合程度高,miRNA有望成為慢阻肺的診斷標志物,并且有助于慢阻肺的嚴重程度評估。
miRNA一般通過其對靶基因的調控發揮其調節作用,本研究對差異性表達miRNA的靶基因進行GO注釋和KEGG pathway分析,其結果示差異miRNA的靶基因主要參與NF-kB、Wnt、MAPK信號通路。Wnt 信號通路包含控制各種發育和生理過程的信號分子家族,研究表明,Wnt/β-連環蛋白信號傳導參與肺發育、體內平衡、肺上皮損傷和修復過程[27]。慢阻肺的特點是由氣道重塑和慢性支氣管炎組成的小氣道疾病和導致肺氣腫的肺實質破壞引起的慢性氣流受限。研究顯示Wnt/β-catenin 信號在慢阻肺和肺氣腫患者及其動物模型中降低,通過Wnt/β-catenin 信號的激活可減少空腔擴大和膠原含量來減輕慢阻肺的癥狀[28]。典型Wnt/β-連環蛋白激活還可導致肺上皮細胞增殖增加,表明激活Wnt信號傳導可引起上皮細胞的修復機制[29]。本研究發現miR-130a上調,與之前研究表達一致,研究發現抑制miR-130a顯著減輕香煙誘導的對細胞遷移和增殖的抑制,miR-130a 過表達可阻斷香煙刺激引起Wnt 信號的激活,抑制 miR-130a 可通過 Wnt/β-Catenin 信號通路減輕香煙煙霧引起的肺損傷[30]。miR-23a、miR-25、miR-145 和 miR-224也被證明參與了Wnt信號通路[15]。目前研究發現Wnt信號通路以不同方式參與慢阻肺中的慢性氣道炎癥、氣道重塑、肺氣腫的發生發展[31],但仍需要進一步研究以確定miRNA如何調控Wnt信號通路在慢阻肺中的調節機制。
NF-κB作為一種核轉錄因子, 與免疫應答、炎癥反應及細胞的增生、轉化、凋亡等病理生理過程密切相關。NF-κB的活化是慢阻肺發病的重要炎癥通路之一, 通過調控TNF-α、IL-6、IL-8、IL-12、COX-2、粘附分子、趨化因子、集落刺激因子等參與了炎癥反應各階段和早期的免疫反應[32]。在嚙齒動物中有研究報道miR-30、miR-146、miR-132 和 miR-155參與 NF-κB 通路激活[33],進一步研究證實在慢阻肺患者的氣道或樣本中NF-κB通路活性標志物的增加,包括慢阻肺惡化期間的痰巨噬細胞以及在穩定期慢阻肺患者的支氣管活檢中[34]。如miR-218通過腫瘤壞死因子受體I(TNFR1)介導的NF-κB激活起作用,參與炎癥和粘蛋白(MUC5AC)過度產生,而顯著下調的 MiR-218被認為是慢阻肺的保護因素[24]。香煙煙霧誘導慢阻肺小鼠氣道上皮和肺巨噬細胞中miR-21表達上調,通過miR-21/SATB1/S100A9/ NF-κB軸在慢阻肺中發揮致病作用,遂miR-21可作為慢阻肺的潛在治療靶點[35]。因此,抑制NF-κB信號通路的活化,可能減少氣道細胞中炎癥基因的表達,NF-κB抑制劑或調節劑可能具有干預及治療慢阻肺的潛力。參與NF-κB通路的miRNA可能成為慢阻肺治療新靶點。
本文通過miRNA靶基因的預測,進一步構建了潛在的miRNA-mRNA調控網絡。其中4個miRNA(miR-574-3p、hsa-miR-628-3p、hsa-miR-107、hsa-miR-769-5p)為核心基因。之前研究報道miR-628-3p在慢阻肺和有無吸煙患者中表達下調,miR-574-5p在香煙煙霧(0.5%)處理3天的人支氣管上皮細胞,較對照組表達下調[36],這均與本研究結果一致,提示吸煙可能影響慢阻肺患者miRNA的表達。miR-107、miR-769-5p的失調與各種腫瘤發展有關,尚未有研究探索miR-107、miR-769-5p與慢阻肺之間的關系。本研究中SLC4A8受7種miRNA調控,可能成為慢阻肺的核心基因。預測SLC4A8存在多個miRNA結合位點,后續的的研究需進一步探索其在慢阻肺中的作用機制。通過PPI分析,ACVR1B、ACVRL1、KRT2、KRT74、KRT82、NR4A1可能是參與慢性阻塞性肺疾病發生發展的核心基因。其中ACVR1B是TGF-β配體家族的成員,可控制許多細胞過程,研究顯示其表達水平與肺氣腫的分布具有顯著關聯[37]。ACVRL1是一種參與血管生成的內皮轉化生長因子β受體[38],在肺動脈高壓中發現ACVRL1突變,會導致 TGF-β/(BMP)信號受損,改變的BMP受體信號影響炎癥及其消退、DNA損傷及其修復以及細胞代謝的調節[39]。NR4A1參與FSTL-1(卵泡抑素樣1)缺陷型肺氣腫的形成[40],也可通過NF-κB信號傳導,進而以負反饋方式下調炎癥因子,減輕氣道炎癥[41]。KRT2、KRT74、KRT82編碼不同角蛋白,其與角質形成細胞活化、增殖和角質化相關,但目前在慢阻肺中的報道有限,還需要進一步研究。綜上所述,繞著這些基因深入研究,有望進一步闡明慢阻肺的發病機制,為慢阻肺的干預尋找到新的靶點。
研究已發現miRNA促進肺部發育、成熟,且在維持肺功能中發揮重要作用[42]。miRNA可從多個層面、多個機制參與慢阻肺的發病機制。慢阻肺一個重要發病機制是由慢性炎癥引起的支氣管上皮細胞的凋亡和增殖變化。多種miRNA被發現參與氣道炎癥,如miR-24-3p, miR-93-5p, miR-320a/b, miR-1273-3p,其靶基因主要是促炎基因,富集于NOD樣受體 ( NLR ) 和 Toll樣受體(TLR)通路[43]。此外還有miR-233被證實可控制肺細胞中HDAC2的表達和活性,從而調節慢阻肺的肺部炎癥[44]。Izzotti等[45]的研究分析了暴露于香煙煙霧環境的大鼠肺中126個 miRNAs下調,其主要調節應激反應、細胞凋亡、增殖、血管生成和基因表達,提示miRNA可能與慢阻肺的損傷與修復有關。Conickx等[24]的研究發現miR-218-5p的表達與氣道阻塞密切相關,在沒有氣流受限的吸煙者的肺中,miR-218-5p在戒煙至少1年后表現出正常的表達水平,無論是否吸煙,慢阻肺患者 miR-218-5p的不可逆下調可能導致持續的全身和肺部炎癥。miR-195在慢阻肺患者中表達增加,進一步研究發現抑制miR-195 可減輕吸煙誘導的肺損傷并阻斷體內炎癥過程,可能為慢阻肺的治療提供新靶點[46]。
免疫失調是慢阻肺的另一個重要發病機制。Shi等[47]證明miR-203通過靶基因TAK1和 PIK3CA發揮免疫反應抑制劑的作用,miR-203可能通過抑制吸煙者的免疫反應,促進慢阻肺的發生發展。據報道,慢阻肺患者的調節性T細胞中的 miRNA表達譜顯著改變。其中miR-199a-5p對調節性T細胞具有特異性,miR-199a-5p可通過影響Th1-Th17 平衡從而導致慢阻肺的發生[48]。自噬和細胞凋亡也在慢性阻塞性中發揮重要作用。在人支氣管上皮細胞中miR-21參與香煙煙霧誘導的自噬和細胞凋亡[49]。過表達miR-34a可顯著增加人肺微血管內皮細胞的凋亡率。Notch1的過表達對由miR-34a升高引起的人肺微血管內皮細胞凋亡具有保護作用[50]。差異表達的miRNA對慢阻肺也有保護作用。如miRNA-212-5p在體外細胞通過促進細胞增殖和抑制香煙煙霧誘導的慢阻肺相關基因和蛋白質的表達對慢阻肺發揮保護作用[51]。miR-483通過促進細胞生長和激活α-SMA和纖連蛋白發揮保護作用[52]。以上研究表明,miRNA從多個方面參與了慢阻肺的發病機制,未來需更多的研究明確與發病機制相關的miRNA和其關鍵通路,為慢阻肺探索新生物標志物。
4 結論
本研究結果發現慢阻肺患者血清中存在差異miRNA,其對于慢阻肺的診斷有一定幫助。經過靶基因的預測及和相關通路分析,提示miRNA與慢阻肺發病機制密切相關,也為后續的研究提供了生物信息學的基礎。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年21期)2018-11-09 01:23:06
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國衛生(2015年9期)2015-11-10 03:11:12
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
