UBE2L3參與肺腺癌細胞有氧糖酵解的機制研究
2023-02-19 07:01:26潘歡楊帆戚維波馬興杰
浙江醫(yī)學 2023年1期
關鍵詞:水平
潘歡 楊帆 戚維波 馬興杰
肺癌是我國最常見的惡性腫瘤,其發(fā)病率和死亡率均位居惡性腫瘤的第一位[1]。非小細胞肺癌(nonsmall cell lung cancer,NSCLC)是腫瘤最常見的組織學類型,約占肺癌的80%~85%[2]。其中,肺腺癌又是NSCLC中最常見的病理組織亞型。由于肺腺癌具有高侵襲性、高早期轉(zhuǎn)移率和高術后復發(fā)率等特點,所以患者的總體預后仍然不容樂觀[3]。研究表明腫瘤的發(fā)生和發(fā)展是一個復雜的多步驟過程,包括抑癌基因失活、癌基因的激活、腫瘤細胞的增殖和凋亡失衡,最終導致腫瘤細胞的惡性增殖。即使在氧氣充足的條件下,腫瘤細胞仍然傾向于將糖酵解作為其主要的能量代謝方式,這種現(xiàn)象被稱為Warburg效應[4]。深入研究有氧糖酵解過程中的分子調(diào)控機制有助于發(fā)現(xiàn)新的腫瘤治療靶點。本研究旨在探索泛素蛋白連接酶L3(ubiquitin conjugating enzyme E2 L3,UBE2L3)對肺腺癌細胞有氧糖酵解的影響,并初步研究UBE2L3調(diào)控有氧糖酵解的分子機制。
1 材料和方法
1.1 生物信息學分析 本研究生物信息學分析數(shù)據(jù)下載自腫瘤基因組圖譜(the Cancer Genome Atlas,TCGA)數(shù)據(jù)庫(https://www.cancer.gov/tcga)。504例肺腺癌組織和59例正常肺組織列入本研究,比較肺腺癌組織和正常肺組織中UBE2L3蛋白表達水平。根據(jù)UBE2L3在肺腺癌組織中的表達中位值將肺腺癌組織分為UBE2L3高表達組和低表達組,運用R語言中的“survival”和“survminer”生存分析包分析UBE2L3 mRNA表達水平與肺腺癌患者總生存期的關系。根據(jù)肺腺癌組織中UBE2L3 mRNA表達水平的中位值將肺腺癌患者分為高表達組316例和低表達組188例,分析UBE2L3 mRNA表達水平對肺腺癌組織中有氧糖酵解水平的影響,即運用Spearman秩相關分析UBE2L3 mRNA表達水平與LDHA和PKM2 mRNA表達水平的相關性。
1.2 實驗細胞、試劑和儀器 人肺腺癌細胞株HCC827和H1299均購自中國科學院上海細胞庫。FBS購自美國Hyclone公司;RPMI 1640培養(yǎng)基購自美國Corning公司;UBE2L3過表達質(zhì)粒、小干擾RNA(small interfering RNA,siRNA)質(zhì)粒及陰性對照質(zhì)粒均購自上海吉瑪公司;胰酶、支原體檢測試劑盒、苯甲基磺酰氟(phenylmethanesulfonylfluoride or phenylmethylsulfonyl fluorid,PMSF)蛋白酶與磷酸酶抑制劑、細胞裂解液RIPA、BCA蛋白定量試劑盒、硝酸纖維素膜、脫脂奶粉、辣根過氧化物酶標記羊抗人的IgG、ECL化學發(fā)光顯影液、磷脂酰肌醇-3-激酶(phosphatidylinositol-3-hydroxykinase,PI3K)/蛋白激酶 B(protein kinase B,Akt)抑制劑Y294002、二甲基亞砜(dimethyl sulfoxide,DMSO)均購自中國上海碧云天生物技術有限公司;Lipofectamine 3000轉(zhuǎn)染試劑購自美國Thermo Fisher公司;CCK-8試劑盒購自日本Dojindo公司;葡萄糖測定試劑盒、乳酸檢測試劑盒均購自美國Sigma公司;UBE2L3抗體、M2-型丙酮酸激酶(pyruvate kinase M2,PKM2)抗體、L-乳酸脫氫酶(lactate dehydrogenase A,LDHA)抗體、Akt抗體、磷酸化 Akt(phosphorylated protein kinase B,p-Akt)抗體、甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗體均購自美國Abcam公司。倒置熒光相差顯微鏡(型號:CKX53)購自日本Olympus公司;垂直電泳儀(Power Pac Basic)、蛋白轉(zhuǎn)印模塊(Mini TransBlot Cell)和凝膠成像系統(tǒng)(型號:BIORAD Gel Doc XR+)均購自美國Bio-Rad公司;CO2細胞培養(yǎng)箱、低溫離心機(型號:Thermo Scientific MicroCL 21R)均購自美國Thermo Fisher公司;酶標儀(型號:ELX-800)購自美國Bio-Tek公司。
1.3 細胞培養(yǎng) 人肺腺癌細胞株HCC827和H1299均使用含10%FBS的RPMI 1640培養(yǎng)基培養(yǎng)于37℃、5%CO2培養(yǎng)箱中,隔天更換新鮮培養(yǎng)基。細胞定期檢測有無支原體污染,當細胞融合度達到80%~90%時用胰酶消化傳代。取對數(shù)生長期的細胞用于實驗。
1.4 細胞轉(zhuǎn)染 將實驗分為兩個部分。(1)UBE2L3過表達細胞:選擇UBE2L3表達水平較低的HCC827細胞為研究對象,將對數(shù)生長期的細胞均勻鋪于6孔板中,待細胞融合度達到60%~70%時采用Lipofectamine 3000轉(zhuǎn)染試劑,按說明書進行轉(zhuǎn)染操作,分別將UBE2L3過表達質(zhì)粒以及陰性對照質(zhì)粒轉(zhuǎn)染到細胞中,轉(zhuǎn)染48 h后,熒光顯微鏡下觀察細胞的轉(zhuǎn)染效率,構建UBE2L3過表達質(zhì)粒細胞HCC827/UBE2L3及陰性對照細胞HCC827/NC,收集細胞用于后續(xù)實驗。(2)UBE2L3敲低細胞:選擇UBE2L3表達水平較高的H1299細胞為研究對象,如上述方法鋪板,再分別將siRNA質(zhì)粒以及陰性對照質(zhì)粒轉(zhuǎn)染到細胞中,轉(zhuǎn)染48 h后,熒光顯微鏡下觀察細胞的轉(zhuǎn)染效率,構建UBE2L3敲低細胞H1299/si-UBE2L3及陰性對照細胞H1299/si-NC,收集細胞用于后續(xù)實驗。
1.5 細胞增殖能力檢測 采用CCK-8法。將上述轉(zhuǎn)染成功的細胞接種于96孔板中,每孔加入100 μl細胞懸液(含3×103個細胞)。每組設置5個復孔,分別于0、24、48、72、96 h加入10 μl CCK-8溶液,于培養(yǎng)箱中37℃孵育4 h。采用酶標儀測定450 nm波長處的吸光度(optical density,OD)值,并繪制細胞生長曲線。
1.6 細胞中葡萄糖消耗量檢測 采用葡萄糖測定試劑盒。嚴格按照試劑盒操作說明書配置工作液。將上述轉(zhuǎn)染成功的細胞和工作液加入96孔板中,并設置3個復孔。置于37℃培養(yǎng)箱中孵育30 min。取出96孔板,去除氣泡后置于酶標儀上檢測630 nm波長處的OD值,繪制葡萄糖消耗量曲線,計算細胞中葡萄糖消耗量。
1.7 細胞中乳酸生成量檢測 采用乳酸檢測試劑盒。收集上述轉(zhuǎn)染成功的細胞培養(yǎng)基上清液2 ml,加入1 ml酶工作液,置于37℃培養(yǎng)箱中孵育10 min,隨后加入終止液22 ml終止酶促反應。將上清混合液加入96孔板中,置于37℃培養(yǎng)箱中培養(yǎng)30 min。取出96孔板,使用酶標儀測定490 nm波長處的OD值,計算細胞中乳酸生成量。
1.8 UBE2L3、有氧糖酵解相關蛋白PKM2、LDHA表達水平檢測 采用Western blot法。使用含1%PMSF蛋白酶抑制劑的細胞裂解液RIPA裂解上述轉(zhuǎn)染成功的細胞。置于冰上均勻搖晃15 min后,采用細胞刮子收集裂解液。裂解液于4℃、12 000 g離心15 min,收集上清液采用BCA法測定蛋白濃度。加入蛋白緩沖液調(diào)節(jié)蛋白濃度,于100℃沸水中水浴5 min使其蛋白變性,將蛋白裂解液置于-80℃冰箱備用。每孔加入30 μg蛋白樣品,經(jīng)10%聚丙烯酰胺凝膠電泳并轉(zhuǎn)移至硝酸纖維素膜上。將整張膜浸泡5%脫脂奶粉常溫封閉1 h,加入UBE2L3(1∶1 000)抗體、PKM2(1∶1 000)抗體、LDHA(1∶1 000)抗體、Akt(1:1 000)抗體、p-Akt抗體和GAPDH(1∶2 000)抗體于4℃下?lián)u床過夜孵育。PBST緩沖液洗膜后加入辣根過氧化物酶標記的二抗,常溫孵育1 h。采用化學發(fā)光顯影液發(fā)光,在成像系統(tǒng)中掃描并分析結(jié)果。使用Image J軟件計算各個條帶的灰度值,即蛋白表達水平。
1.9 PI3K/Akt抑制劑對HCC827/UBE2L3細胞的有氧糖酵解的影響 將上述轉(zhuǎn)染成功后的HCC827/UBE2L3細胞,分別加入PI3K/Akt抑制劑Y294002(1.5 μmol/L)(HCC827/UBE2L3+LY)及DMSO(1.5 μmol/L)作為陰性對照(HCC827/UBE2L3+DMSO)。細胞培養(yǎng)24 h后,收集細胞,上述方法檢測細胞增殖能力、葡萄糖消耗量、乳酸生成量及UBE2L3、有氧糖酵解相關蛋白PKM2、LDHA表達水平。
1.10 統(tǒng)計學處理 采用SPSS 25.0統(tǒng)計軟件。計量資料組間比較采用兩獨立樣本t檢驗。P<0.05為差異有統(tǒng)計學意義。
2 結(jié)果
2.1 肺腺癌組織和正常肺組織中UBE2L3蛋白表達水平比較 生物信息學分析提示,肺腺癌組織中UBE2L3蛋白表達水平高于正常肺組織(P<0.05),見圖1。
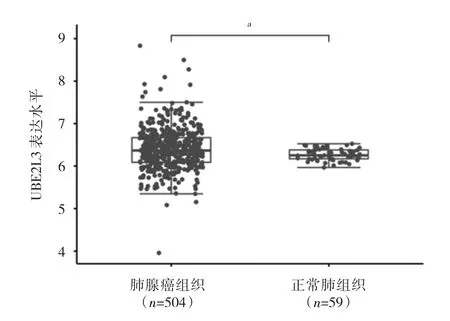
圖1 肺腺癌組織和正常肺組織中UBE2L3蛋白表達水平比較
2.2 不同UBE2L3表達水平肺腺癌患者總生存期的比較 UBE2L3高表達組患者總生存期低于低表達組(P<0.05),見圖2。
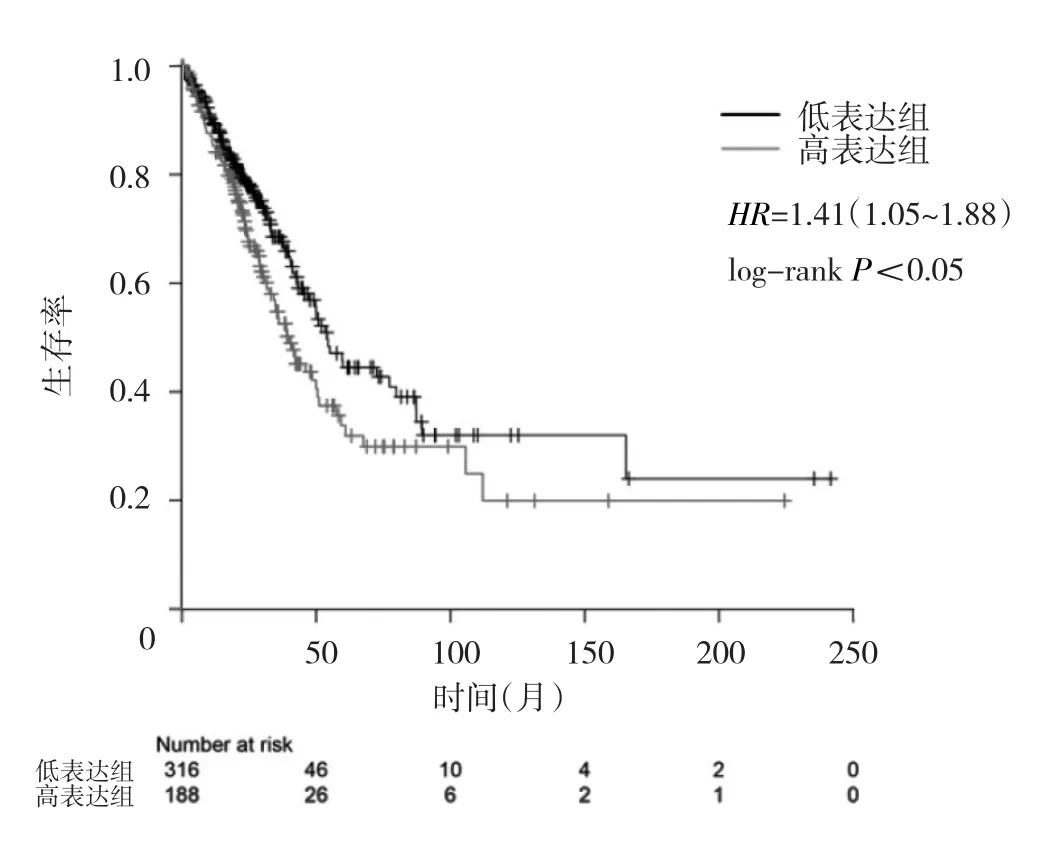
圖2 不同UBE2L3表達水平肺腺癌患者總生存期的比較
2.3 UBE2L3 mRNA表達水平對肺腺癌組織中有氧糖酵解水平的影響 肺腺癌組織中UBE2L3 mRNA表達水平升高,同時LDHA和PKM2 mRNA表達水平也隨著升高。UBE2L3 mRNA表達水平與LDHA和PKM2 mRNA表達水平均呈正相關(r=0.258和0.404,均P<0.01),見圖3。

圖3 UBE2L3 mRNA表達水平對肺腺癌組織中有氧糖酵解水平的影響(A:肺腺癌組織中UBE2L3 mRNA表達水平與LDHA mRNA表達水平相關性分析的散點圖;B:肺腺癌組織中UBE2L3 mRNA表達水平與PKM2 mRNA表達水平相關性分析的散點圖)
2.4 UBE2L3表達水平對肺腺癌細胞增殖能力的影響 HCC827/UBE2L3組細胞在48、72、96 h時增殖能力均高于HCC827/NC組(均P<0.05),見圖4A。H1299/si-NC組細胞在48、72、96 h時增殖能力均高于H1299/si-UBE2L3組(均P<0.05),見圖4B。

圖4 UBE2L3表達水平對肺腺癌細胞增殖能力的影響(A:HCC827/UBE2L3組和HCC827/NC組細胞不同時點增殖能力比較;B:H1299/si-UBE2L3組和H1299/si-NC組細胞不同時點增殖能力比較)
2.5 UBE2L3表達水平對肺腺癌細胞中葡萄糖消耗量與乳酸代謝的影響 有氧糖酵解實驗結(jié)果顯示,HCC827/UBE2L3組細胞葡萄糖消耗量大于HCC827/NC組,H1299/si-UBE2L3組細胞葡萄糖消耗量小于H1299/si-NC組,差異均有統(tǒng)計學意義(均P<0.05),見圖5A。HCC827/UBE2L3組細胞乳酸生成量大于HCC827/NC組,H1299/si-UBE2L3組細胞乳酸生成量小于H1299/si-NC組,差異均有統(tǒng)計學意義(均P<0.05),見圖5B。
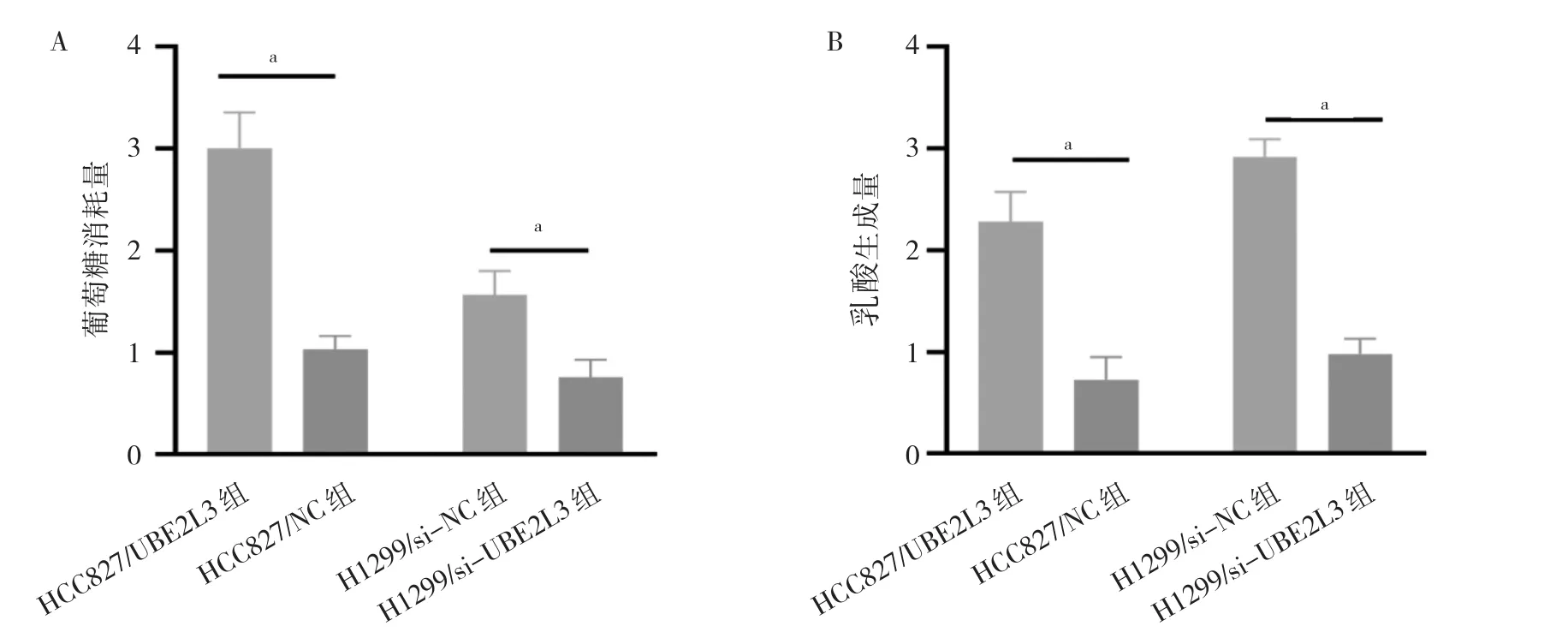
圖5 UBE2L3表達水平對肺腺癌細胞中葡萄糖消耗量與乳酸代謝的影響(A:不同組間葡萄糖消耗量比較;B:不同組間乳酸生成量比較)
2.6 UBE2L3表達水平對肺腺癌細胞中有氧糖酵解相關蛋白表達的影響 HCC827/UBE2L3組細胞PKM2和LDHA蛋白表達水平明顯高于HCC827/NC組,而H1299/si-UBE2L3組細胞PKM2、LDHA蛋白表達水平明顯低于H1299/si-NC組,見圖6。

圖6 UBE2L3 表達水平影響肺腺癌細胞有氧糖酵解相關蛋白表達的電泳圖
2.7 UBE2L3表達水平對PI3K/Akt信號通路蛋白表達的影響 過表達UBE2L3后,HCC827/UBE2L3組細胞p-Akt蛋白表達水平明顯高于HCC827/NC組,而兩組細胞Akt表達水平無明顯變化;H1299/si-UBE2L3組細胞p-Akt蛋白表達水平低于H1299/si-NC組,而兩組細胞Akt表達水平無明顯變化,見圖7。

圖7 UBE2L3表達水平影響PI3K/Akt信號通路相關蛋白表達的電泳圖
2.8 PI3K/Akt抑制劑對UBE2L3的有氧糖酵解的影響 經(jīng)過抑制劑Y294002處理HCC827/UBE2L3細胞后,Akt活性被抑制,p-Akt、LDHA和PKM2蛋白表達水平均下降,見圖8A。同樣,細胞增殖實驗結(jié)果提示,經(jīng)過抑制劑處理后的HCC827/UBE2L3細胞增殖能力較HCC827/UBE2L3+DMSO組下降,見8B。通過有氧糖酵解實驗可以看出,經(jīng)過抑制劑處理后的HCC827/UBE2L3細胞葡萄糖消耗量較HCC827/UBE2L3+DMSO組明顯下降,見8C。乳酸生成量檢測結(jié)果也可以看出,經(jīng)過抑制劑處理后的HCC827/UBE2L3細胞乳酸生成量較HCC827/UBE2L3+DMSO組明顯下降,見圖8D。

圖8 PI3K/Akt抑制劑對UBE2L3的有氧糖酵解的影響(A:HCC827/UBE2L3細胞中加入抑制劑后p-Akt、PKM2和LDHA蛋白表達的電泳圖;B:加入抑制劑后對HCC827/UBE2L3細胞增殖的影響;C:加入抑制劑后對HCC827/UBE2L3細胞葡萄糖消耗量的影響;D:加入抑制劑后對HCC827/UBE2L3細胞乳酸生成量的影響)
3 討論
在有氧條件下,正常細胞通過線粒體氧化磷酸化提供能量,而腫瘤細胞通過糖酵解途徑產(chǎn)生絕大部分維持生存和快速增殖的能量,該現(xiàn)象被稱為Warburg效應或有氧糖酵解[5]。癌細胞代謝重編輯被認為是惡性腫瘤的十大重要特征之一[6]。糖酵解使得腫瘤細胞在即使缺氧的條件下也能存活下來,并為腫瘤細胞提供快速增殖所需的能量。同時糖酵解過程中產(chǎn)生的酸性環(huán)境有利于腫瘤細胞的遷移和侵襲,增加腫瘤遠處轉(zhuǎn)移率[7-8]。因此腫瘤的有氧糖酵解過程是近年來腫瘤領域的研究熱點。盡管Warburg效應的發(fā)現(xiàn)已有很長一段時間,但其具體調(diào)控機制仍然尚不清楚。深入研究有利于探索新的腫瘤治療靶點,改善腫瘤患者的生存預后。
UBE2L3是泛素結(jié)合酶家族成員之一,越來越多的研究表明UBE2L3的異常表達參與了多種腫瘤的發(fā)展過程。在肝癌中,UBE2L3促進肝癌細胞的增殖和轉(zhuǎn)移[9-10]。在前列腺癌中,UBE2L3參與調(diào)控上皮-間充質(zhì)細胞轉(zhuǎn)化過程,并且發(fā)揮癌基因的作用[11]。在宮頸癌中,UBE2L3參與調(diào)控順鉑化療耐藥的分子機制[12]。之前研究報道UBE2L3在肺腺癌組織中異常高表達,且與不良臨床病理特征有關[13]。體內(nèi)外實驗均表明,UBE2L3能夠顯著增強肺腺癌細胞的增殖能力。UBE2L3通過SCF(Skp2)泛素連接酶復合物促進p27kip1的泛素化降解調(diào)控細胞周期進程[13]。然而,UBE2L3是否參與肺腺癌的有氧糖酵解過程尚不清楚。葡萄糖消耗量和乳酸生成量是最常用的檢測糖酵解活性的指標。同時糖酵解過程受到LDHA和PKM2等關鍵蛋白酶的限速調(diào)控。因此,檢測細胞糖酵解關鍵酶的表達量也能夠檢測糖酵解的活性。本研究中,在HCC827細胞中過表達UBE2L3能夠顯著增加細胞對葡萄糖的消耗和乳酸的產(chǎn)量,同時提高糖酵解相關蛋白PKM2和LDHA蛋白的表達水平。而在H1299細胞中UBE2L3沉默能夠抑制細胞對葡萄糖的消耗和乳酸的產(chǎn)量,降低PKM2和LDHA蛋白的表達量。因此,UBE2L3參與了肺腺癌有氧糖酵解過程。
腫瘤中通常存在PI3K/Akt信號通路異常活化,參與多種惡性生物學過程,尤其是激活的PI3K/Akt信號通路能夠增加腫瘤細胞對葡萄糖的攝取和糖酵解相關基因的表達,從而促進腫瘤細胞的糖酵解過程[14-16]。深入研究發(fā)現(xiàn),過表達UBE2L3顯著增加p-Akt蛋白的表達;敲低UBE2L3后p-Akt蛋白水平降低。為了進一步驗證UBE2L3通過激活PI3K/Akt信號通路參與有氧糖酵解過程,采用PI3K/Akt抑制劑LY294002阻斷PI3K/Akt信號通路。UBE2L3過表達細胞中加入PI3K/Akt信號通路抑制劑Y294002處理后,葡萄糖消耗量和乳酸生成量明顯減少,而PKM2和LDHA蛋白表達水平下降。并且Y294002處理后HCC827/UBE2L3細胞的增殖能力明顯受到抑制。因此,UBE2L3可能參與激活PI3K/Akt信號通路從而促進有氧糖酵解過程,以此增加肺腺癌的惡性增殖。
綜上所述,本研究證實UBE2L3能夠通過激活PI3K/Akt信號通路促進肺腺癌葡萄糖的攝取,增強糖酵解過程,促進肺腺癌惡性增殖,為深入研究肺腺癌的發(fā)病機制提供了新的線索。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
