基于ERK/mTOR信號通路探討六味地黃丸對氧化應(yīng)激狀態(tài)下成骨細(xì)胞自噬的影響
2023-02-25 03:06:54謝麗華郭瀾陳賽楠李生強(qiáng)陳娟葛繼榮
中國骨質(zhì)疏松雜志 2023年1期
謝麗華 郭瀾 陳賽楠 李生強(qiáng) 陳娟 葛繼榮*
1.福建省中醫(yī)藥科學(xué)院骨質(zhì)疏松證候基因組學(xué)研究室,福建 福州 350003
2.福建省中西醫(yī)結(jié)合防治骨質(zhì)疏松重點(diǎn)實(shí)驗(yàn)室(福建省中醫(yī)藥科學(xué)院、福建中醫(yī)藥大學(xué)附屬康復(fù)醫(yī)院),福建 福州 350003
3.福建中醫(yī)藥大學(xué),福建 福州 350122
絕經(jīng)后骨質(zhì)疏松癥(postmenopausal osteoporosis,PMOP)是絕經(jīng)后婦女雌激素水平下降引起的一種全身骨代謝性疾病,其特征為單位體積骨量減少,骨組織微結(jié)構(gòu)破壞,從而導(dǎo)致骨脆性增加,骨折風(fēng)險(xiǎn)升高[1]。腎陰虛證是絕經(jīng)后骨質(zhì)疏松癥臨床常見證型之一,六味地黃丸具有滋陰補(bǔ)腎的功效,常用于治療腎陰虛型絕經(jīng)后骨質(zhì)疏松癥,能夠改善臨床癥狀、提高骨密度[2-3]。本課題組持續(xù)關(guān)注六味地黃丸治療絕經(jīng)后骨質(zhì)疏松癥分子機(jī)制問題[4-7],前期研究結(jié)果顯示,六味地黃丸防治骨質(zhì)疏松的作用機(jī)制可能與自噬相關(guān)[8],但其作用的具體機(jī)制需進(jìn)一步探討。
自噬是一種高度保守的細(xì)胞內(nèi)物質(zhì)分解代謝、循環(huán)過程[9],在正常情況下,適宜水平的自噬可以促進(jìn)成骨細(xì)胞的增殖并減少其凋亡。研究證明,氧化應(yīng)激產(chǎn)生的活性氧(reactive oxygen species,ROS)能誘導(dǎo)自噬發(fā)生,自噬又可以降低ROS水平,調(diào)節(jié)細(xì)胞氧化應(yīng)激水平[10]。目前,對于自噬在PMOP方面相關(guān)通路的研究主要以mTOR為主要“中轉(zhuǎn)站”的信號通路為主[11]。ERK通路在機(jī)體自噬調(diào)節(jié)過程中也起到了重要作用[12],但是具體機(jī)制尚不明確。研究表明[13],自噬作為潛在靶點(diǎn)應(yīng)用于臨床骨病藥物的開發(fā)將是一大熱點(diǎn)。本研究通過H2O2誘導(dǎo)成骨細(xì)胞模擬細(xì)胞氧化應(yīng)激狀態(tài),觀察六味地黃丸含藥血清調(diào)控ERK/mTOR信號通路對氧化應(yīng)激狀態(tài)下成骨細(xì)胞自噬的影響,為六味地黃丸臨床治療PMOP提供新的理論依據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)動物
SPF級雄性SD大鼠20只,體質(zhì)量180~220 g,購自上海杰思捷實(shí)驗(yàn)動物有限公司,許可證號:SCXK(滬)2018-0004。于福建省中醫(yī)藥科學(xué)院動物房適應(yīng)性喂養(yǎng)一周后開始實(shí)驗(yàn)。動物倫理審查編號:FJATCM-IAEC2020021。
1.2 主要試劑
小鼠顱頂前骨細(xì)胞亞克隆MC3T3-E1細(xì)胞購自中國科學(xué)院細(xì)胞庫;六味地黃丸濃縮丸(同仁堂,國藥準(zhǔn)字:Z19993068);α-MEM培養(yǎng)基、胎牛血清、胰蛋白酶(Gibco公司);雷帕霉素Rap(HY-10219)、U0126(HY-12031A)、H2DCFDA(HY-D0940)、CCK-8試劑盒(HY-K0301)均購自MedChemExpress公司;ERK1/2(9101 S)、p-ERK1/2(4370 T)、mTOR(2983 T)、p-mTOR(5536 T)均購自Cell Signaling Technology公司;Anti-LC3B(Abcam,ab192890);Rabbit Anti-GAPDH(Bioss,bs-10900R);超敏ECL化學(xué)發(fā)光試劑盒(碧云天)。
1.3 方法
1.3.1細(xì)胞培養(yǎng):使用含10%胎牛血清,1%青霉素鏈霉素雙抗的α-MEM培養(yǎng)基復(fù)蘇MC3T3-E1細(xì)胞,置于37 ℃,5%的CO2環(huán)境的培養(yǎng)箱中培養(yǎng)。2 d換液一次,當(dāng)細(xì)胞生長融合度達(dá)到80%時(shí),用胰蛋白酶消化細(xì)胞進(jìn)行傳代,取對數(shù)生長期的細(xì)胞實(shí)驗(yàn)。
1.3.2含藥血清制備:將20只SD大鼠隨機(jī)分為空白組和六味地黃丸組,每組10只。參照文獻(xiàn)[14]將六味地黃丸配置成生藥濃度0.8 kg/L的水溶液,每天灌胃2次。空白組則灌等量蒸餾水。連續(xù)灌胃7 d,第7天首次灌藥2 h后腹主動脈取血,制備六味地黃丸(LWDH)含藥血清及空白血清。
1.3.3分組及干預(yù):根據(jù)課題組前期實(shí)驗(yàn)結(jié)果[8],采用0.10 mmol/L H2O2干預(yù)細(xì)胞24 h,建立體外氧化應(yīng)激細(xì)胞模型。分為正常對照組(Control):不做干預(yù);模型組(Model):0.10 mmol/L H2O2;空白對照組(Blank):20%空白血清+0.10 mmol/L H2O2;六味地黃丸組(LWDH):20%六味地黃丸含藥血清+0.10 mmol/L H2O2;雷帕霉素組(Rap):5 μmol/L Rap+0.10 mmol/L H2O2;U0126組(U0126):10 μmol/L U0126+0.10 mmol/L H2O2;雷帕霉素+六味地黃丸組(Rap+LWDH):5 μmol/L Rap+20%六味地黃丸含藥血清+0.10 mmol/L H2O2;U0126+六味地黃丸組(U0126+LWDH):10 μmol/L U0126+20%六味地黃丸含藥血清+0.10 mmol/L H2O2,所有組均干預(yù)24 h。
1.3.4細(xì)胞活性氧水平檢測:細(xì)胞接種于6孔板,分組干預(yù)后,采用熒光探針檢測胞內(nèi)ROS的含量。探針終濃度為10 μmol/L,置于培養(yǎng)箱中培養(yǎng)1 h后,PBS清洗,置于倒置熒光顯微鏡下觀察。
1.3.5蛋白印跡法:分組收集細(xì)胞,提取總蛋白,根據(jù)不同蛋白的分子量選擇6%(mTOR、p-mTOR)、10%(GAPDH、ERK1/2、p-ERK1/2)、15%(LC3B)SDS-PAGE凝膠上進(jìn)行電泳分離,轉(zhuǎn)膜。脫脂奶粉封閉4 h,磷酸化蛋白用5% BSA進(jìn)行封閉。加入對應(yīng)一抗4 ℃孵育過夜,TBST洗滌后,加入二抗Goat Anti-Rabbit IgG H&L antibody,室溫孵育1.5 h,清洗。使用特超敏ECL化學(xué)發(fā)光試劑盒曝光,用多色熒光/化學(xué)發(fā)光成像儀拍照。
1.4 統(tǒng)計(jì)學(xué)分析
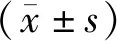
2 結(jié)果
2.1 六味地黃丸對成骨細(xì)胞ROS的影響
細(xì)胞活性氧水平顯示,與Control組相比,Model組ROS水平顯著提高(#P<0.05);與Model組相比,LWDH組、Rap組、U0126組、Rap+LWDH組及U0126+LWDH組細(xì)胞內(nèi)ROS水平皆下降(*P<0.05)。見圖1。
圖1 各組大鼠成骨細(xì)胞ROS情況Fig.1 ROS of osteoblasts in each group
2.2 六味地黃丸對成骨細(xì)胞自噬的影響
Western blot結(jié)果顯示,與Control組相比,Model組蛋白LC3Ⅱ/Ⅰ降低(#P<0.05);與Model組相比,LWDH組、Rap組、Rap+LWDH組蛋白LC3Ⅱ/Ⅰ增高(*P<0.05);與Model組相比,U0126組及LWDH+U0126組蛋白LC3Ⅱ/Ⅰ有增高趨勢,但差異無統(tǒng)計(jì)學(xué)意義(P>0.05)。見圖2。
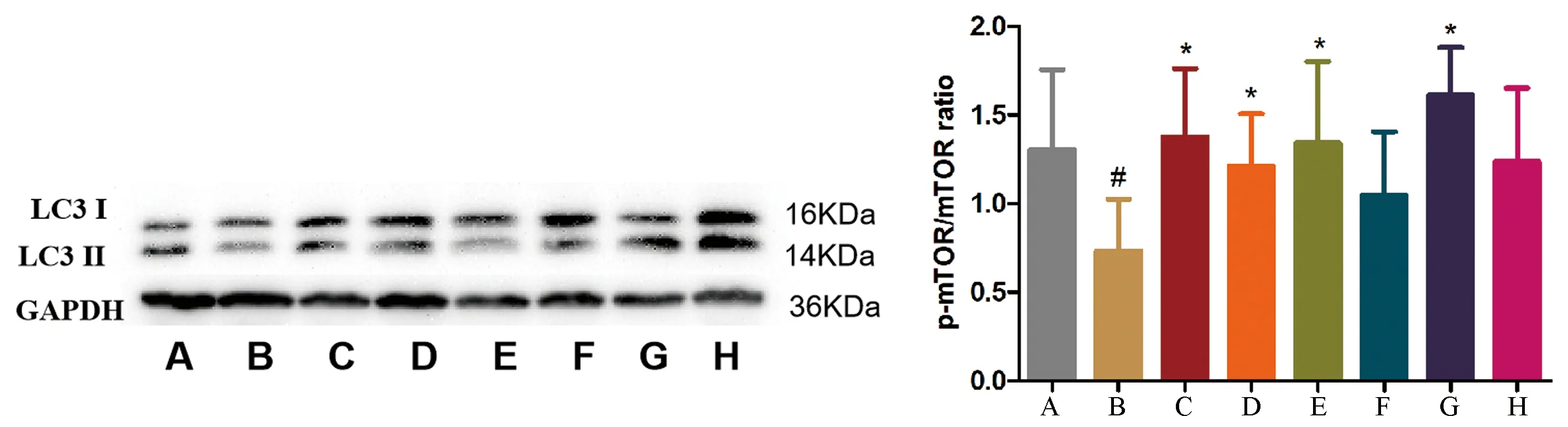
圖2 各組大鼠細(xì)胞LC3B蛋白表達(dá)情況Fig.2 Expression of LC3B protein in each group of cells注:A:Control組;B:Model組;C:LWDH組;D:Blank組;E:Rap組;F:U0126組;G:Rap+LWDH組;H:U0126+LWDH組。與Control組相比,#P<0.05;與Model組相比,*P<0.05。
2.3 六味地黃丸含藥血清對ERK/mTOR通路的影響
Western blot結(jié)果顯示,與Model組相比,LWDH組、Rap組與Rap+LWDH組的p-mTOR蛋白表達(dá)下調(diào)(*P<0.05)。與Model組相比,Blank組、LWDH組、U0126組的p-ERK1/2蛋白表達(dá)下調(diào)(*P<0.05);U0126+LWDH組p-ERK1/2蛋白表達(dá)有下調(diào)趨勢,但差異無統(tǒng)計(jì)學(xué)意義(P>0.05)。見圖3。

圖3 各組間p-mTOR、p-ERK蛋白的表達(dá)情況Fig.3 Expression of p-mTOR and p-ERK proteins among the groups注:A:Control組;B:Model組,C:Blank組;D:LWDH組;E:Rap組;F:U0126組;G:Rap+LWDH組;H:U0126+LWDH組。與Model組相比,*P<0.05。
3 討論
氧化應(yīng)激是機(jī)體的氧化和抗氧化系統(tǒng)之間的穩(wěn)態(tài)被破壞而造成的應(yīng)激狀態(tài)。當(dāng)人體因?yàn)樗ダ稀⒓膊〉仍虍a(chǎn)生ROS過多,而機(jī)體消除ROS能力不足時(shí),便產(chǎn)生氧化應(yīng)激反應(yīng)。研究表明[15],絕經(jīng)后女性雌激素水平降低,抗氧化能力減弱,導(dǎo)致機(jī)體的ROS不斷堆積無法清除,誘導(dǎo)機(jī)體發(fā)生氧化應(yīng)激反應(yīng),而骨骼中過多的ROS會抑制成骨細(xì)胞的增殖、分化和骨基質(zhì)的礦化[16],最終導(dǎo)致骨質(zhì)疏松[17-18]。課題組前期研究發(fā)現(xiàn)[8],0.10 mmol/L H2O2能夠抑制MC3T3-E1成骨細(xì)胞增殖,使細(xì)胞處于氧化應(yīng)激狀態(tài),可作為成骨細(xì)胞氧化損傷模型;且六味地黃丸含藥血清體積濃度為20%時(shí),可促進(jìn)H2O2作用下的成骨細(xì)胞增殖活性。本研究結(jié)果再次證實(shí)0.10 mmol/L H2O2可以作為成骨細(xì)胞氧化損傷模型,20%六味地黃丸含藥血清以及抑制ERK/mTOR通路都可以抑制細(xì)胞內(nèi)ROS水平,減輕細(xì)胞氧化損傷。
有研究表明,自噬可以抑制氧化應(yīng)激造成的成骨細(xì)胞凋亡[19]。成骨細(xì)胞特異性敲除自噬基因的小鼠骨礦化能力降低,氧化應(yīng)激增加,骨小梁骨密度降低了50%[20]。自噬發(fā)生過程中最關(guān)鍵的是自噬小體的形成,LC3-Ⅱ通常被看作成自噬形成的標(biāo)志性分子。mTOR是缺乏氨基酸、生長因子、氧或ATP等細(xì)胞應(yīng)激的自噬反應(yīng)的匯集點(diǎn),其通過負(fù)向調(diào)節(jié)的反饋機(jī)制,誘導(dǎo)細(xì)胞自噬的發(fā)生[21]。雷帕霉素通過抑制mTOR通路激活細(xì)胞自噬。ERK通路能對細(xì)胞外各種信號刺激作出反應(yīng),起到一個(gè)細(xì)胞內(nèi)外信號傳導(dǎo)的作用[22]。研究表明,ERK被抑制后將導(dǎo)致mTOR信號下降,自噬信號提高,自噬通量增加[23]。U0126是ERK通路抑制劑,所以說抑制ERK通路也能促進(jìn)細(xì)胞自噬。
本研究發(fā)現(xiàn),六味地黃丸組、雷帕霉素組以及兩者聯(lián)合干預(yù)組的自噬蛋白LC3Ⅱ/Ⅰ比值都顯著提高,U0126組自噬蛋白有增高趨勢,提示六味地黃丸或者抑制ERK/mTOR通路可以促進(jìn)氧化應(yīng)激狀態(tài)下成骨細(xì)胞的自噬水平。朱仲康等[24]研究發(fā)現(xiàn)六味地黃丸能增加海馬神經(jīng)元中LC3表達(dá),提高海馬神經(jīng)元的自噬水平保護(hù)海馬神經(jīng)元細(xì)胞。本研究結(jié)果顯示,六味地黃組p-mTOR蛋白表達(dá)下調(diào),與雷帕霉素的作用相似,都可以抑制p-mTOR蛋白的表達(dá);六味地黃丸組p-ERK1/2蛋白表達(dá)下調(diào),與U0126的作用相似,都可以抑制p-ERK1/2蛋白的表達(dá)。六味地黃丸與U0126聯(lián)合作用組p-ERK1/2蛋白表達(dá)有下調(diào)趨勢,但還沒達(dá)到統(tǒng)計(jì)學(xué)意義,可能是六味地黃丸與U0126產(chǎn)生了某種化學(xué)反應(yīng),反而影響了相互的藥效。以上結(jié)果提示,六味地黃丸可以抑制ERK/mTOR通路。通過抑制ERK/mTOR通路,促進(jìn)氧化應(yīng)激狀態(tài)下成骨細(xì)胞的自噬,進(jìn)而抑制細(xì)胞ROS水平,減輕細(xì)胞氧化損傷。
綜上所述,六味地黃丸含藥血清通過抑制ERK/mTOR信號通路來誘導(dǎo)MC3T3-E1成骨細(xì)胞的自噬,以減輕細(xì)胞的氧化應(yīng)激損傷,這可能是六味地黃丸治療絕經(jīng)后骨質(zhì)疏松癥的機(jī)制之一。但本研究尚有不足之處,針對六味地黃丸抑制氧化應(yīng)激狀態(tài)下成骨細(xì)胞的自噬機(jī)制研究尚欠,后續(xù)將進(jìn)一步深入探究。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
中老年保健(2021年3期)2021-08-22 06:50:04
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
天津醫(yī)科大學(xué)學(xué)報(bào)(2021年2期)2021-03-29 05:31:08
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
世界科學(xué)技術(shù)-中醫(yī)藥現(xiàn)代化(2020年2期)2020-07-25 02:05:56
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
西南軍醫(yī)(2016年6期)2016-01-23 02:21:19
西南軍醫(yī)(2015年2期)2015-01-22 09:09:37
