全基因組關聯分析研究植物與微生物組的互作遺傳機制
2023-03-07 12:54:44李凱航王浩臣程可心楊艷金一何曉青
生物技術通報 2023年2期
李凱航 王浩臣 程可心 楊艷 金一 何曉青
(北京林業大學生物科學與技術學院,北京 100083)
自然界中,正常生長的植物表面和植物內部富集了數量龐大且種類繁多的微生物,這些微生物的集合被稱為植物微生物組[1]。植物為微生物組(包括細菌、真菌、原生生物、線蟲和病毒)的生長繁殖提供生態位點,在植物微生物組中數目最多的是細菌和真菌[2]。植物的健康與高度多樣化和動態的微生物組有關,微生物組對其宿主具有重要的功能[3-5],在促進植物礦質元素吸收、提高植物抗病性、協助植物抵抗非生物脅迫、直接調節植物生長發育等方面有重要影響[6-7]。植物與微生物的互作現象及其機制,是生命科學研究關注的熱點,也是利用微生物的關鍵問題。目前只有一小部分微生物與植物的相互作用得到了深入研究,其他大量微生物種類與功能缺乏系統性研究,它們與植物的互作機理尚不清楚[8-9]。隨著國內外微生物組研究的開展,植物微生物組的整體功能更受關注,闡明植物-微生物組互作的遺傳機制及互作在促進植物健康中的作用,將為植物的優良性狀培育提供新的思路,具有重要的科學意義和應用價值。
隨著測序技術和統計學的發展,GWAS 已成為有效方法來識別與微生物組相關的植物遺傳變異[4],并且已經被應用在多種植物上[10-14]。GWAS 是利用全基因組范圍內群體中高密度的分子標記,鑒定與復雜性狀表型變異相關聯的分子標記,進而挖掘與表型相關基因的方法。宿主由于基因型的差異直接影響了表型的變化,因此,GWAS 已經被廣泛應用于動物和植物以及人體的研究中[15-17]。而由于宿主基因型差異而改變的植物微生物組,同樣影響著植物的健康,可以將微生物組的差異作為植物的“拓展表型”,并利用關聯分析找到影響植物微生物組的基因[4],為分子育種等提供重要的理論基礎和遺傳資源,是未來研究方向的熱點。在植物與微生物組GWAS 分析中,如何表征微生物組差異并用于后續關聯是保證結果準確性的關鍵以及難點所在。
1 植物微生物組的結構和功能
植物并非單獨存在,而是與多樣化的微生物共存,在環境、宿主植物與微生物以及微生物與微生物之間的相互作用下共同塑造植物相關的微生物群落[7]。在不同的生態位展現了一個多樣性的植物微生物組,主要包括根際微生物組、葉際微生物組以及內生微生物組。受宿主和環境雙重影響,不同區系的微生物組在組成和功能上存在差異,是植物與土壤、大氣交互作用的媒介。
土壤作為微生物種子庫,為植物提供了豐富的微生物資源[18]。根際微生物主要來自于土壤,并召集了特定的微生物[19]。Wang 等[20]研究發現微生物的絕對數量從土壤到根際是逐漸增加的,提出根際微生物組“擴增-選擇”組裝新模型。有研究表明,相比于土壤,根際中的變形菌門、放線菌門和厚壁菌門細菌略有增加[2,21],根際微生物中的復雜微生物群落具有活化根區養分、促進植物生長、增強植物抗逆、抑制土傳病害等功能[22-27]。除了根瘤中的固氮細菌外,與自然界植物相關的其他根際微生物也已成為植物氮素營養的巨大來源[28-29]。植物定殖細菌甚至可以調節植物開花的時間[30-31],還能幫助宿主避免滅絕的風險[32]。最近的研究表明,植物根際微生物群可緩解植物干旱脅迫,使作物的生產可持續進行[26,33]。葉際能夠過濾來自更廣闊環境的微生物,并允許特定類型的微生物在植物定殖,進而服務于宿主和葉際生態系統[34-36]。葉際微生物組主要由變形菌門、擬桿菌門、厚壁菌門和放線菌門組成,其中變形菌門占群落組成的近50%[2]。葉際微生物組也已被證明會影響宿主健康和生長、對非生物脅迫的恢復力和對病原體的抵抗力,具體包括固氮作用、通過產生VOCs 提高抗逆并且阻止病原菌侵染、介導植物防御、抗干旱和紫外線以及促進植物代謝等[37-39]。植物內生菌群經常富含有變形菌門和厚壁菌門,而擬桿菌門的含量則較低[2],內生菌在促進植物生長、提高抗病性和緩解脅迫耐受性也發揮了重要作用[2]。根際內生菌可以幫助抵御病原菌的入侵,水稻種子內生菌可以幫助抗病等[40-41]。
2 植物對微生物組的影響
植物相關微生物組的巨大功能潛力已經得到證實,宿主因素對微生物組的影響也進行了大量的研究[42-43]。在特定的土壤類型及環境下,植物微生物組差異主要由宿主調控[44-45]。植物在募集微生物組、篩選有益微生物并且抵御有害菌以及調控微生物之間的互作等方面發揮重要作用。不同植物品種及基因型在對微生物的募集上表現出明顯的差異[21,26,46-47]。植物基因型影響了根系代謝物、免疫系統功能及根系分泌物的組成,進而影響了微生物組的活動與結構[6,48]。在面對生物或者非生物脅迫時,植物會通過遺傳因子整合脅迫信號并參與主動重塑植物微生物群,利用“呼救”(cry for help)策略招募有益菌[46,49-50],例如,小麥植株招募嗜麥芽寡養單孢菌(Stenotrophomonas rhizophilaSR80)來抑制土傳病原體[51]。甜菜根在根內層富集噬幾丁質菌科(Chitinophagaceae)和黃桿菌科(Flavobacteriaceae),這促進了植物抗真菌[40]。在最近的一項研究中,通過比較已感染受柑橘黑點病菌Diaporthe citri和未感染受柑橘黑點病菌D. citri柑橘葉之間的葉際微生物組的變化,發現柑橘會主動招募有益菌(包括泛菌Pantoeaasv90 和甲基桿菌Methylobacteriumasv41等)[39]。對優先考慮的番茄數量性狀位點的基因內容分析表明,各種根際菌系的不同招募有其遺傳基礎[52]。通過對143 個玉米自交系材料進行根系細菌群落組成研究發現,玉米根系細菌存在差異[53-54]。也有研究發現,由于物種繼承了祖先的遺傳物質和性狀,親緣關系越密切的物種比進化距離遙遠的物種微生物組更相似[55],說明宿主遺傳因素影響了微生物組組成。Bergelson 等[42]總結了植物影響微生物組的主要通路,包括生物鐘、生長發育、物理屏障、免疫系統、植物激素、初級代謝、次級代謝以及植物與微生物的共生。
2.1 植物基因型差異對微生物組網絡的影響
微生物在植物的作用下,通過競爭或者合作等方式,形成網絡結構來維持微生物組內部的穩定[6]。Xiong 等[56]研究發現,由于植物的選擇效應,影響了植物微生物組群落構建及網絡復雜度,網絡復雜度從土壤到根際到植物內部逐漸降低,并以葉內最低。不同生態位的微生物組也因植物基因型因素的差異而有區別,包括根際和葉際等(圖1)[29,34]。盡管研究人員已在多種植物中開展植物基因型影響微生物組結構的研究,但對植物如何控制微生物組平衡來影響其健康的探究相對不足[55],參與招募并塑造微生物組的主要基因有待挖掘。
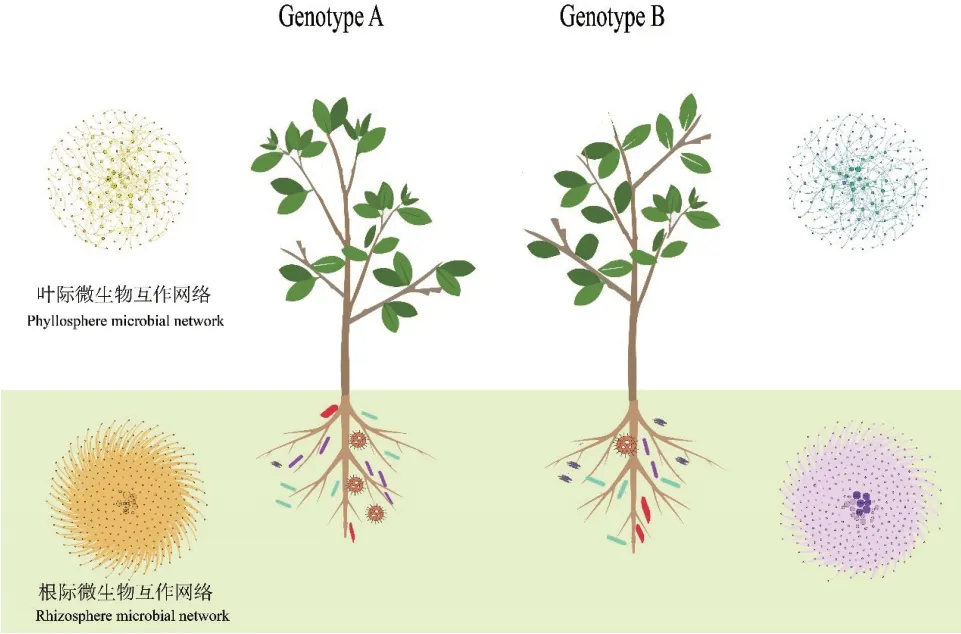
圖1 植物基因型差異影響根際和葉際的微生物互作網絡Fig. 1 Plant genotype differences affecting the microbial interaction network at the rhizosphere and phyllosphere
2.2 遺傳力表征植物對微生物以及微生物組的影響
為了解釋基因型和數量性狀之間復雜的關系,育種學家和遺傳學家已通過計算遺傳力來表示基因型對表型的影響大小。某一表型數據遺傳力越大,表示其與宿主基因型關系越大。植物微生物組差異作為一種數量性狀,也將遺傳力計算應用在植物基因對微生物組的影響上。諸多學者將植物微生物組豐度、α 和β 多樣性等參數視為數量性狀,估計它們的遺傳力,辨別受宿主遺傳因素調控的微生物群落,并進一步通過GWAS 分析挖掘導致植物微生物組可遺傳的宿主遺傳變異[11-12]。研究微生物組遺傳力,一方面可以加強我們對植物微生物組互作的影響,另一方面也可以作為幫助我們篩選益生菌的途徑,加強對有益微生物的利用。
針對于植物微生物組的遺傳力,其大小并非表示后代傳遞微生物組的能力,更應該準確的表示為描述最終由宿主遺傳引起的群落組裝的決定性成分[57]。通過對玉米根際微生物研究發現,單個微生物遺傳力大小范圍為0.15-0.25,雖然相比于開花時間的和其他傳統的農藝性狀來說遺傳力較低,但仍能體現出植物基因對特定分類群的影響。其中,遺傳力較高的微生物普遍為α 變形菌綱,并且發現7 種核心微生物(多個樣品中普遍出現的微生物)中有5 種屬于高遺傳力的微生物[54]。Brachi等[58]以擬南芥葉際微生物為研究對象,篩選出了既是可遺傳的并且在微生物互作網絡中為hub 微生物,屬于原霉菌(Protomyces inouyei)。Li 等[59]計算了單個微生物的遺傳力,發現有15 個屬于網絡中hub 微生物同樣具有相對較高的遺傳力(H2>0.10),包括變形菌門(Proteobacteria)、擔子菌門(Basidiomycota)、子囊菌門(Ascomycota)以及放線菌門(Actinobacteria)。Deng 等[12]計算了高粱根際微生物組的遺傳力值,以降維后的主成分值數據作為不同樣品的微生物組結構,研究發現,第一主成分解釋了微生物群落21%的方差,并且第一主成分遺傳力為0.35,證明了在高粱中微生物組具有較高的遺傳力。Wallace 等[11]對玉米葉際微生物組研究,計算出某些單個的微生物遺傳力較高,包括甲基桿菌屬(Methylobacterium,H2=0.457)等,利用PICRUST 預測的方法,提出大量微生物功能也具有遺傳性。
3 全基因組關聯分析在植物微生物組中的應用
研究植物與微生物組的互作遺傳機制需要構建系統的方法[4,19]。GWAS 是應對這一挑戰的一個強大而靈活的方法,由于GWAS 分析只需要表型數據和基因型數據,通過鑒定對宿主基因型敏感的微生物或者微生物組,并將它們與影響其定殖的基因位點聯系起來,幫助我們挖掘各種性狀顯著相關的數量性狀座位(SNP)及其候選基因位點。因此,全基因組關聯分析作為一種有效的手段幫助我們理解植物關鍵調控因子和相關信號途徑如何主動重塑相關微生物組組裝以及調控微生物組的穩定。
3.1 植物與微生物組關聯分析的數量遺傳模型的應用
GWAS 的基礎是連鎖不平衡(linkage disequilibrium, LD)[60],LD 的大小主要受群體遺傳多樣性的影響, 在不同物種和群體中差異很大。GWAS 需要考慮的問題包括群體的選取、群體結構分析、表型鑒定、全基因組關聯分析方法選擇及結果矯正[61]。在選擇微生物組作為植物的表型進行關聯分析時,首先是考慮對樣品的選擇,樣本量是保證結果可靠性的一個重要因素,一般選擇200 個以上[62]。然后進行群體結構分析,群體結構分析主要分為兩類,包括主成分分析(principal component analysis, PCA)和顯示生成模型的方法[61]。在獲得基因型數據和表型數據后進行關聯分析時,由于宿主群體結構以及個體間的關系,會導致關聯結果出現假陽性,在做關聯分析時,我們需要考慮群體結構(G)和親緣關系(K)。并將其作為協變量,通過引入一般線性模型(general linear model, GLM)和混合線性模型(mixed linear model, MLM)來矯正群落結構和親緣關系的影響,計算膨脹系數并結合Quantile-Quantile 圖選擇合適的模型篩選顯著性位點[61]。目前可用于GWAS的軟件包越來越多,Yin 等[63]利用R 語言,創建了rMVP 包,通過引入GLM、MLM 以及FarmCPU 三種模型,采用分塊矩陣計算、并行加速、計算流程優化等策略,加快了計算。同時,更多的GWAS 模型包括Meta-GWAS、多表型GWAS 等,其用于植物微生物組關聯分析上仍有待探索[4]。
3.2 微生物組差異作為植物表型的選擇
在應用關聯分析揭示植物對微生物組的調控中,如何表征以及量化植物體內的微生物組是我們需要思考的問題。傳統的研究主要依賴于在實驗室條件下開展植物與單一微生物的互作關系研究,很少在自然狀態下研究微生物組與宿主植物共存的機制[1],例如病原菌的豐度會影響植物健康,早期人們為了研究病原菌于宿主的關系,以病原菌豐度為表型數據調查植物對病原菌的調控[64];也有研究者對共生微生物進行研究,比如豆科植物和根瘤菌之間的關系[65]。隨著測序技術的發展,擴增子測序使得人們可以了解到植物體內的微生物群落。因此也有不少研究以相對豐度最高的微生物為表型數據進行關聯分析[10]。Bergelson 等[42]提出只考慮相對豐度高的微生物會忽略遺傳力高的微生物,確定可遺傳的微生物可能有助于建立宿主遺傳變異和微生物生態網絡之間的聯系。
當前人們越來越重視微生物組,希望可以深入了解植物與微生物組的互作機制。已有多種表征微生物組差異的類型,包括α 多樣性指數、β 多樣性指數、微生物組功能差異、hub 微生物的相對豐度及網絡特征值[4,13-14]。α 多樣性主要關注局域均勻生境下的物種數目,因此也被稱為生境內的多樣性,包括了Chao1、Ace、Shannon、Simpson、Richness 等指數。β 多樣性不僅可以反映樣本之間的多樣性距離關系,而且還可以反映生物群落之間的分化程度。其使用統計算法Euclidean,Bray-Curtis,Unweighted_unifrac,weighted_unifrac 等,計算兩兩樣品間距離,獲得距離矩陣,通過降維方式展示微生物組差異,包括主成分分析(principal component analysis,PCA)、主坐標分析(principal co-ordinates analysis,PCoA)、非度量多維尺度分析(non-metric multidimensional scaling,NMDS 分 析)、非 加 權 組平均聚類分析(unweighted pair-group method with arithmetic means,UPGMA)等分析方法,從中發現不同樣品之間微生物組的差異。還可以利用一些功能數據來表示微生物組之間的差異,包括利用宏基因組展示微生物組功能差異、基于擴增子測序數據預測的微生物群落功能(利用PICRUSt、Tax4Fun、FAPROTAX 及BugBase 等方法)[66-67]。
植物微生物組之間通過互作方式形成復雜的網絡結構,特別是細菌-真菌之間的相互作用,在植物生長中發揮重要作用[68]。在陸地生態系統的養分循環及植物群落動態等過程有重要調節作用[69-70]。微生物組差異也可以通過一些網絡參數體現出來,其中,一些hub 微生物在網絡中發揮重要作用,宿主也會通過影響hub 微生物來調控微生物互作網絡[71]。因此,有研究利用不同樣品間微生物互作網絡中的hub 微生物為表型數據來代表不同樣品之間的微生物組差異。通過了解對核心微生物的分析加強對微生物組的理解[13]。Jiang 等[72]用數學模型將微生物的競爭與合作系統分解為共生、拮抗、攻擊和利他網絡,He 等[14]整合行為生態學與基因作圖理論,將網絡嵌入到基因定位的框架,構建了網絡作圖模型(network mapping)(圖2)。利用數學模型計算了不同互作關系下的六種微生物互作網絡特征值(Connectivity、Closeness、Betweenness、Eccentricity、Eigenvector、PageRank),并以網絡特征值來表征不同樣品之間微生物互作網絡的差異。
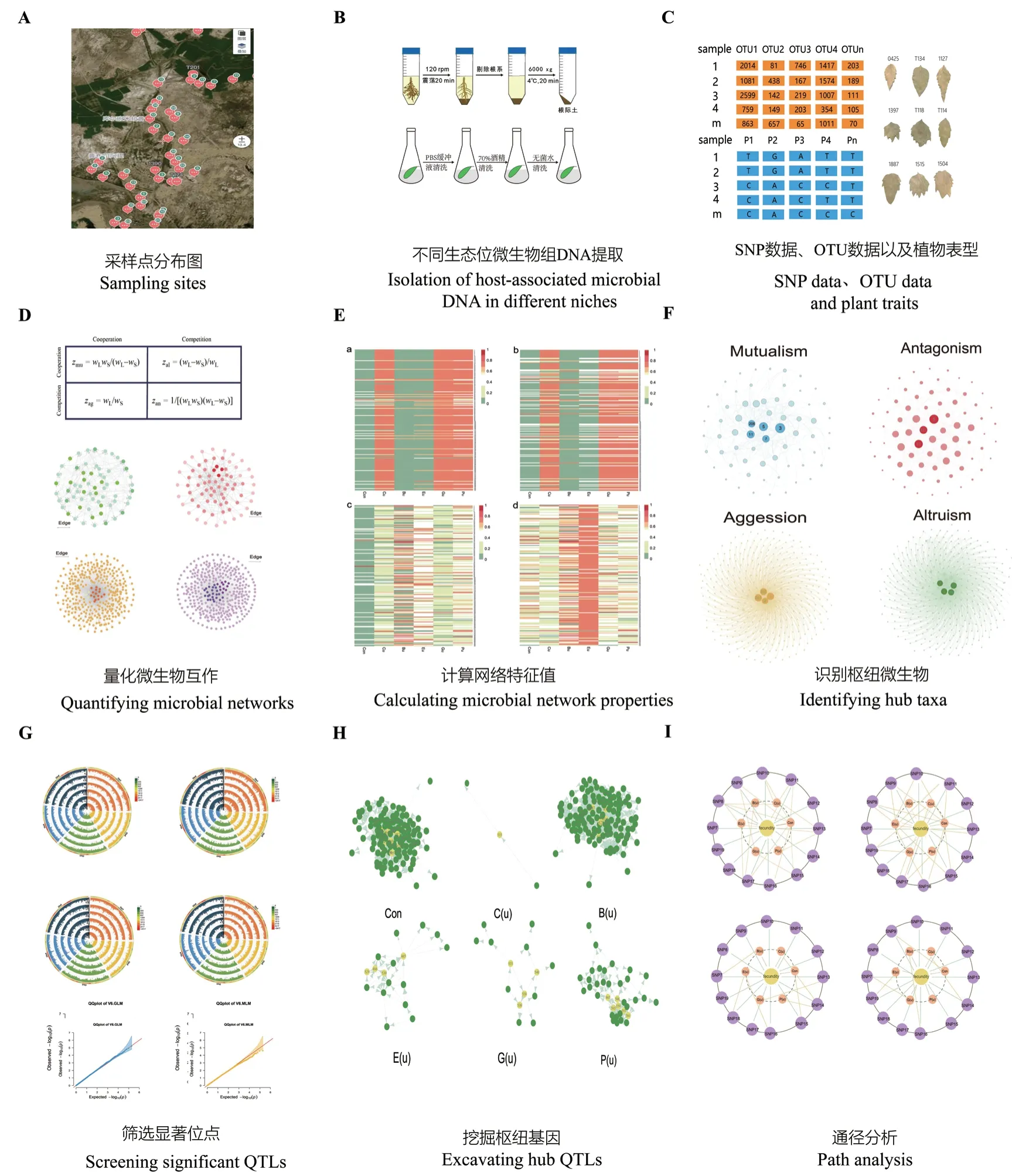
圖2 網絡作圖研究植物與微生物組互作Fig. 2 Studying plant-microbiome interactions by network mapping
3.3 利用GWAS進行的植物微生物組研究
選擇合適的微生物組表型差異數據后,可以將基因型數據與表型數據進行關聯,挖掘調控微生物組的植物候選基因。Wallace 等[11]計算了玉米葉際微生物組的α 和β 多樣性的多種參數(α 多樣性指數計算了除Michaelis-Menten 之外的其他所有參數,利用加權的UniFrac、未加權的UniFrac 和Bray-Curtis 距離度量計算了β 多樣性,同時利用PICRUST 的方法預測了微生物組的功能)并以此為表型數據進行關聯分析,發現了影響玉米葉際微生物組的管家基因(包括AGPv3等)。在最近的一項研究中,通過對高粱根際微生物組進行分析,以不同高粱根際微生物組PCA 結果的第一主成分來表征不同樣品間微生物組差異,選到RGA2、CHAF1A等基因,結果顯示這些基因表現出較強的根特異性活性[12]。Koprivova 等[73]以擬南芥根際土壤中微生物組硫酸鹽酶活性為表型數據,提出編碼色素P450 的基因影響根際硫酸鹽酶活性來反映出微生物組的差異。有研究以水稻葉際微生物互作網絡中的hub 微生物的相對豐度為表型數據,獲得了編碼氧化氫同工酶、ATP 依賴的蛋白酶等的基因(OsNippo02g019000、OsNippo11g101050)[13];He 等[14]將 擬 南 芥 基 因型與根際微生物組網絡特征值進行關聯,篩選到一些影響擬南芥根系生長以及一些抗逆基因,例如TMK3,IBR1B等;Li 等[59]同樣利用網絡作圖發現一些在植物生長中發揮重要作用的基因也影響了擬南芥葉際微生物互作,包括ADA2B,HAL3A等。同時,我們總結了在不同植物以及不同生態位中與微生物組有關的植物基因(表1)
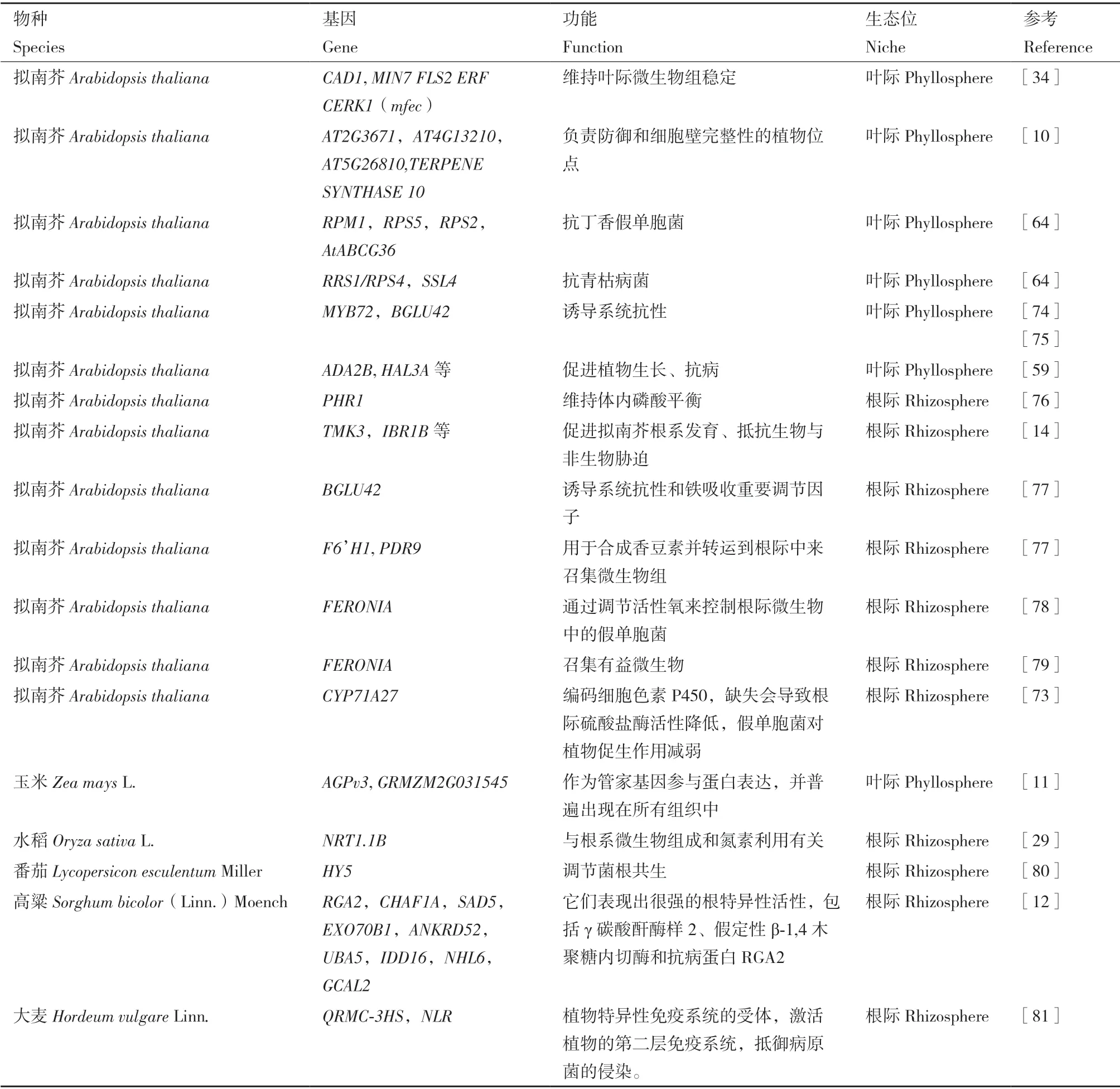
表1 與塑造微生物組有關的植物基因Table 1 Plant genes associated with the microbiome
4 展望
植物與自身微生物組作為一個整體,對于植物調控微生物組組裝的機制已有相關研究,微生物組在植物抗逆抗病等方面的作用也得到了廣泛證實,植物微生物組對植物生長發育的影響及其與植物的互作機制是國際前沿研究領域和重點研究方向[21]。微生物組差異作為植物的“拓展表型”應用于GWAS 幫助我們尋找可以調控微生物組的顯著位點,但是仍然有很多未來需要關注的方向。
首先,隨著測序技術迅速發展,提高了我們對微生物組的認識。為了進一步研究植物對微生物組的調控,我們仍然需要從多組學的角度出發,來量化微生物組功能差異,包括代謝組學以及蛋白質組學等,并將其作為表型數據引入到GWAS 分析中成為未來發展的方向。其次,為了充分發揮植物與微生物組之間的有益協作關系,保證微生物組在可持續農業中發揮更積極的作用,將篩選到的顯著基因應用到植物中,通過改善植物基因表達情況來操控微生物群的穩態或者優化微生物群,進而幫助植物健康生長和抵抗脅迫環境。
對于在關聯分析中尋找到的益生菌,合成菌群(SynComs)作為一種有效的手段,已經嘗試將微生物組利用在植物上[82]。有關微生物群落中樞紐菌株和關鍵物種的新信息為利用有益微生物提供了進一步的指導,通過培養和篩選核心微生物,進一步創制新型菌劑,作為挖掘利用微生物組的重要途經,加強對有益微生物的利用。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03