草魚單養(yǎng)和混養(yǎng)池塘的水質(zhì)與生物組成特征
2023-04-29 20:08:07肖述文劉興國陸詩敏趙宇曦顧兆俊周潤鋒
水生態(tài)學雜志 2023年6期
肖述文 劉興國 陸詩敏 趙宇曦 顧兆俊 周潤鋒
摘要:為比較單養(yǎng)、混養(yǎng)草魚(Ctenopharyngodon idella)養(yǎng)殖池塘的水質(zhì)與生物組成特點,采取水質(zhì)分析、環(huán)境DNA與傳統(tǒng)鑒別方法對草魚單養(yǎng)、混養(yǎng)(草魚:混養(yǎng)魚為80:20)兩種池塘的水質(zhì)變化、浮游生物、底棲生物、菌群結(jié)構(gòu)進行了分析。結(jié)果顯示:混養(yǎng)池塘的水質(zhì)優(yōu)于單養(yǎng)池塘,混養(yǎng)池塘水體中總氮(TN)、硝態(tài)氮(NO[-3]-N)、氨氮(NH[+4]-N)、亞硝態(tài)氮(NO[-2]-N)的濃度比單養(yǎng)池塘分別低10.15%、3.78%、5.07%、80.18%,總磷(TP)和活性磷(SRP)的濃度分別低27.14%和56.26%;兩類池塘中浮游植物均以綠藻門(Chlorophyta)、藍藻門(Cyanophyta)、隱藻門(Cryptophyta)為優(yōu)勢種,但單養(yǎng)池塘中的藻類密度為3.00×107 個/L,低于混養(yǎng)池塘1.04×108 個/L;兩類池塘中的浮游動物均以輪蟲和原生動物為優(yōu)勢種,枝角類和橈足類生物數(shù)量較少,單養(yǎng)池塘中浮游動物密度高于混養(yǎng)池塘;在底棲動物方面,單養(yǎng)池塘存在螺類、水蚯蚓和搖蚊幼蟲,而混養(yǎng)池塘僅有螺類和搖蚊幼蟲。在菌群組成方面,單養(yǎng)池塘水體中以厚壁菌門(Firmicutes)為優(yōu)勢類群,混養(yǎng)池塘水體中以變形菌門(Proteobacteria)為優(yōu)勢類群;但在兩種池塘底泥中,均以變形菌門為優(yōu)勢類群。研究表明,草魚混養(yǎng)有利于改善養(yǎng)殖池塘水質(zhì),增加浮游植物豐富度,改變養(yǎng)殖水體菌群的結(jié)構(gòu)。研究結(jié)果為優(yōu)化草魚池塘養(yǎng)殖結(jié)構(gòu)、改善水質(zhì)、構(gòu)建高效池塘養(yǎng)殖模式提供了依據(jù)。
關(guān)鍵詞:草魚池塘;養(yǎng)殖模式;水質(zhì)指標;生物組成;環(huán)境DNA
中圖分類號:S964.3,Q178.1? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2023)06-0079-09
草魚(Ctenopharyngodon idella)是中國年產(chǎn)量最高的淡水魚類,有池塘養(yǎng)殖、網(wǎng)箱養(yǎng)殖、水庫養(yǎng)殖、稻田養(yǎng)殖等多種模式,其中池塘養(yǎng)殖占草魚養(yǎng)殖總產(chǎn)量的74%(國家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系,2021);在華南、華東、華中等草魚主產(chǎn)區(qū),其池塘養(yǎng)殖模式主要有單養(yǎng)和混養(yǎng)兩種(常華,2009;楊帆,2016;陳小江和汪祈超,2016;梅順和任青松,2017)。草魚單養(yǎng)模式由于放養(yǎng)密度大,輪捕輪放能獲得較高的收益,但存在水質(zhì)管理難度大、易暴發(fā)疾病等問題(劉玲儀和金有坤,1994;高攀等,2009;劉朋, 2012);草魚混養(yǎng)模式主要為80:20方式(草魚:混養(yǎng)魚),混養(yǎng)品種多為鰱(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、鯽(Carassius auratus)等(占家智和羊茜,2015);其中鰱、鳙可攝食浮游動植物和有機碎屑,抑制藍綠藻的過度生長并改善養(yǎng)殖水質(zhì)(孫云飛,2013),鯽作為底層魚類能夠充分利用水體空間,攝食底層殘餌。近年來,隨著養(yǎng)殖尾水問題加重,這種多營養(yǎng)層級的草魚混養(yǎng)模式被廣泛應用,通過充分利用系統(tǒng)空間和資源,提高了系統(tǒng)內(nèi)營養(yǎng)物質(zhì)的循環(huán)利用率,防止了病害發(fā)生并改善水質(zhì)(董雙林,2014;周波,2015)。
環(huán)境DNA(Environmental DNA, eDNA)方法是從環(huán)境樣本中提取DNA,針對目標類群設計通用引物,通過PCR擴增并結(jié)合高通量測序,對環(huán)境中的目標物種進行識別和分析(Taberlet et al, 2012)。環(huán)境DNA方法能夠測定傳統(tǒng)方法中難以觀察到的物種,不會對物種和生態(tài)系統(tǒng)造成破壞,也能更加準確、快速地判斷目標物種(巴日斯,2018)。近年來,環(huán)境DNA分析技術(shù)已被廣泛應用于海洋、湖泊、池塘等生態(tài)系統(tǒng)中的無脊椎動物、浮游植物、細菌等生物檢測(李萌等,2019;舒璐等,2020;張麗娟等,2021;許鄭超,2021;Singer et al, 2021);而我國應用環(huán)境DNA技術(shù)調(diào)查池塘生態(tài)系統(tǒng)中生物的研究并不常見。目前主要采用傳統(tǒng)方法來監(jiān)測池塘中浮游動植物及底棲生物等(李瑞嬌,2014;葉曉彤等,2020;陳佳林等,2021),但傳統(tǒng)方法存在監(jiān)測方法復雜、物種難以鑒別等問題,故本文將兩種方法相結(jié)合以快速、有效地開展池塘浮游生物的監(jiān)測工作。
目前,我國草魚池塘養(yǎng)殖依然存在飼料利用率低、水質(zhì)調(diào)控難等問題,亟需系統(tǒng)研究不同養(yǎng)殖模式的生態(tài)特征,優(yōu)化池塘生態(tài)系統(tǒng)(Li et al,2019;Niu et al,2023)。本文采用水質(zhì)分析、環(huán)境DNA技術(shù)及傳統(tǒng)鑒別方法,對單養(yǎng)和混養(yǎng)草魚養(yǎng)殖模式中的水質(zhì)、浮游生物、底棲生物、細菌等進行研究,旨在為評價草魚養(yǎng)殖模式、構(gòu)建高效草魚池塘系統(tǒng)提供依據(jù),并對環(huán)境DNA方法監(jiān)測池塘生態(tài)系統(tǒng)中出現(xiàn)的問題進行探究。
1? ?材料與方法
1.1? ?試驗池塘
在上海市松江區(qū)三泖水產(chǎn)養(yǎng)殖場選取兩類草魚養(yǎng)殖池塘進行試驗,池塘規(guī)格為100 m × 50 m,平均水深1.6 m,水面積0.47 hm2。
單養(yǎng)池塘放養(yǎng)0.57 kg/尾的草魚1 322尾,出于實際養(yǎng)殖需要,考慮到鰱并不攝食飼料,因此在單養(yǎng)塘投入0.15 kg/尾的鰱277尾;混養(yǎng)(80:20)池塘放養(yǎng)草魚897尾、0.50 kg/尾,鰱123尾、1.25 kg/尾,鳙529尾、0.26 kg/尾,鯽158尾、0.09 kg/尾,鳙魚苗2 000尾、0.5 g/尾,鯽魚苗1 000尾、0.25 g/尾。
試驗期間的增氧、投飼等管理方式一致,養(yǎng)殖期間不施肥、不噴灑殺蟲劑。
1.2? ?樣本采集及測定
選取池塘對角線上的3個點,使用3點法對池塘水體及底泥樣品進行采集,測定理化指標并采集池塘中的生物樣本。水樣使用5 L有機玻璃采水器于池塘1 m深處進行采集,底泥使用規(guī)格為1/40 m2彼得森采泥器抓取。
水體理化指標采取國標法測定。總氮(TN)采用GB 11894-89(國家環(huán)境保護局規(guī)劃標準處,1989)過硫酸鉀氧化紫外分光光度法,硝態(tài)氮(NO3--N)采用HJ/T346-2007(國家環(huán)境保護總局科技標準司,2007)紫外分光光度法,氨氮(NH4+-N)采用HJ 535-2009(國家環(huán)境保護總局科技標準司,2009)納氏試劑光度法,亞硝態(tài)氮(NO2--N)采用GB 7493-87(國家環(huán)境保護局規(guī)劃標準處,1987)N-(1-萘基)-乙二胺光度法,總磷 (TP)、活性磷(SRP)采用GB 11893-89(國家環(huán)境保護局標準處,1989)鉬銻抗分光光度法。
底泥總氮、總磷采取水浸法預處理,濾液采取國標法測定。
浮游生物與底棲生物的采樣定量參考《水和廢水監(jiān)測分析方法》(國家環(huán)境保護總局,2002)。
1.3? ?環(huán)境DNA樣品
3點法對池塘水樣和底泥取樣后,等體積均勻混合,取1.0 L水體及50.0 g底泥各為1個檢測樣本。每100 mL水樣使用0.22 μm的濾膜進行抽濾處理,濾膜和底泥樣本置于-20℃冷凍保存,送至上海派森諾生物科技有限公司提取DNA并進行后續(xù)測序分析。
分別選取引物16S V3V4、18S V4、COI用于細菌、浮游植物、浮游動物的PCR擴增。細菌擴增選取16S V3V4區(qū)域,上游引物:ACTCCTACGGGAGGCAGCA,下游引物:GGACTACNVGGGTWTCTAAT;浮游植物選取18S V4區(qū)域,上游引物:CCAGCASCYGCGGTAATTCC,下游引物:ACTTTCGTTCTTGATYRA;浮游動物選取COI基因上的區(qū)域,上游引物:GGWACWGGWTGAACWGTWTAYCCYCC,下游引物:TANACYTCNGGRTGNCCRAARAAYCA。
擴增體系(25 μL):5 μL 5×reaction緩沖液,5 μL 5×GC緩沖液,2 μL dNTPs(2.5 mM),正反引物(10 μM)各1 μL,2 μL DNA模板(20 ng/μL),8.75 μL ddH2O,0.25 μL Q5 DNA聚合酶。反應條件為98℃預變性2 min,98℃變性15 s、55℃退火30 s、72℃延伸30 s、25~30個循環(huán),72℃最后延伸5 min。
1.2%瓊脂糖凝膠電泳檢測PCR擴增結(jié)果,之后通過Illumina NovaSeq測序平臺進行高通量測序和序列處理工作。
1.4? ?數(shù)據(jù)處理與分析
測序的原始數(shù)據(jù)利用Vsearch軟件進行去引物、拼接、質(zhì)量過濾、去重、去嵌合體、聚類等步驟后,將得到的序列與數(shù)據(jù)庫進行對比,進行α-多樣性指數(shù)分析。細菌選取Silvar數(shù)據(jù)庫,浮游植物和浮游動物選取NCBI數(shù)據(jù)庫。
α-多樣性指數(shù)包括物種豐富度指數(shù)(Chao1指數(shù))、物種多樣性指數(shù)(Shannon指數(shù))、覆蓋率(Good's Coverage),均使用QIIME軟件計算。
利用Excel對數(shù)據(jù)進行初步統(tǒng)計,通過SPSS 26.0軟件對池塘數(shù)據(jù)進行獨立樣本T檢驗分析,使用Origin 2016作圖。
2? ?結(jié)果與分析
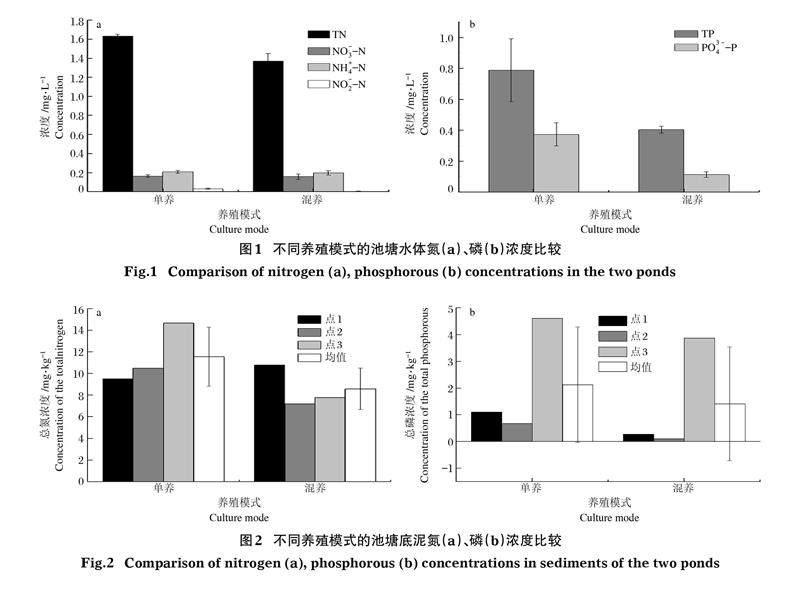
2.1? ?池塘環(huán)境氮磷濃度
對試驗期間兩池塘水質(zhì)進行檢測,單養(yǎng)塘中的總氮、硝態(tài)氮、氨氮、亞硝態(tài)氮的濃度分別為(1.633± 0.018)、(0.164±0.015)、(0.207±0.016)、(0.031±0.004)mg/L,混養(yǎng)塘中的對應濃度分別為(1.368±0.083)、(0.158±0.029)、(0.197±0.0240)、(0.004±0.002)mg/L,混養(yǎng)池塘水體中總氮、硝態(tài)氮、氨氮、亞硝態(tài)氮的濃度比單養(yǎng)池塘分別低10.15%、3.78%、5.07%、80.18%(圖1-a)。兩池塘中的總氮、亞硝態(tài)氮濃度差異顯著(P<0.05),硝態(tài)氮、氨氮濃度差異不顯著(P>0.05)。
混養(yǎng)池塘中總磷和活性磷的濃度比單養(yǎng)池塘低27.14%和56.26%(圖1-b)。單養(yǎng)池塘總磷和活性磷的濃度分別為(0.788±0.204)、(0.373 ± 0.075)mg/L,混養(yǎng)池塘為(0.403±0.022)、(0.112 ± 0.018)mg/L,兩池塘中的總氮和活性磷濃度均存在顯著差異(P<0.05)。
兩池塘底泥中的氮、磷含量檢測結(jié)果見圖2。單養(yǎng)池塘3個采樣點(點1、點2、點3)的總氮濃度分別為9.509、10.484、14.651 mg/kg,混養(yǎng)池塘分別為10.774、7.192、7.772 mg/kg;單養(yǎng)池塘3個采樣點的總磷濃度分別為1.100、0.670、4.606 mg/kg,混養(yǎng)池塘分別為0.273、0.094、3.869 mg/kg。兩池塘各區(qū)域總氮、總磷的含量均有所差異,總氮在各區(qū)域分布無明顯規(guī)律而總磷的含量差異較大,位于出水口附近區(qū)域的總磷含量尤其高,可能是由于飼料中未分解的磷元素沉積到底泥并被水流沖刷到出水口造成的。
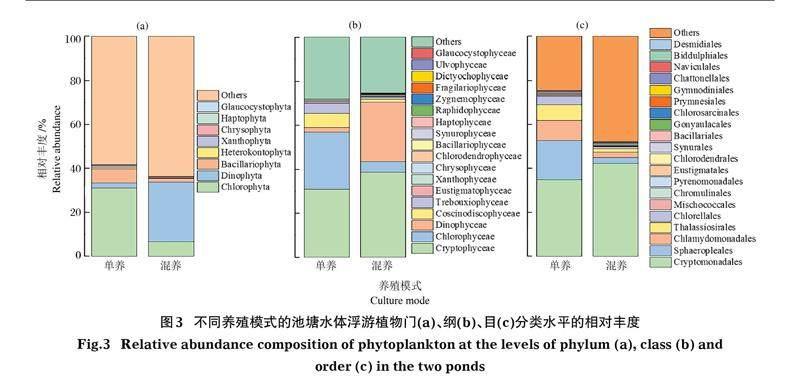
2.2? ?浮游植物相對豐度和密度
采用環(huán)境DNA檢測兩池塘的浮游植物組成,其相對豐度結(jié)果見圖3。池塘中的主要藻類為綠藻門(Chlorophyta)、甲藻門(Dinophyta)、隱藻門(Cryptophyta)、硅藻門(Bacillariophyta)、金藻門(Chrysophyta)、黃藻門(Xanthophyta)、異鞭藻門(Heterokontophyta)。
草魚單養(yǎng)池塘共計9門14綱17目,以綠藻門(31.06%)為優(yōu)勢類群,門水平檢測出來的藻類達41.73%。綱分類水平檢測出來的藻類占72.07%,以隱藻綱(Cryptophyceae)30.85%為優(yōu)勢,其次是綠藻綱(Chlorophyceae)25.97%、圓篩藻綱(Coscinodiscophyceae)6.32%、共球藻綱(Trebouxiophyceae)4.57%、甲藻綱(Dinophyceae)2.22%。目分類水平檢出藻類占78.28%,以隱鞭藻目(Cryptomonadales)30.85%為優(yōu)勢。
混養(yǎng)池塘共計10門17綱19目,以甲藻門(26.93%)為優(yōu)勢類群,門水平檢測出來的藻類可達36.30%。綱分類水平檢測出來的藻類占74.60%,隱藻綱38.68%、甲藻綱26.93%、綠藻綱4.81%、圓篩藻綱1.50%、共球藻綱0.95%。目分類水平檢出藻類占56.30%,以隱鞭藻目38.68%為優(yōu)勢。
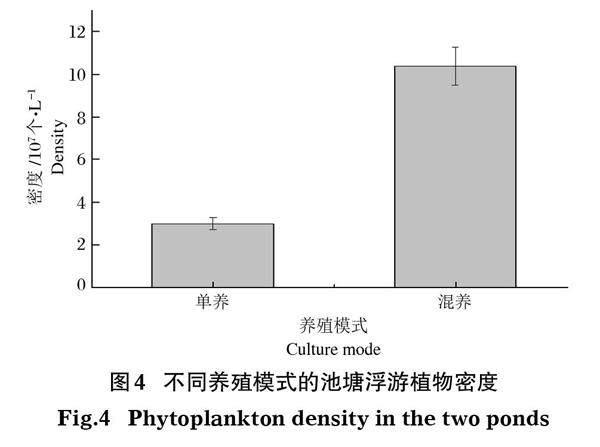
試驗期間,使用傳統(tǒng)方法對池塘中藻類進行鑒別計數(shù)(圖4)。結(jié)果表明,單養(yǎng)池塘的浮游植物為3.00×107個/L,混養(yǎng)池塘可達1.04×108個/L,混養(yǎng)池塘中的浮游植物數(shù)目高于單養(yǎng)池塘,兩池塘浮游植物密度存在顯著性差異(P<0.05)。
2.3? ?浮游動物和底棲動物豐度
兩類池塘浮游動物和底棲動物組成及相對豐度見圖5。單養(yǎng)池塘浮游動物和底棲動物共10門,混養(yǎng)池塘9門,兩類池塘均以袋形動物門(Aschelminthes)、節(jié)肢動物門(Arthropoda)、軟體動物門(Mollusca)、脊索動物門(Chordata)、刺胞動物門(Cnidaria)為主,棘皮動物門(Echinodermata)、紐形動物門(Nemertea)、海綿動物門(Porifera)等檢測量均不超過1%。目分類水平上,兩類池塘中浮游動物均以游泳輪蟲目(Ploima)為優(yōu)勢,豐度分別可以達到21.54%和20.83%,存在劍水蚤目(Cyclopoida)、雙甲目(Diplostraca)、哲水蚤目(Calanoida)。屬分類水平上,兩池塘以多肢輪屬(Polyarthra)、無柄輪屬(Ascomorpha)為優(yōu)勢,存在溫劍水蚤屬(Thermocyclops)、華哲水蚤屬(Sinocalanus)、臂尾輪屬(Brachionus)、龜甲輪屬(Keratella),單養(yǎng)池塘發(fā)現(xiàn)少量偽鏢水蚤屬(Pseudodiaptomus),混養(yǎng)池塘未發(fā)現(xiàn)。
采取傳統(tǒng)方法對兩池塘中原生動物(Protozoa)、輪蟲(Rotifera)、枝角類(Cladocera)、橈足類(Copepoda)、底棲動物的鑒別計數(shù)結(jié)果見圖6-a。兩池塘均以原生動物和輪蟲為優(yōu)勢,存在枝角類、橈足類等甲殼動物,輪蟲均以多肢輪屬為優(yōu)勢,其次是無柄輪屬、臂尾輪和少量異尾輪屬(Trichocercidae)和龜甲輪屬,橈足類多為劍水蚤和哲水蚤,單養(yǎng)池塘的浮游動物數(shù)目高于混養(yǎng)池塘,兩池塘橈足類密度差異顯著(P<0.05)。
使用傳統(tǒng)方法對底棲生物的計數(shù)結(jié)果見圖6-b。單養(yǎng)池塘發(fā)現(xiàn)了腹足動物(Gastropoda)、寡毛類(Oligochaeta)和搖蚊幼蟲(Chironomidae),而混養(yǎng)池塘觀察到腹足動物(螺類)和搖蚊幼蟲。混養(yǎng)池塘的螺類多于單養(yǎng)池塘。
2.4? ?菌群組成
2.4.1? ?菌群豐度? ?使用環(huán)境DNA技術(shù)檢測池塘中細菌的相對豐度,結(jié)果如圖7所示。
單養(yǎng)池塘水體中微生物共計15門27綱38目,優(yōu)勢菌群為厚壁菌門(Firmicutes)81.86%,其次是變形菌門(Proteobacteria)6.40%、放線菌門(Actinobacteria)5.40%、擬桿菌門(Bacteroidetes)2.20%。綱分類水平以芽孢桿菌綱(Bacilli)81.76%占優(yōu),其次是γ-變形菌綱(Gammaproteobacteria)4.33%,放線菌綱(Actinobacteria)4.26%。屬分類水平,以微小桿菌屬(Exiguobacterium)77.40%占優(yōu)。
單養(yǎng)池塘底泥中微生物共檢測出46門,以變形菌門44.61%為優(yōu)勢菌,其次為厚壁菌門23.42%、綠彎菌門(Chloroflexi)11.22%、擬桿菌門5.67%、酸桿菌門(Acidobacteria)24.97%、放線菌門2.92%。綱分類水平以γ-變形菌綱34.64%占優(yōu),其次是芽孢桿菌綱18.41%,厭氧繩菌綱(Anaerolineae)10.36%,δ-變形菌綱(Deltaproteobacteria)8.06%,擬桿菌綱(Bacteroidia)4.69%,梭狀芽胞桿菌(Clostridia)4.69%。屬分類水平以Paenisporosarcina(13.90%)占優(yōu)。
混養(yǎng)池塘水體中微生物共16門34綱46目,優(yōu)勢菌群為變形菌門44.15%,其次是厚壁菌門35.08%、放線菌門8.66%、擬桿菌門2.68%、疣微菌門(Verrucomicrobia)2.20%。綱分類水平以γ-變形菌綱41.02%占優(yōu),其次是芽孢桿菌綱34.89%,放線菌綱6.96%,產(chǎn)氧光細菌綱(Oxyphotobacteria)5.75%。屬分類水平,以不動桿菌屬(Acinetobacter)(35.52%)占優(yōu)。
底泥中微生物共37門,變形菌門54.56%為優(yōu)勢菌,還存在厚壁菌門16.11%、放線菌門8.20%、綠彎菌門6.77%、擬桿菌門4.75%、酸桿菌門4.68%。綱分類水平,以γ-變形菌綱45.25%占優(yōu),其次是芽孢桿菌綱15.09%,放線菌綱6.80%,α-變形菌綱(Alphaproteobacteria)4.86%,δ-變形菌綱4.41%,擬桿菌綱4.24%。屬分類水平以假單胞菌屬(Pseudomonas)(16.51%)占優(yōu)。
2.4.2? ?α-多樣性指數(shù)? ?以97%序列相似度為閾值生成操作性分類單元(Operational Taxonomic Units, OTUs)后,計算得出α-多樣性指數(shù)如表1所示。水體中Chao1指數(shù)和Shannon指數(shù)均為混養(yǎng)池塘>單養(yǎng)池塘,而底泥中Chao1指數(shù)和Shannon指數(shù)均為單養(yǎng)池塘>混養(yǎng)池塘。無論是Chao1指數(shù)還是Shannon指數(shù)均為底泥樣本>水體樣本,這表明兩池塘底泥中菌群的豐富度和多樣性均高于水體,混養(yǎng)池塘水體中菌群的豐富度和多樣性高于單養(yǎng)池塘,而單養(yǎng)池塘底泥中菌群的豐富度和多樣性高于混養(yǎng)池塘。
3? ?討論
本研究結(jié)合環(huán)境DNA和傳統(tǒng)鑒別方法對單養(yǎng)和混養(yǎng)兩種草魚池塘養(yǎng)殖模式的生物組成進行了鑒別和分析,從生態(tài)角度對兩種養(yǎng)殖模式進行了比較,在水質(zhì)、浮游生物、菌群結(jié)構(gòu)等方面具體分析了兩類草魚池塘的差異,為構(gòu)建物質(zhì)能量模型、進一步分析養(yǎng)殖生態(tài)系統(tǒng)的結(jié)構(gòu)特征提供了依據(jù)。使用傳統(tǒng)方法對池塘物種鑒別較為準確,但采樣操作復雜、觀測難度大,且會對生境造成破壞;環(huán)境DNA方法的效率和靈敏度高,能夠監(jiān)測環(huán)境中隱蔽和難以識別的物種,但易受環(huán)境中其他因素污染。環(huán)境DNA結(jié)合傳統(tǒng)方法進行物種鑒別,不僅能比較兩種方法的差異,二者也相互補充,提高了對養(yǎng)殖系統(tǒng)內(nèi)生物分析的準確性。
3.1? ?環(huán)境DNA鑒別方法存在一定的局限性
本研究結(jié)果中,環(huán)境DNA方法和傳統(tǒng)鑒別方法得出的結(jié)論有部分出入,如二者對藻類的結(jié)果有出入,環(huán)境DNA方法監(jiān)測到池塘中不存在的物種等。這種差異可能由于鳥類、昆蟲的停靠及降水帶來的DNA等對池塘DNA造成了一定污染;底泥沉積中未被降解的DNA也可能會使不存在的物種被檢測到,從而導致環(huán)境DNA監(jiān)測結(jié)果出現(xiàn)一定偏差;同時,池塘水體過小且流動能力弱,使DNA無法均勻分散而在某一區(qū)域堆積,也可能造成調(diào)查的偏差;而環(huán)境DNA的靈敏性也使得其在擴增測序及對比分析的過程中易產(chǎn)生偏差。因此,對于目標物種引物以及數(shù)據(jù)庫的選擇需要慎重,使用傳統(tǒng)監(jiān)測手段對環(huán)境DNA技術(shù)進行補充驗證是必要的。
3.2? ?混養(yǎng)池塘高密度藻類有助于吸收水體氮磷
本試驗結(jié)果表明,單養(yǎng)池塘水體和底泥中的氮磷含量均高于混養(yǎng)池塘,這與其他學者的研究結(jié)論相似(劉朋,2012;孫云飛,2013)。養(yǎng)殖水體中的氮磷輸入以飼料投放為主,過度投放的飼料未被養(yǎng)殖生物完全攝食,會增加水體中有機質(zhì)的含量。單養(yǎng)池塘本身生物組成簡單、食物鏈短,也會造成營養(yǎng)物質(zhì)的浪費,水質(zhì)容易受到影響;而混養(yǎng)池塘中的鰱、鳙等通過濾食作用過濾掉水中的殘餌,減少飼料中有機物的溶解釋放,對水質(zhì)凈化起到了一定作用。氮磷是藻類增殖所必需的營養(yǎng)鹽(孫靈毅等,2003),養(yǎng)殖水體中的溶解性氮磷在藻類生長過程中被大量消耗吸收,能夠降低池塘中氮磷含量。混養(yǎng)池塘的藻類密度遠高于單養(yǎng)池塘,因此對水中氮磷的吸收利用更好(張雷鳴等,2020),對養(yǎng)殖水體能起到一定的凈化效果。
3.3? ?池塘混養(yǎng)可減少藍綠藻并使浮游動物小型化
試驗發(fā)現(xiàn),單養(yǎng)池塘和混養(yǎng)池塘的浮游植物主要為綠藻門、藍藻門和隱藻門,硅藻、甲藻和少量金藻也在兩類池塘觀察到,單養(yǎng)池塘以衣藻(Chlamydomonas)、隱藻屬(Cryptomonas)、海鏈藻(Thalassiosira)為優(yōu)勢,混養(yǎng)池塘以隱藻屬為優(yōu)勢。這與李瑞嬌(2014)的研究結(jié)果相似,即草魚池塘養(yǎng)殖期間,浮游植物優(yōu)勢種集中在藍藻門、隱藻門和綠藻門;Turker等(2003)研究發(fā)現(xiàn),鰱、鳙能夠通過濾食作用顯著減少藍綠藻的數(shù)量,這可能是混養(yǎng)池塘以隱藻屬為優(yōu)勢種的原因之一。大量研究表明,適量放養(yǎng)鰱、鳙等濾食性魚類,可促進水體養(yǎng)分循環(huán),有助于浮游植物密度和生物量的增加(趙玉寶,1993;谷孝鴻和劉桂英,1996;李瑞嬌,2014),這與本試驗混養(yǎng)池塘的藻類密度高于單養(yǎng)池塘的結(jié)果相符。
單養(yǎng)池塘和混養(yǎng)池塘中浮游動物、底棲生物組成相似,均以多肢輪屬、無柄輪屬、溫劍水蚤屬、華哲水蚤屬、異尾輪科等為優(yōu)勢。輪蟲和原生動物均具有孤雌生殖方式,生長發(fā)育迅速,能在短期內(nèi)達到較高的豐度(陳佳林等,2021),因此在池塘生態(tài)系統(tǒng)的浮游動物中占優(yōu)勢。兩類池塘中輪蟲較多且大都為廣溫性種類,水體均傾向于中營養(yǎng)化狀態(tài)(葉曉彤等,2020)。單養(yǎng)池塘的浮游動物數(shù)目明顯高于混養(yǎng)池塘,與陳雷等(2009)研究結(jié)果不符,可能由于混養(yǎng)池塘中的甲藻對浮游動物產(chǎn)生一定的危害性;而混養(yǎng)池塘放養(yǎng)的濾食性鰱、鳙對浮游動物的攝食壓力增大(王玉彬,2007),也會導致浮游動物數(shù)量減少并趨向小型化(陳光榮等,2008)。
3.4? ?池塘水體和底泥優(yōu)勢菌群及其生態(tài)環(huán)境功效
兩類池塘菌群均以變形菌門和厚壁菌門為優(yōu)勢類群,但菌群的結(jié)構(gòu)和相對豐度有所差異。單養(yǎng)池塘水體以厚壁菌門(81.86%)、芽孢桿菌綱(81.76%)、微小桿菌屬(77.40%)為優(yōu)勢菌,混養(yǎng)池塘水體以變形菌門(44.15%)、γ-變形菌綱(41.02%)、不動桿菌屬(35.52%)為優(yōu)勢菌。兩類池塘底泥均以變形菌門、γ-變形菌綱和厚壁菌門、芽孢桿菌綱為優(yōu)勢亞群,以Paenisporosarcina和假單胞菌屬為優(yōu)勢菌群。底泥中的菌群比水體豐富,存在大量的厭氧型菌群,與氮循環(huán)關(guān)系密切的Nitrospirae和Nitrospinae在底泥中的含量遠高于水體。池塘中的微小桿菌屬在降解有機物、去除重金屬、脫蛋白以及處理有機高鹽廢水等方面發(fā)揮著重要的作用(徐蓮等,2012;張瑩等,2013)。γ-變形菌作為海洋生態(tài)系統(tǒng)中普遍存在的細菌,在有機質(zhì)含量較高的水域中多以硫氧化細菌及固氮細菌為優(yōu)勢菌群(Ivanova et al, 2003;Payne et al, 2006),能夠在厭氧條件下進行光合作用,對環(huán)境中的碳、硫循環(huán)起著重要作用(白潔等,2009;Bakunina et al, 2020)。本試驗還發(fā)現(xiàn),混養(yǎng)池塘水體的菌群多樣性和豐富度均高于單養(yǎng)池塘,而在底泥中呈現(xiàn)相反的趨勢,這與田相利等(2012)對草魚養(yǎng)殖初期菌群多樣性的研究結(jié)論不一致,與養(yǎng)殖末期菌群多樣性的結(jié)論一致。可見在養(yǎng)殖過程中菌群的演替已經(jīng)開始,試驗結(jié)果同時也受到溫度、飼料投放、養(yǎng)殖水體等外界條件的影響。
綜上所述,合理混養(yǎng)可以充分利用飼料(劉朋,2012)、調(diào)節(jié)養(yǎng)殖系統(tǒng)中的菌落結(jié)構(gòu)(李秋芬等,2002)、改變浮游動植物的豐富度(張雷鳴等,2020)。作為我國目前應用較廣的兩種草魚池塘養(yǎng)殖模式,單養(yǎng)和混養(yǎng)草魚在浮游生物、菌群組成與豐富度上存在著一定的差異性。草魚養(yǎng)殖池塘中合理混養(yǎng)鰱、鳙等濾食性魚類,能夠有效地降低水體中氮磷的含量,增加浮游植物的數(shù)目,改變菌群的結(jié)構(gòu)和豐富度。草魚混養(yǎng)對改善池塘的水質(zhì)、浮游生物以及菌群的組成結(jié)構(gòu)等方面存在一定的正面作用,而兩種養(yǎng)殖方式在生態(tài)系統(tǒng)結(jié)構(gòu)、能量流動以及營養(yǎng)動力學等方面的具體特征和差異,以及能否通過改善魚種的搭配提高生態(tài)效益、經(jīng)濟效益還有待進一步探究。
參考文獻
巴日斯,2018. 環(huán)境DNA及其在水生生態(tài)系統(tǒng)保護中的應用研究[J]. 資源節(jié)約與環(huán)保, (3):17,25.
白潔,李海艷,趙陽國,2009. 黃海北部不同站位海洋細菌群落分布特征[J]. 微生物學報, 49(3):343-350.
常華,2009. 廣東草魚養(yǎng)殖概況及現(xiàn)狀分析[J]. 海洋與漁業(yè):水產(chǎn)前沿, (6):50-52.
陳光榮,鐘萍,張修峰,等,2008. 惠州西湖浮游動物及其與水質(zhì)的關(guān)系[J]. 湖泊科學, 20(3):351-356.
陳佳林,余海軍,王茜,2021. 大清河流域浮游動物的群落特征研究[J]. 水生態(tài)學雜志, 42(3):72-78.
陳雷,徐兆禮,姚煒民,等,2009. 甌江口春季營養(yǎng)鹽、浮游植物和浮游動物的分布[J]. 生態(tài)學報, (3):1571-1577.
陳小江,汪祈超,2016. 湖北通海口地區(qū)草魚養(yǎng)殖模式存在的問題及對策[J]. 漁業(yè)致富指南, 4(20):34-35.
董雙林,2014. 中國綜合水產(chǎn)養(yǎng)殖的生態(tài)學基礎[M]. 北京:科學出版社.
高攀,蔣明,趙宇江,等,2009. 主養(yǎng)草魚池塘水質(zhì)指標的變化規(guī)律和氮磷收支[J]. 云南農(nóng)業(yè)大學學報, 24(1):71-77.
谷孝鴻,劉桂英,1996. 濾食性鰱鳙魚對池塘浮游生物的影響[J]. 農(nóng)村生態(tài)環(huán)境, 12(1):6-10,41.
國家環(huán)境保護局標準處,1989. 水質(zhì) 總磷的測定: 鉬酸銨分光光度法: GB/T 11893-1989[S].
國家環(huán)境保護局規(guī)劃標準處,1987. 水質(zhì) 亞硝酸鹽氮的測定: 分光光度法: GB 7493-87[S].
國家環(huán)境保護局規(guī)劃標準處,1989. 水質(zhì) 總氮的測定: 堿性過硫酸鉀消解紫外分光光度法: GB 11894-89[S].
國家環(huán)境保護總局,2002. 水和廢水監(jiān)測分析方法[M].4版. 北京:中國環(huán)境科學出版社.
國家環(huán)境保護總局科技標準司,2007. 水質(zhì) 硝酸鹽氮的測定: 紫外分光光度法(試行): HJ/T346-2007[S]. 北京: 中國環(huán)境科學出版社.
國家環(huán)境保護總局科技標準司,2009.水質(zhì) 氨氮的測定: 納氏試劑分光光度法: HJ 535-2009[S]. 北京: 中國環(huán)境科學出版社.
國家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系,2021. 草魚產(chǎn)業(yè)發(fā)展報告[J]. 中國水產(chǎn), (2):26-37.
李萌,尉婷婷,史博洋,等,2019. 環(huán)境DNA技術(shù)在淡水底棲大型無脊椎動物多樣性監(jiān)測中的應用[J].生物多樣性, 27(5):480-490.
李秋芬,陳碧鵑,曲克明,等,2002. 魚蝦混養(yǎng)生態(tài)系中細菌動態(tài)變化的研究[J]. 應用生態(tài)學報, 13(6):731-734.
李瑞嬌,2014. 草魚池塘浮游植物群落結(jié)構(gòu)及初級生產(chǎn)力、能量轉(zhuǎn)換的初步研究[D]. 武漢:華中農(nóng)業(yè)大學.
劉玲儀,金有坤,1994. 養(yǎng)殖魚類細菌性敗血癥暴發(fā)期間魚池水化學環(huán)境的初步調(diào)查[J]. 水產(chǎn)學報, 18(2):153-156.
劉朋,2012. 草魚不同混養(yǎng)系統(tǒng)有機碳、氮和磷收支及變動的初步研究[D]. 青島:中國海洋大學.
梅順,任青松,2017. 南陵縣草魚實用養(yǎng)殖模式調(diào)查研究[J]. 現(xiàn)代農(nóng)業(yè)科技,(15):236,240.
舒璐,林佳艷,徐源,等,2020. 基于環(huán)境DNA宏條形碼的洱海魚類多樣性研究[J]. 水生生物學報, 44(5):1080-1086.
孫靈毅,王力勇,徐惠章,2003. 單細胞藻類對營養(yǎng)鹽類的吸收[J]. 水產(chǎn)科技情報, (3):119-120.
孫云飛,2013. 草魚(Ctenopharyngodon idellus)混養(yǎng)系統(tǒng)氮磷收支和池塘水質(zhì)與底質(zhì)的比較研究[D]. 青島:中國海洋大學.
田相利,鄭瑤瑤,柳炳俊,等,2012. 草魚混養(yǎng)系統(tǒng)細菌數(shù)量變動和群落功能多樣性研究[J]. 中國海洋大學學報, 42(11):19-27.
王玉彬,2007. 茅蓮湖水產(chǎn)養(yǎng)殖池塘中浮游生物的研究[D]. 南昌:南昌大學.
徐蓮,孫紀全,吳曉磊,等,2012. 菌株Rhodococcus sp. Chr-9和Exiguobacterium sp. Chr-43的除鉻(VI)特性[J]. 應用與環(huán)境生物學報, 18 (6):971-977.
許鄭超,鄧燕飛,周軍,等,2021. 中華絨螯蟹養(yǎng)殖池塘菌群結(jié)構(gòu)和功能特征[J]. 生態(tài)學雜志, 40(7):2223-2233.
楊帆,2016. 湖北草魚養(yǎng)殖模式總結(jié)之新洲篇[J]. 當代水產(chǎn), 41(2):94-95.
葉曉彤,梁迪文,王慶,等,2020. 洞庭湖流域常德柳葉湖及其鄰近水體輪蟲群落結(jié)構(gòu)變化及其對環(huán)境因子的響應[J]. 湖泊科學, 32(4):1126-1139.
占家智,羊茜,2015. 高效養(yǎng)淡水魚[M]. 北京:機械工業(yè)出版社.
張雷鳴,原居林,倪蒙,等,2020. 兩種池塘養(yǎng)殖模式水質(zhì)因子和浮游植物群落比較分析[J]. 浙江農(nóng)業(yè)學報, 32(2):317-326.
張麗娟,徐杉,趙崢,等,2021. 環(huán)境DNA宏條形碼監(jiān)測湖泊真核浮游植物的精準性[J]. 環(huán)境科學, 42(2):796-807.
張瑩,石萍,馬炯,2013. 微小桿菌Exiguobacterium spp.及其環(huán)境應用研究進展[J]. 應用與環(huán)境生物學報, 19(5):898-904.
趙玉寶,1993. 鯉魚種和鰱鳙對池塘浮游生物的影響[J]. 生態(tài)學報,13(4):348-355.
周波,2015. 基于EwE模型的草魚綜合養(yǎng)殖池塘生態(tài)系統(tǒng)研究[D]. 青島:中國海洋大學.
Bakunina I I, Shevchenko L S, Nedashkovskaia O I, et al, 2000. Screening of marine bacteria for fucoidan hydrolases[J]. Mikrobiologiia, 69(3):370.
Harper L R, Buxton A S, Rees H C, et al, 2019. Prospects and challenges of environmental DNA (eDNA) monitoring in freshwater ponds[J]. Hydrobiologia, 826(1):25-41.
Ivanova E P, Sawabe T, Zhukova N V, et al, 2003. Occurrence and diversity of mesophilic Shewanella strains isolated from the North-West Pacific Ocean[J]. Systematic & Applied Microbiology, 26(2):293-301.
Li Z, Wang G, Yu E, et al, 2019. Artificial substrata increase pond farming density of grass carp (Ctenopharyngodon idella) by increasing the bacteria that participate in nitrogen and phosphorus cycles in pond water. PeerJ, 7:e7906.
Niu S, Zhang K, Li Z, et al, 2023. Analysis of the structure and function of microbial community in late-stage of grass carp (Ctenopharyngodon idella) farming ponds[J]. Aquaculture Reports, 30:101556.
Payne M S, Hall M R, Bannister R, et al, 2006. Microbial diversity within the water column of a larval rearing system for the ornate rock lobster (Panulirus ornatus)[J]. Aquaculture, 258(1/4):80-90.
Singer D, Seppey C, Lentendu G, et al, 2021. Protist taxonomic and functional diversity in soil, freshwater and marine ecosystems[J]. Environment International, 146:106262.
Taberlet P, Coissac E, Hajibabaei M, et al, 2012. Environmental DNA[J]. Molecular Ecology, 21(8):1789-1793.
Turker H, Eversole A G, Brune D E, 2003. Comparative Nile tilapia and silver carp filtration rates of Partitioned Aquaculture System phytoplankton[J]. Aquaculture, 220(1/4):449-457.
(責任編輯? ?張俊友? ?熊美華)
Water Quality and Bio-composition in a Monoculture
and Polyculture Grass Carp Pond
XIAO Shu‐wen1,2, LIU Xing‐guo2, LU Shi‐min2, ZHAO Yu‐xi1,2, GU Zhao‐jun2, ZHOU Run‐feng1,2
(1. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai? ?201306, P.R. China;
2. Fishery Machinery and Instrument Research Institute, Chinese Academy of Fishery Sciences,
Shanghai? ?200092, P.R. China)
Abstract:In this study, we analyzed the differences in water quality, plankton, benthos and bacterial community structure between monoculture and polyculture grass carp ponds (80:20). Water quality analysis, environmental DNA and traditional identification methods were used, and the water quality and biological composition of the two culturing methods were compared. We aimed to provide evidence for evaluating grass carp culture modes and construction of highly efficient grass carp culture systems. This study also explored the existing problems of using environmental DNA to monitor culture pond ecosystems. Results show that water quality in the polyculture pond was better than in the monoculture pond. Concentrations of TN, NO3--N, NH4+-N and NO2--N in the polyculture pond were lower, respectively, by 10.15%, 3.78%, 5.07% and 80.18%, and the concentrations of TP and SRP were 27.14% and 56.26% lower than in the monoculture pond. Chlorophyta, Cyanophyta and Cryptophyta were dominated in both ponds. The density of phytoplankton in the monoculture pond was 3.00×107 ind/L, lower than in polyculture pond (1.04×108 ind/L). Rotifers and protozoa were the dominant zooplankton groups in both ponds, with lower densities of Cladophora and Copepoda. The zooplankton density in the monoculture pond was higher than in the polyculture pond. In terms of benthos, Gastropoda, Oligochaeta and Chironomidae were observed in the monoculture pond, while only Gastropoda and Chironomidae were found in the polyculture pond. In terms of the microbial composition, Firmicutes was the dominant group in the monoculture pond, Proteobacteri was the dominant group in the polyculture pond and Proteobacteria was the dominant group in the sediments of both ponds. In conclusion, grass carp polyculture can improve the water quality of aquaculture ponds, increase phytoplankton richness and change the structure of aquaculture pond flora. This study provides a basis for optimizing grass carp culture, improving water quality and constructing an efficient culture system.
Key words:grass carp pond; culture mode; water quality parameters; biological composition; environmental DNA
收稿日期:2022-01-10? ? ? 修回日期:2022-10-21
基金項目:國家現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系建設項目(CARS-46);國家重點研發(fā)計劃項目(2019YFD0900300)。
作者簡介:肖述文,1998年生,女,碩士研究生,研究方向為水域生態(tài)修復。E-mail:949960731@qq.com
通信作者:劉興國。E-mail:liuxingguo@fmiri.ac.cn
